
朱顶红花部相关性状的杂种优势与混合遗传分析
2022-10-17 08:19汤肖玮邵和平仰小东张宁宁
江苏农业科学 2022年18期
汤肖玮, 刘 晨, 邵和平, 仰小东, 张宁宁
(1.江苏丘陵地区南京农业科学研究所,江苏南京 210046; 2.江苏开放大学,江苏南京 210036)
朱顶红()为石蒜科朱顶红属多年生花卉,花型、花色张扬大方,近年出现在各大花事活动上。不断扩大的需求促使朱顶红行业快速发展,我国朱顶红行业发展的“卡脖子”问题是自主知识产权品种匮乏。我国朱顶红育种研究起步较晚,加之育种周期长,使我国朱顶红产业落后于荷兰等花卉大国。所以,促进朱顶红育种效率的提升、缩短育种周期是第一要务,将主基因+多基因混合遗传模型理论方法结合杂交育种手段培育朱顶红新种质是较高效的途径之一,该方法能够有效地分析出目标性状主基因和多基因的数量,以及估计基因效应等相关遗传参数。在作物有关研究中,大豆耐盐性、甜玉米植株及穗部性状、水稻产量及穗部性状等已广泛运用此方法。在观赏植物的遗传分析中,该方法也备受青睐。Ye等分别利用此方法研究了紫薇植株结构、牡丹表型性状、月季花色以及菊花杂交群体花部性状的遗传规律。闫芳等对朱顶红若干观赏性状进行了遗传特性分析,但尚未通过数量性状主基因+多基因混合遗传模型分析其遗传机制。
目前,科研人员对于朱顶红的探索热衷于组培、基因功能分析和遗传多样性等方面。国内关于朱顶红杂交育种以及观赏性状方面已开展了相关研究,张宁宁等记录了24个朱顶红品种花期等观赏相关性状的差异。耿楠等发现朱顶红不同品种之间花茎、株高和冠幅等观赏性状变异性较大,且性状间具有不同程度的相关性,其中花茎高与株高、冠幅具有较高相关性。杨柳燕等通过设计34对朱顶红杂交组合,发现“圣诞快乐”作为母本时具有较高杂交结实率。花部性状作为朱顶红最主要的观赏性状,进一步探索其遗传机制是必要的。
本研究将花瓣数性状差异较大的朱顶红品种“粉色惊奇”和“火焰孔雀”进行人工杂交构建群体,分析其杂交后代花部相关性状的遗传分离情况。运用主基因+多基因的混合遗传理论模型,探索朱顶红杂交组合F代群体5个花部相关性状遗传规律,估算主基因的遗传率,掌握朱顶红5个花部性状的最适遗传模型和基因效应,以期为进一步提高朱顶红观赏价值、新种质的选育及后期基因定位等分子育种相关研究提供理论指导。
1 材料与方法
1.1 试验材料
2017年3月下旬,以江苏丘陵地区南京农业科学研究所保存的朱顶红品种“粉色惊奇”(单瓣)为母本,“火焰孔雀”(重瓣)为父本进行人工杂交。盛花期采集“火焰孔雀”的花粉均匀涂在“粉色惊奇”的柱头上,3 d后第2次授粉。果荚变黄变干后于2017年5月上旬采收,置于阴处晾干至开裂,将种子剥离,共获267粒F杂交种子。于5月中旬穴盘点播,后定植于盆内,同亲本于玻璃温室中统一管理。
1.2 田间性状调查
在2020、2021年连续2年春季对F群体(共88株)的5个花部相关性状进行观测,测定指标为花瓣数、花梗长、最大花被片长、最大花被片宽和花径,并计算2年性状调查的平均值。花瓣数中单重瓣分类标准按照6片花瓣的为单瓣,大于6片花瓣的为重瓣。
1.3 杂种优势分析及显著性检验
杂种优势通过中亲优势()和中亲优势率()表示。=-;=(/)×100%。其中,为杂交F后代单个性状的均值,(mid-parents value)为双亲单个性状的均值。利用SPSS 25.0数据分析软件的单样本均值检验分析F群体中亲优势的显著性。
1.4 主基因+多基因混合遗传分析

2 结果与分析
2.1 朱顶红F1代花部性状表型特征及杂种优势表现
亲本“粉色惊奇”“火焰孔雀”和F群体中部分品种表型见图1。在F后代群体中,花色以白色、白粉色、红色为主,多呈双亲过渡型颜色,花瓣数遗传变异范围为6~23瓣。
由表1可知, 5个花部相关性状在F代变异系数分别为46.99%、29.39%、8.35%、19.08%、13.37%,分离较广泛。其中,花瓣数变异系数最大,说明其离散程度最大,花径的变异系数最小,其离散程度最小。

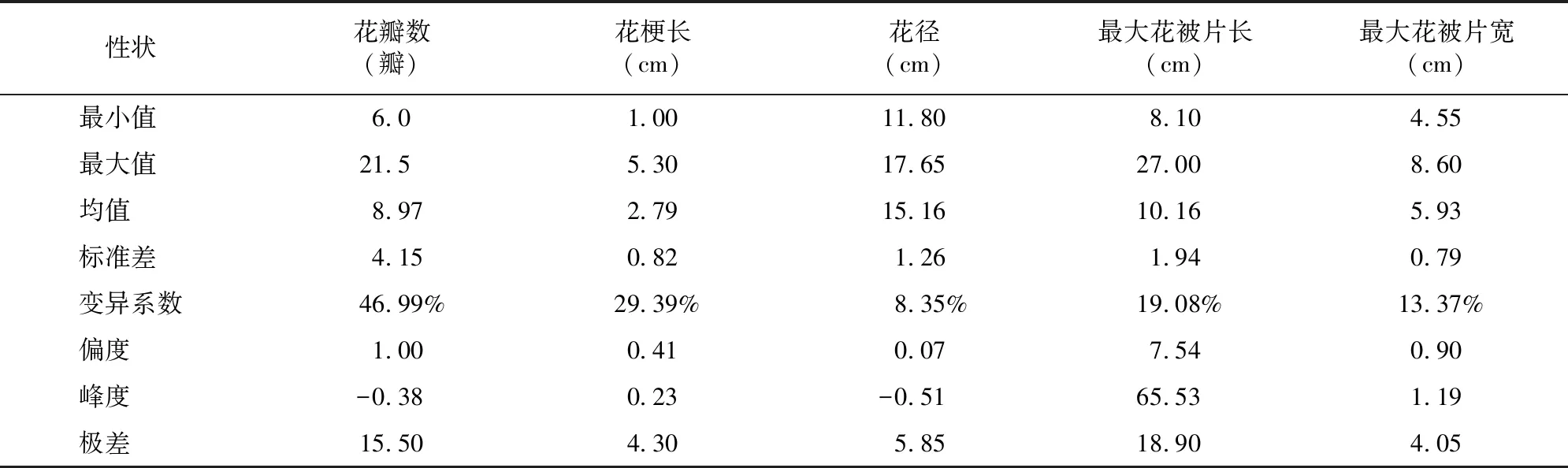
表1 朱顶红“粉色惊奇”ד火焰孔雀”杂交F1代花部相关性状表型特征值
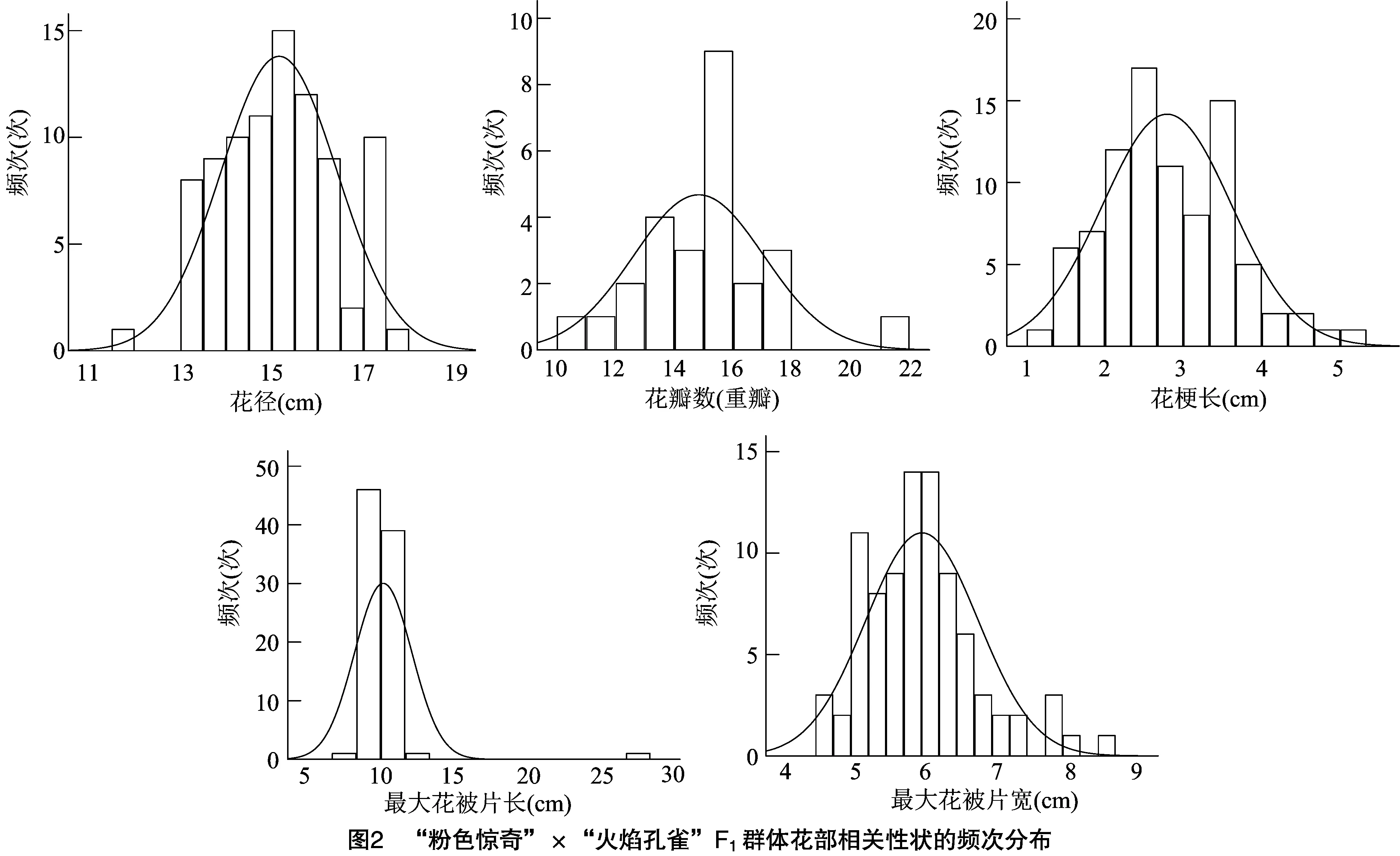
从F群体各花部相关性状的频次分布(图2)可知,花径、花梗长和最大花被片宽均呈连续性较好的正态分布。在统计的88份F群体中,按照花瓣数区分成单瓣和重瓣2类,其中单瓣共54个,占比为61.4%;重瓣26个,占比为29.5%,重瓣F代的花瓣数分析结果符合正态分布;另外有8株的2年数据瓣型相反,花瓣数不稳定。
由表2可以看出,中亲优势均达到极显著水平,F群体中5个性状中亲优势为负值,说明当前F代存在衰退现象。F代群体5个性状的值分别与亲本相比,均存在小于低亲或超过高亲的个体,说明F代普遍发生超亲分离,F群体中各性状的平均值均小于高值亲本,其中花瓣数和花梗长均值介于双亲之间,花径、最大花瓣片长和宽均值均低于低值亲本。

表2 朱顶红“粉色惊奇”ד火焰孔雀”杂交F1代花部相关性状的杂种优势表现
2.2 朱顶红花部相关性状的相关性分析
由表3可知,在10对花部性状间共有4对性状的相关性达到极显著水平,3对性状表现为显著相关。其中,花梗长与花径、 花梗长与花瓣数、最大花被片长与最大花被片宽均呈极显著正相关,相关系数分别为0.373、0.301、0.250;花瓣数与花径、花径与最大花被片长、花梗长与最大花被片宽呈显著正相关,相关系数分别为0.233、0.227、0.221;花瓣数与最大花被片宽呈极显著负相关,相关系数为-0.363。
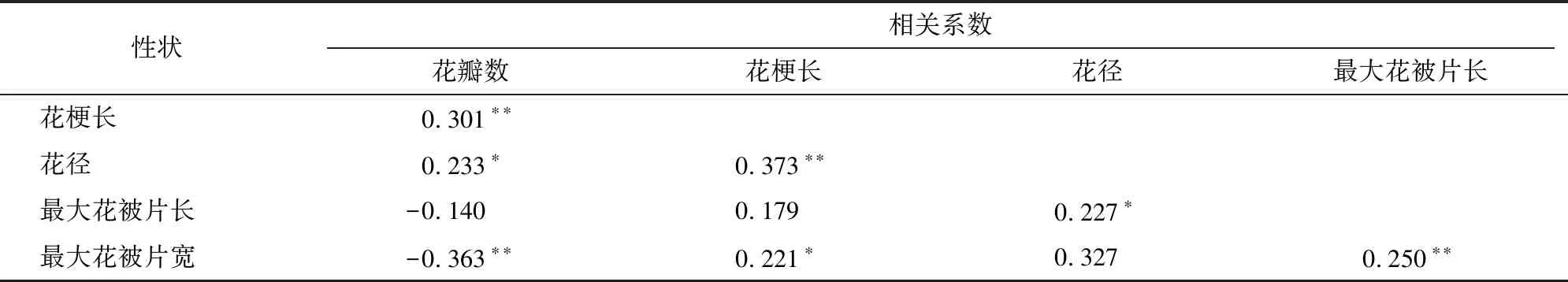
表3 F1代群体花部相关性状的相关性分析
2.3 最优遗传模型选择和适合性分析
利用朱顶红杂交后代5个花部性状2年数据均值,通过混合遗传模型分离分析法分析出极大似然值和AIC值,选取AIC最小的3个值对应的模型作为最适模型备选。如表4所示,A-0、A-2和B-2等3个模型花径对应的AIC值较小,分别为295.11、294.27和295.26。再通过适合性检验(表5),结合达到显著统计量个数最少原则选出最适模型。各性状筛选结果为:花梗长最优模型为A-0,无主基因控制;最大花被片长符合A-1模型, 受表现为加性和部分显性或超显性的1对主基因控制;花径最适模型为A-2模型,受表现为加性的1对主基因控制,说明显性效应值为0;最大花被片宽符合B-1模型,受加性-显性-上位性2对主基因控制;花瓣数的模型为B-2,受表现为加性-显性2对主基因控制。
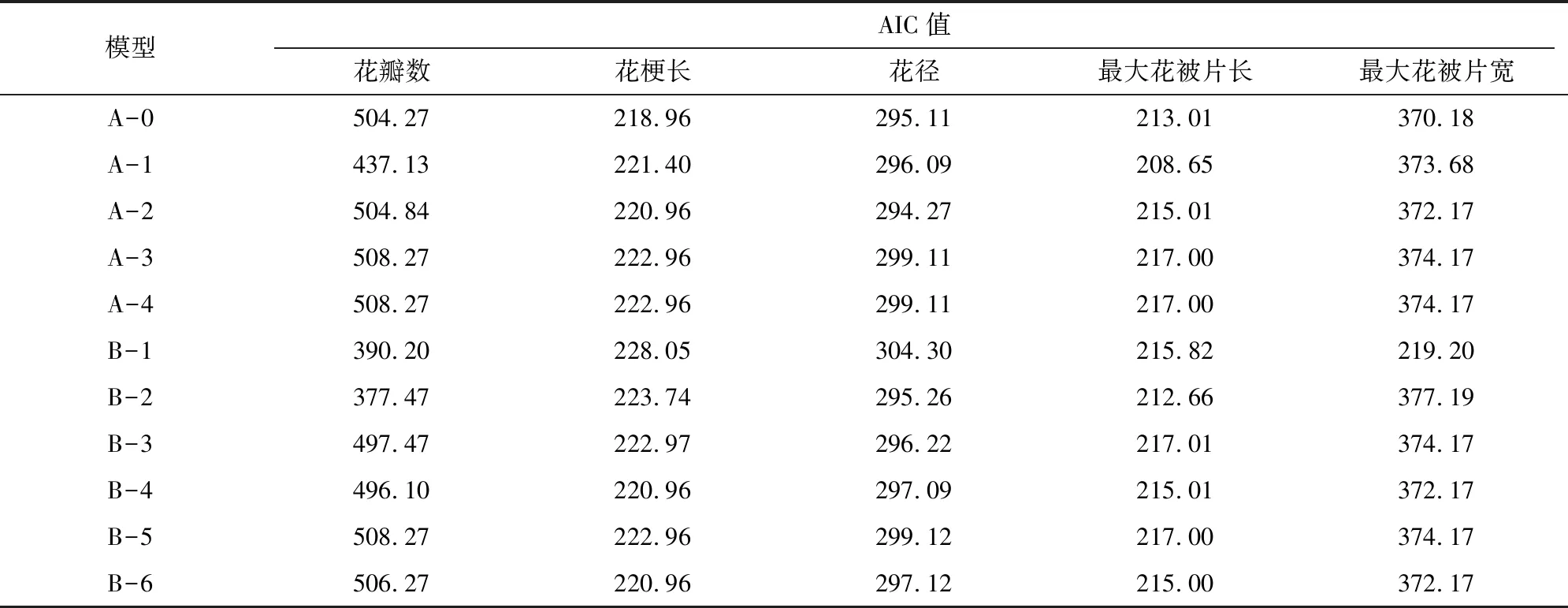
表4 朱顶红“粉色惊奇”ד火焰孔雀”杂交F1代花部相关性状分离分析的AIC值

表5 F1代群体花部相关性状入选模型的适合性测验结果
2.4 相关遗传参数
各性状最优模型的相关遗传参数见表6,花瓣数的2对主基因的加性效应值(、)均是正数,分别为4.35、3.56的正向增效,第1对主基因的加性效应大于第2对,所以该性状的加性效应以第1对为主,2对主基因的显性效应值是分别为-6.12和 -2.11,且显性效应应以第1对为主。受2对主基因影响的最大花被片宽的加性效应值均为4.26,且显性效应值分别为-6.39、-4.26,均为负向,两者之间的加性-加性效应、加性-显性效应、显性-加性效应、显性-显性效应值分别为4.26、-4.26、-2.13、6.39。花梗长和最大花被片长主基因的显性效应值均为负,分别为-0.54、-0.69,其加性效应均表现为正向效应,分别为0.56、0.69。花瓣数和最大花被片长的主基因遗传率分别为96%、47%。

表6 F1代群体花部相关性状在最优模型下的遗传参数估计值
3 讨论与结论
朱顶红易于在我国栽培,且近几年市场需求日益扩大。花部相关性状是衡量朱顶红观赏价值的重要指标,分析和利用其遗传特性就显得格外重要。本研究分析结果显示,5个花部相关性状在F群体的变异较大,变异系数为8.35%~46.99%,拓宽了其遗传基础。其中花瓣数变异系数最大,这是由于本研究选择的“粉色惊奇”“火焰孔雀”双亲在花瓣数性状上差异极大,导致该性状分离明显。此外,在本研究杂交群体中,比较5个花部性状的相关性发现,花径与最大花被片长呈显著性相关,与最大花被片宽相关性不显著,这与王秀刚等关于百合杂交F代群体遗传分析中,花冠直径和外轮花被片长具有极显著的相关性,与外轮花被片宽无显著相关性的结果一致。前人研究表明,不同性状之间的关系可以运用于育种,来预判某些相关性状的表现,以提高育种效率。本研究的5个性状均呈现负向的中亲性状,说明这些性状可能存在显性遗传效应,或F后代可能暂未表现出明显的超亲优势。闫芳等在朱顶红相关观赏性状遗传分析的研究中发现F代表现出较明显的衰退。
目前,已有较多观赏植物花部性状遗传机制相关的研究,主要包括兰花花部性状形态性状变异、分离和QTL定位的遗传分析、荷花花瓣数和花色等表型性状多样性的分析、菊花花径等园艺性状的关联分析、月季花径和花瓣数等性状的变异分析等。然而,目前关于朱顶红花部相关性状的遗传研究并未深入,极大程度限制了朱顶红花型育种进程。本研究使用主基因+多基因混合遗传分离分析法深入分析了朱顶红花部相关性状。结果发现,朱顶红的花梗长符合无主基因控制的A-0模型,其原因或是被环境影响比较大的多基因所控,其遗传规律可能较为复杂;花瓣数和最大花被片宽均受2对主基因控制,主基因表现分别为加性-显性和加性-显性-上位性;花径由1对主基因控制,这与张飞等菊花花器性状混合遗传分析中结果相同;最大花被片宽性状最适模型受2对主基因控制,二者之间存在上位性互作效应,而杂种优势的来源之一就是上位性互作效应,这为朱顶红花部相关性状的杂交育种奠定理论依据;花瓣数和最大花被片长2者主基因遗传效率分别为96%、47%,表明花瓣数属于高度遗传,即不易受环境影响,可为朱顶红育种的早期世代选择提供研究方向。
本研究初步分析了朱顶红花部性状的遗传规律及主基因间互作效应,有助于进一步阐明其遗传模式,为后续提高朱顶红花部相关性状育种效率奠定基础。
猜你喜欢
山西农业大学学报(自然科学版)(2023年2期)2023-06-23
当代人(2022年8期)2022-08-23
浙江林业(2021年8期)2021-09-22
延河·绿色文学(2020年6期)2020-09-10
东坡赤壁诗词(2019年5期)2019-11-14
中成药(2018年10期)2018-10-26
现代园艺(2018年3期)2018-02-10
现代园艺(2017年19期)2018-01-19
东坡赤壁诗词(2016年4期)2016-08-15
