
DNA去甲基化特征、功能及其在节肢动物中的研究现状
2022-10-15 09:02刘逸尘何雨欣张亦陈孙金生
天津师范大学学报(自然科学版) 2022年5期
刘逸尘,何雨欣,张亦陈,孙金生
(天津师范大学生命科学学院,天津 300387)
生物性状的表达不仅受基因控制,还受环境的影响.在环境影响下,生物基因未改变但表型发生可遗传变化的现象称为表观遗传,涉及DNA甲基化、组蛋白修饰、染色体重塑、非编码RNA调控等过程,其中研究最为广泛的是DNA甲基化[1].DNA甲基化是DNA的修饰方式之一,常见的修饰方式为DNA甲基转移酶(DNA methyltransferases,DNMTs)催化S-甲硫腺苷(SAM)的1个甲基转移到胞嘧啶(C)5号碳原子上,产生5-甲基化胞嘧啶(5mC),这种修饰作用可以影响转录因子、甲基化结合蛋白以及染色体结构,从而影响基因表达,参与转座因子沉默、基因组印记和X染色体失活等多种生物学过程.动物中最常见的为CpG二核苷酸位点中的甲基化,但基因组DNA甲基化的实际模式是高度可变的,尤其是在脊椎动物和无脊椎动物之间.无脊椎动物具有与脊椎动物类似的甲基化模式,但大部分的甲基化水平较脊椎动物低,还存在CHG(H代表A/T/C)和CHH序列上的甲基化修饰,例如果蝇(Drosophila melanogaster)中存在的CpA、CpT甲基化修饰以及菜蛾(Plutella xylostella)中存在的5′-CCGG-3′序列甲基化修饰[2].
DNA甲基化是动态变化的,它的异常可能会产生癌症、基因印记以及精神相关疾病的风险[3-4].作为DNA甲基化的逆向过程,DNA去甲基化在动物发育、脑可塑性、基因组印记等过程和由环境损伤引起的表观基因调控中具有高度灵活性[5],因此正常的DNA去甲基化修饰在维持DNA甲基化调控的稳定性方面具有重要作用.深入了解DNA去甲基化,能够完善对DNA甲基化动态调控的认识,这不但能拓宽人们对表观遗传调控的理解,还对体细胞重编程、再生医学以及医学治疗等具有重要意义.相比脊椎动物,DNA去甲基化在无脊椎动物中研究得较少,其中对节肢动物的研究大多聚焦在昆虫当中.本文对DNA去甲基化的机制、功能及其在节肢动物中的研究现状进行综述,以期为甲壳动物DNA去甲基化的研究提供借鉴和参考.
1 DNA去甲基化的方式及其作用机制
DNA去甲基化涉及被动和主动2种方式.被动去甲基化主要发生在细胞分裂时期,其实质是在DNMT被抑制的前提下通过DNA的复制对甲基化进行稀释,并不能完全去除DNA的甲基化标签.Leu等[6]研究发现,小的干扰RNA(smallinterferingRNA,siRNA)干扰卵巢癌细胞中的DNMT1和DNMT3b会引起部分基因位点的去甲基化,从而恢复基因表达,这也进一步说明了抑制DNMT的活性可以引起被动去甲基化.而主动去甲基化则不需要依靠DNA复制,早在1986年Razin等[7]就提出分化过程中的DNA去甲基化是通过一种酶反应机制实现的,即5mC被C取代.主动去甲基化的途径如图1[8-9]所示,共分为4种[10]:①碱基切除修复途径(base excision repair,BER),碱基切除修复过程中直接切除5mC并启动DNA糖基化酶来实现主动DNA去甲基化,这种机制多见于植物中;②脱氨基-错配修复途径,胞苷脱氨基酶(cytidine deaminase,CDA)可以催化5mC转变为胸腺嘧啶(T),产生的G∶T易突变体激活错配修复,错配修复将T修复为C从而达到去甲基化的目的[11];③TET-糖基化酶-切除修复途径,这是研究最为广泛也最公认的去甲基化途径[10-11],易位甲基胞嘧啶双加氧酶(ten-eleven translocation,TET)蛋白是一种依赖α-酮戊二酸(α-KG)和Fe2+的双加氧酶,TET蛋白能够将5mC氧化为5-羟甲基胞嘧啶(5hmC),并进一步将5 hmC氧化成甲酰胞嘧啶(5fC)、羧基胞嘧啶(5caC)[12],随后被T-DNA糖苷酶(TDG)或U-DNA糖苷酶(UDG)识别切除产生一个无碱基位点,进而经过碱基切除修复途径被修复为无修饰的胞嘧啶[13-14];④TET-脱氨基偶联途径,TET将5mC氧化为5hmC,CDA催化5hmC脱氨基产生的5-羟甲基尿嘧啶(5hmU)被TDG识别切除,达到去甲基化的目的[15-16].

图1 哺乳动物中DNA甲基化以及去甲基化途径Fig.1 DNA methylation and demethylation pathways in mammals
主动去甲基化与被动去甲基化过程并非完全独立和相互排斥,甚至存在关联.例如,哺乳动物生殖细胞发育过程中以及受精后均会伴随全基因组的去甲基化,其中,在合子基因组的去甲基化过程中,母系和父系基因组利用了不同的去甲基机制,父系基因组主动去甲基化,而母系基因组被动去甲基化[17].
2 DNA去甲基化的功能
哺乳动物中存在不同形式的DNA甲基化模式,包括启动子和非启动子的甲基化.启动子上的DNA甲基化通常会抑制基因表达,因此其变化在基因表达中具有开关的作用,对维持生物正常的细胞功能、遗传印记和胚胎发育极其重要.DNA去甲基化作为DNA甲基化修饰动态变化中的关键过程,与甲基化的作用紧密相关.
2.1 调控发育进程
DNA甲基化的动态调控对生物胚胎和脑的发育具有重要作用.在有性生殖中,基因组甲基化共发生2次动态变化,如图2[18]所示.一次发生在生殖细胞发育时期:原始生殖细胞需要经过被动的全基因组去甲基化以及主动的特异性去甲基化,消除体细胞中的特异甲基化修饰,随后在生殖细胞形成和配子发生过程中建立性别和生殖细胞特异性DNA甲基化模式,形成基因组印记,以便完成高度复杂化和专业化的减数分裂和受精过程;另一次则发生在早期胚胎发育时期:受精后的配子去除了除基因印记区和转座子以外的大部分甲基化标记,之后又在胚胎着床前重建DNA甲基化模式[18].Inoue等[19]研究发现,TET1缺失的小鼠胚胎干细胞不仅呈现出形态学异常,还表现出生长速率的下降.此外,相关研究表明,DNA去甲基化在神经元发育中起着关键作用,DNA去甲基化的异常通常伴随着雷特综合征、孤独症、阿尔茨海默病等疾病的发生.Szulwach等[20]研究脑发育时发现,5 hmC的获得与激活发育基因有关.5hmC、5fC和5caC是DNA去甲基化过程中关键的中间体,它们的含量在神经元发育以及神经干细胞分化过程中表现出时空差异性,TETs也具有类似的时空特异性表达[21],这也表明发育过程中DNA去甲基化可能有助于调节基因的正确表达.
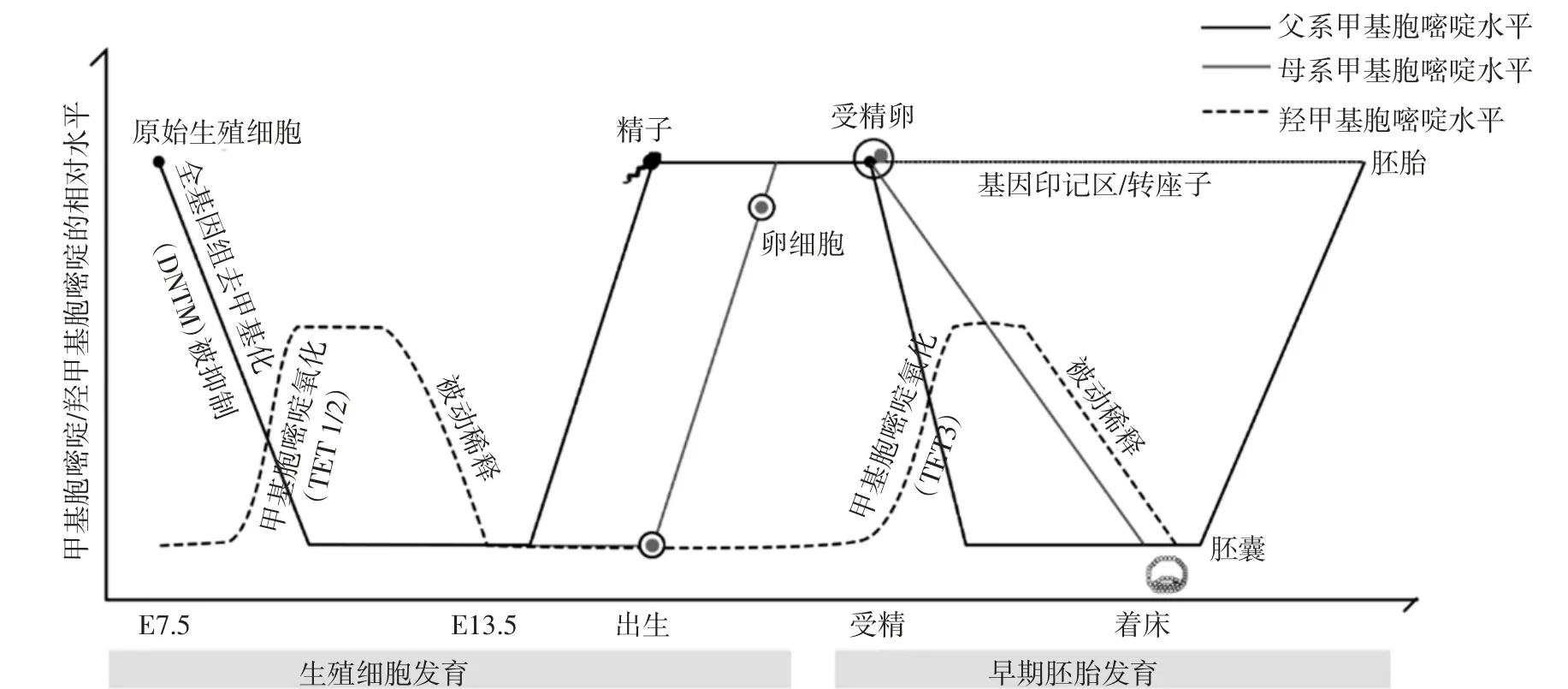
图2 DNA甲基化水平在配子发育以及早期胚胎发育时期的变化Fig.2 Changes of DNA methylation level during gametogenesis and embryogenesis
2.2 参与免疫防御过程
肿瘤形成与DNA去甲基化的异常有关.在髓系恶性肿瘤中首次发现的TET2突变基因使学者们逐渐关注TET2在免疫调节中的作用,后续研究发现,TET2丢失会使肥大细胞基因表达发生中断且基因组甲基化模式发生改变,从而影响肥大细胞的分化和细胞因子的产生[22].进一步研究发现,TET2还参与了调控机体的炎症反应:浆细胞样树突状细胞(plasmacytoid dendritic cells,pDCs)在病毒感染初期具有呈递抗原以及诱发炎症反应的重要作用,CXXC5(CXXC finger protein 5)招募TET2以促进干扰素调控因子IRF7的表达,从而参与调控干扰素IFN-α的表达,其诱导的IFN反应被证明与诱导T细胞反应密切相关[23].TET2还能改变mRNA的甲基化模式.巨噬细胞TET2被ADAR1招募于细胞因子信号抑制因子(suppressor of cytokine signaling 3,SOCS3)mRNA的3′端非翻译区(3′-UTR)上,引起SOCS3 mRNA的降解从而解除对JAK/STAT3通路的抑制[24].而炎症消退时,巨噬细胞中TET2结合组蛋白去乙酰化酶1/2(HDAC1/2)以促进组蛋白脱乙酰化,并抑制白介素IL-6和IL-1β的表达,也表明TET2参与的表观遗传调控是解决炎症的关键因素[25-26].
除此之外,组织特异性DNA去甲基化近年来也被认为是免疫T细胞和B细胞正常分化和作用所必需的,如图3所示[26-27].全基因组绘制5hmC图谱发现,5hmC富集与基因表达正相关,5hmC不仅在一些关键的T细胞谱系特异性转录因子(如Th-POK、T-bet、Foxp3等)基因体中沉积,还在免疫应答相关基因(如IL-4、IFNγ和IL-17)的转录区域中富集,促进表达以调控T细胞的分化和功能[27].TET2的突变会导致CD4-T淋巴细胞异常增殖以破坏T细胞的稳定性[28].TET2和TET3 mRNA在B细胞发育的各个阶段都大量表达.与正常小鼠相比,TET2/3缺失的小鼠骨髓中B细胞数量显著减少,并且从pro-B发育到pre-B的过程中出现部分阻滞,TET2/3的缺失还导致干扰素调控因子IRF4表达降低,损害免疫球蛋白Igκ的转录和重排[29-30].TET1蛋白可以通过调控组蛋白H4的乙酰化水平影响DNA修复,也表明了DNA去甲基化在免疫调控、肿瘤抑制以及DNA修复中有着重要作用[31-32].
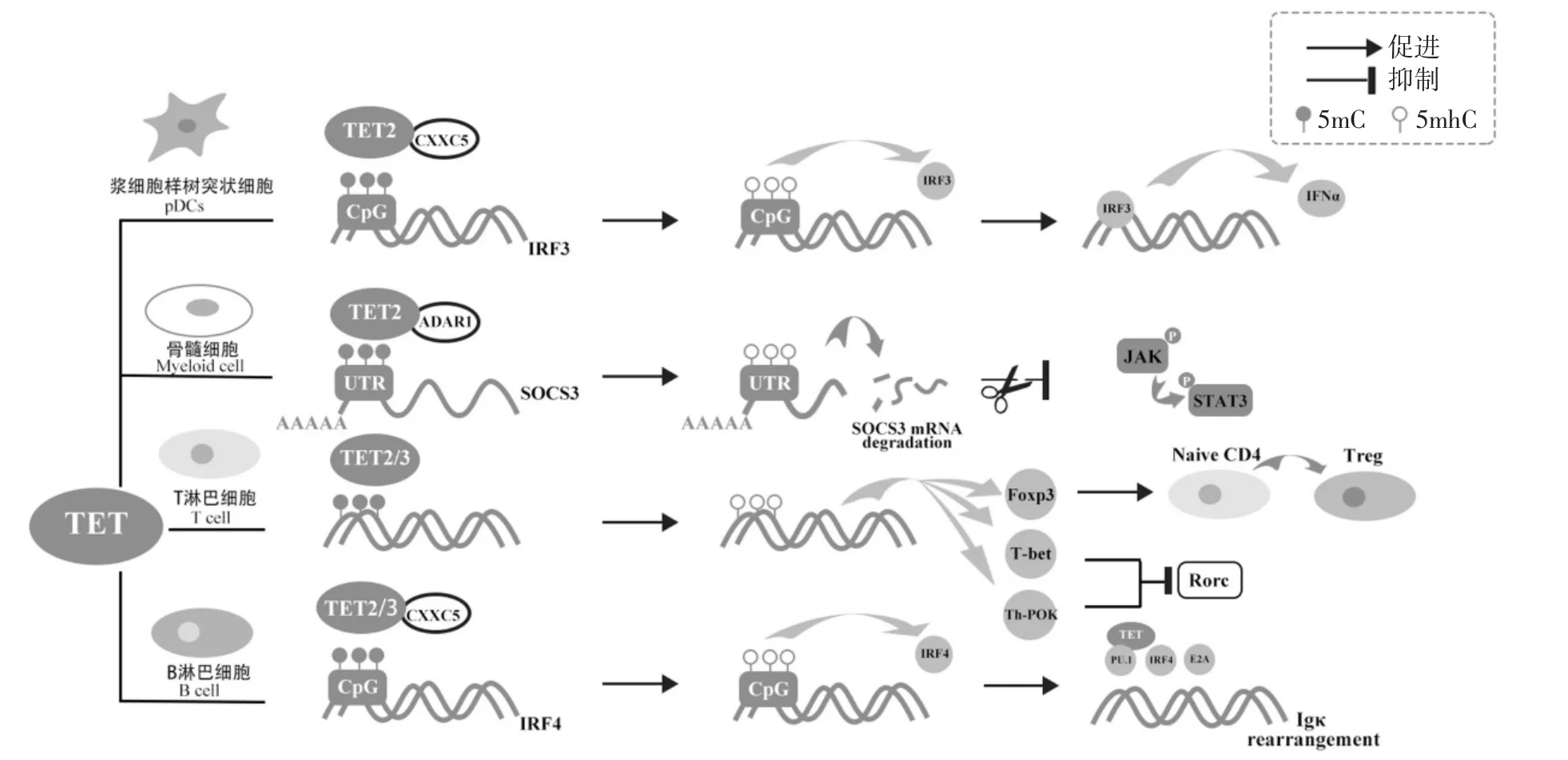
图3 TETs介导的DNA去甲基化在免疫相关应答中的调控Fig.3 Regulation of TETs-mediated DNA demethylation in immune-related responses
2.3 影响代谢通路
机体正常的新陈代谢与DNA甲基化的动态调节有着密切关联:一方面,DNA甲基化的动态修饰离不开机体代谢,其中用于修饰DNA的甲基来源于甲硫氨酸循环的中间产物S-腺苷甲硫氨酸,而参与去甲基化过程的TET双加氧酶活性也受到三羧酸循环中αKG的影响(图1);另一方面,在二型糖尿病患者中发现基因甲基化异常导致胰岛素和胰高血糖素分泌紊乱,这也表明了DNA甲基化对代谢的影响[9].高糖饮食会引发转化生长因子(transforming growth factor β1,TGFβ1)基因的去甲基化和表达,这在糖尿病和肾病的发病机制中起着关键作用[33].另外,TET2可调控脂蛋白脂肪酶(lipoprotein lipase,LPL)的表达,对甘油三酸酯的代谢有重要作用,TET2过度表达会显著抑制动脉粥样硬化病变的形成,这也给代谢紊乱引发的疾病提供了一个潜在的新治疗靶点[34].
2.4 潜在调控物种进化
基于DNA甲基化的动态调控作用,人们逐步开始探究DNA甲基化动态修饰在物种进化上的作用,DNA去甲基化引起的DNA甲基化水平改变可能会导致物种特异性适应以及表型变异.人类和黑猩猩的基因组具有95%的同源性,但逆转录转座子SVA的甲基化在人类和黑猩猩精子中显示出明显的物种特异性差异,SVA在人类精子中表现出低水平的甲基化而在黑猩猩精子中显示出高水平的甲基化,这种变化暗示了DNA去甲基化在进化中的调控作用[35].在脊椎动物中,DNA甲基化常发生在CpG二核苷酸序列上,CpG含量的变化也为甲基化水平的变化提供了一种机制,CpG含量降低可由甲基化胞嘧啶引起的快速脱氨基引起,使CpG转化为TpG[36].Gokhman等[37]在此基础上推断了尼安德特人和杰尼索万人的DNA甲基化模式,并观察到现代人中HOXD9启动子和HOXD10基因体的甲基化水平低于古人类中的甲基化水平,HOXD是调控肢体发育的关键蛋白,它在进化中发生的去甲基化可能解释了古人类与现代人类之间的一些解剖学差异[37-38].
3 节肢动物DNA去甲基化的研究现状
3.1 节肢动物去甲基化的机制
对节肢动物的全基因组进行甲基化检测,发现DNA甲基化存在于肢口纲(美洲鲎)、唇足纲(蜈蚣)、蛛形纲(腹蛛)、甲壳纲(大型溞)以及昆虫纲中,DNA甲基化的水平因物种而异,节肢动物DNA甲基化的机制以及分布如图4和表1[39-40]所示.在昆虫纲中,除双翅目外,DNA甲基化广泛存在于几个当前研究比较广泛的目中,如蜚蠊目、半翅目、膜翅目、鞘翅目、鳞翅目等.结合DNA甲基化转移酶检测发现,这些昆虫的DNA甲基化水平与DNMT1有相关性,与DNMT3之间则缺乏相关性[41].DNMT1和DNMT3在哺乳动物中分别具有催化半甲基化为全甲基化和对DNA进行从头甲基化的功能,但目前没有直接证据证明节肢动物中DNA从头甲基化由DNMT3介导.缺乏DNA甲基化的双翅目昆虫中只发现了DNMT2,这也与DNMT2催化tRNA甲基化的发现相符[40-42].基于DNMT重要的甲基化作用,其活性受到抑制以及表达受到干扰会使节肢动物产生被动的去甲基化.Zhang等[43]研究发现,利用RNA干扰技术敲降黄蜂(Nasonia vitripennis)的DNMT1基因能够显著降低全基因组甲基化水平,表明具有甲基化修饰的节肢动物存在DNA被动去甲基化修饰.

图4 节肢动物中DNMT和TET介导的DNA甲基化和去甲基化途径Fig.4 DNA methylation and demethylation mediated by DNMT and TET in arthropods
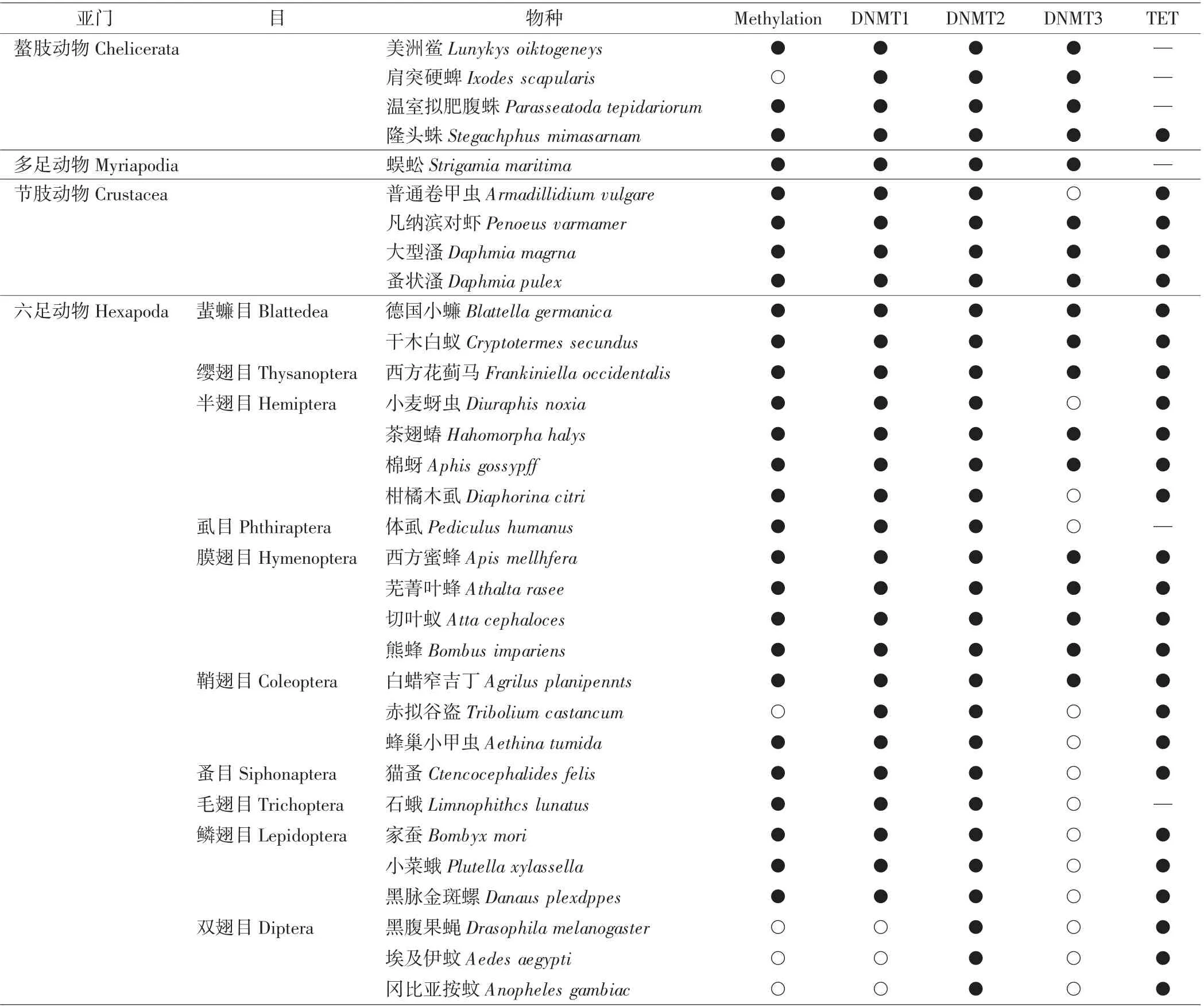
表1 节肢动物中DNMT和TET的分布Tab.1 Distribution of DNMA and TET in arthropods
节肢动物表现出与哺乳动物不同的甲基化模式.节肢动物DNA的甲基化往往以镶嵌模式稀疏地分布,甲基化多发生于基因的编码区且高度富集于编码保守蛋白的基因中,而在基因之间以及转录因子中的甲基化较少,启动子更缺少甲基化[39];相比于哺乳动物,部分节肢动物在CpG岛的甲基化水平较低,但存在CpT和CpA位点上的甲基化.常发生在细菌基因组中的腺嘌呤甲基化修饰(6mA)也存在于双翅目的果蝇中,并且在果蝇胚胎发育过程中具有潜在的调控作用[43].
哺乳动物中TET1/2/3蛋白在介导DNA主动去甲基化过程中有重要作用,在含有5hmC的节肢动物中也检测到了参与DNA主动去甲基化的关键酶类TETs,包括蚜虫(Diuraphis noxia)[44]、黄蜂[45]和蜜蜂(Apis mellifera)[46]等.不同于哺乳动物中常见的TET1/2/3蛋白,部分节肢动物中含有一个TET同系蛋白,即DNA甲基腺嘌呤脱甲基酶(DNA N6-Methyladenine demethylase,DMAD).Engelhardt等[40]研究发现,敲降果蝇的DMAD可导致6mA积累,经体外实验证明DMAD可介导6mA去甲基化.载脂蛋白B催化编辑蛋白家族(apolipoprotein B editing complex proteins,APOBECs)是常见于哺乳动物中的另一种去甲基化酶,虽然发现在无脊椎动物海胆中也存在该种酶[47],但对于节肢动物中APOBECs催化5mC脱氨基的研究尚未开展.虽然已经证明节肢动物中存在TET介导的DNA去甲基化修饰,但TET蛋白修饰后产生的5hmC是否经切除修复还原成未甲基化的胞嘧啶,仍需要进一步探讨.
3.2 节肢动物中DNA去甲基化功能的研究现状
哺乳动物中存在的启动子甲基化模式一般具有抑制效应.在昆虫中高度甲基化的基因通常具有管家基因功能,可以进行高效表达;而低水平甲基化的基因则可能与复杂的基因表达调控有关[48],其中可能涉及调控组蛋白修饰和选择性剪接.多项研究发现,DNA去甲基化对于节肢动物的发育、免疫防御、代谢以及进化具有重要影响,具体如表2所示.

表2 DNA去甲基化在节肢动物的发育、免疫防御、代谢以及进化中的调控作用Tab.2 Regulation of DNA demethylation in the development,immune defense,metabolism and evolution of arthropods
在发育方面,Kucharski等[49]首次在蜜蜂中得到启发,即昆虫的等级分化与表观遗传修饰密切相关,DNMT3基因的沉默会使西方蜜蜂幼虫发育成具有成熟卵巢的蜂皇.保幼激素(juvenile hormone,JH)是昆虫等级发育的关键因子,Cardoso-Júnior等[50]研究发现,无刺蜜蜂中低DNA甲基化水平能够上调保幼激素的表达,DNA甲基化和组蛋白磷酸化协同修饰,增加幼虫发育成蜂皇的可能.除此之外,Feliciello等[57]在赤拟谷盗(Tribolium castaneum)中也发现DNA甲基化参与调控胚胎发育成虫.Pegoraro等[45]研究发现,RNAi敲降黄蜂的Dnmt1a基因后在长日照条件下滞育发生率异常增加,表明甲基化对黄蜂正常光周期反应具有调控作用.Sp-CFSH(甲壳类雌性激素基因)是拟穴青蟹(Scylla paramamosain)性别分化的关键基因,该基因5′端的转录起始位点含有1个CpG岛,Jiang等[51]研究发现,定点突变与转录因子Sp1结合相关的CpG位点降低了启动子活性,表明甲基化可能通过阻断转录因子结合从而抑制Sp-CFSH表达.
在免疫防御方面,无脊椎动物不同于脊椎动物,仅具有天然免疫而不具有适应性免疫.DNA甲基化调控可能通过改变抗病和免疫相关基因的表达进而影响生物的免疫能力.家蚕(Bombyx mori)感染BmNPV病毒后会发生转录谱的变化,与正常家蚕相比有多个差异甲基化基因,其中包含参与JAK/STAT通路以及抗凋亡过程的相关基因,Huang等[53]研究发现,DNMT活性降低可以引起这些相关基因下调,促进感染细胞的凋亡,推测DNA去甲基化在抑制BmNPV病毒增殖方面有重要作用.Li-Byarlay等[54]对感染了IAPV的蜜蜂进行甲基化分析和转录组分析发现,DNA甲基化的快速变化先于并可能触发更广泛的转录组变化.Kumar等[55]用甲基化抑制剂处理小菜蛾(Plutella xylostella)时也发现基因组的5mC水平降低与免疫抑制有关,DNMT的表达和基因组的甲基化水平在被寄生蜂寄生后第3天发生显著降低;此外,他们还发现小菜蛾的5mC水平在被寄生的第2天显著升高,推测有可能是由于PxTET下调减少了5mC去甲基化代谢从而暂时增加了宿主防御能力.
在代谢方面,节肢动物具有与脊椎动物类似的现象,即某一特定基因的去甲基化能够影响相关的代谢调控.Foret等[56]将蜂王和工蜂幼虫的全基因组甲基化特征与基因表达和激素调节联系起来发现,几个高度保守的代谢和信号通路中富含甲基化基因,其中一个为差异甲基化区域编码间变性淋巴瘤激酶(ALK),ALK是新陈代谢重要的调控因子,这表明DNA甲基化表观遗传机制影响代谢通量和信号转导.
在进化方面,DNA去甲基化对节肢动物表型调控以及社会性分工具有调控作用.SAM蚜虫是从亲本SA1发展而来的一类更具毒性的生物型蚜虫,SAM比其亲本SA1具有更高的DNA甲基化调控活性,这可能是SAM能克服植物抗性的基础[44].社会性膜翅目昆虫表现出一定的DNA甲基化相似性和特异性,具有较高CpG含量的同时也存在DNA甲基化的基因调控[58],如具有社会性分工的蜜蜂,它们不同于果蝇,前者具有较高的CpG含量,且甲基化基因与脊椎动物的更相似[59].
目前有关于节肢动物DNA甲基化调控的研究给予了研究者许多启发,提出了进一步的思考:节肢动物是否具有与哺乳动物类似的主动去甲基化模式?环境变化是通过怎样的信号通路激活和调控相应的DNA甲基化调控通路?节肢动物中DNA甲基化是如何调控基因表达的?这种表达调控如何影响生物表型的变化?DNA甲基化调控如何与其他的表观遗传网路协调合作以调控生物表型的变化?
4 结论和展望
作为重要的表观遗传修饰之一,DNA甲基化动态调控已在哺乳动物中得到广泛研究,DNA去甲基化是机体维持正常DNA甲基化水平所必须的.哺乳动物的被动去甲基化和主动去甲基化协调合作,使生物机体维持稳定的DNA甲基化水平从而维系正常的生理活动,亦或改变机体的甲基化水平以应答环境变化.DNA去甲基化会引起哺乳动物生理功能的变化,其原因在于相关基因的甲基化水平发生改变,从而影响了它的表达水平.因此,甲基化与基因表达之间的相关性成为探究节肢动物DNA去甲基化功能和机制的关键问题之一.首先DNA甲基化作为保守的表观修饰之一,已在节肢动物中得到印证;其次,节肢动物中观察到的甲基化模式以及生理功能变化时发生的差异甲基化,暗示了DNA去甲基化与基因表达间的潜在关系,但节肢动物DNA去甲基化与基因表达是否有直接关系还未可知.节肢动物的甲基化常发生在基因体中,这种修饰常被认为调控转录本的选择性剪接和组蛋白修饰,该调控方式最近在蜜蜂[60]和家蚕[61]中得到验证,蜜蜂和家蚕提供了DNA甲基化调控基因表达的直接证据,也为推动节肢动物中DNA去甲基化的功能和机制研究奠定了基础.
DNA去甲基化是DNA甲基化动态调控不可缺少的一部分,要了解节肢动物DNA去甲基化如何响应环境变化,首先要对节肢动物的DNA甲基化和去甲基化机制有更完整的认知.DNA甲基转移酶是合成和维持DNA甲基化的关键因子,对DNA甲基化的动态调节至关重要.在昆虫体内,DNMT1存在率高且与DNA甲基化相关,而DNMT3的缺失率最高且与DNA甲基化缺少相关性,因此节肢动物在DNA甲基化的合成上还有待探究:是通过非DNMT蛋白所触发?还是DNA甲基化不再重新编程而直接由DNMT1维持稳定?为了稳定DNA甲基化修饰,一方面,可以通过DNA复制稀释来降低甲基化水平;另一方面,近年来对5hmC和TETs的探究也初步证明节肢动物可以通过TETs将5mC氧化为5hmC从而降低DNA甲基化水平.然而,TETs是否在节肢动物中介导了与哺乳动物类似的主动去甲基化尚未明确,这还需要对节肢动物中TETs的氧化产物(5fC和5caC)进行检测,并且对TETs与DNA糖苷酶的协同作用进行验证.
另外,DNA甲基化调控对生物生理功能的作用还有待进一步探究.将研究方向从整个组织转向发生显著变化的细胞或基因中,或许能提供更多DNA甲基化调控的线索.通过改变DNA甲基化的水平并分析其导致的相应后果,可以获得DNA甲基化特定作用的直接证据.RNA干扰技术和抑制剂(如DNA甲基化抑制剂5-azacytidine、DNA去甲基化抑制剂DMOG以及zebularine等)常被用于探究DNA甲基化调控功能的实验中,但这两种方法在作用程度以及特异性上仍存在限制.CRISPR技术也常被用于特定基因的功能分析且具有更好的准确性,近年来开发的一种基于CRISPR-Cas9的特异性DNA甲基化工具[62]为深入了解特定基因中甲基化的功能提供了技术支持,如利用dCas9-Tet1或-Dnmt3a融合蛋白靶向特定基因启动子区的去甲基化或甲基化,以观察该基因表达的变化[63].
DNA去甲基化在节肢动物中的研究已经取得了一定的进展,但依然存在许多疑问,而这些疑问也为未来更好地研究DNA甲基化以及去甲基化的作用提供了方向和线索.
猜你喜欢
今日农业(2021年11期)2021-08-13
科学大众(中学)(2021年3期)2021-05-17
林业科学(2020年12期)2021-01-26
中西医结合肝病杂志(2020年2期)2020-10-27
课外生活(小学1-3年级)(2018年10期)2018-11-19
小猕猴学习画刊(2018年9期)2018-09-22
中成药(2018年7期)2018-08-04
癌变·畸变·突变(2015年3期)2015-02-27
现代检验医学杂志(2015年2期)2015-02-06
遗传(2014年3期)2014-02-28
