
濒危植物香果树根萌苗和实生苗的种群特征比较
2022-10-15 09:02:48薛苹苹郭连金毛小涛肖志鹏吴艳萍王宝云范红霞
天津师范大学学报(自然科学版) 2022年5期
薛苹苹,郭连金,王 帆,毛小涛,肖志鹏,吴艳萍,王宝云,曾 颖,范红霞
(1.上饶师范学院生命科学学院,江西 上饶 334001;2.山东省庆云县农业局,山东 德州 253700)
植物种群的自然更新是一个复杂的生态学过程,它对种群的增殖、扩散、延续以及稳定具有重要作用[1].植物种群自然更新的前提是有足够数量的幼苗存活[2],它是打破濒危植物濒临灭绝的关键.目前,对大多数濒危植物的研究主要集中于种群结构及空间分布格局[3-4]、种子雨与种子库[5]、种子萌发及幼苗生长[6]、光合作用与蒸腾作用[7]以及遗传多样性[8-9]等方面.原生生境中部分濒危植物如峨眉冷杉、蒙古扁桃、沙冬青等虽然拥有较高的种子产量和扩散范围,但由于种子被捕食、霉烂以及遭遇环境胁迫等,种子萌发数量或形成幼苗的数量较少[5,10],且大部分物种的实生苗生长缓慢和抗性较差[11],因此在原生生境中有些濒危植物的自然更新主要依赖于伐桩和根上萌蘖产生的萌苗来完成[12-13].有研究表明树木的伐桩及其损伤的根系可产生大量的萌苗[14-15],但伐桩萌苗和根萌苗对植物种群恢复的贡献鲜见报道[16-17],关于伐桩萌苗、根萌苗和实生苗对植物种群更新贡献的比较更为少见[18].科学认识一种植物的天然更新特性是制定合理的人工促进天然更新措施以及进行自然资源保护的生态学基础[15].
香果树(Emmenopterys henryi)为中国特有的茜草科(Rubiaceae Juss.)单种属落叶乔木,是研究茜草科系统发育、形态演化及中国植物地理区系的重要材料.近年来,由于环境及人为干扰等因素的影响,该物种已濒临灭绝,被列为国家Ⅱ级重点保护稀有植物和林业部公布的国家珍贵树种[19].香果树种子微小,寿命短,属于光敏感型种子,千粒重仅为0.50 g左右,原生生境中发芽率极低[20].原生生境中的香果树种群主要存在实生苗和根萌苗2种更新方式[13,21-22].香果树群落郁闭度较高,且幼苗主要存在于乔木树冠下,光照条件较差[13],对于此环境中香果树这2种起源幼苗的生长状况以及它们在香果树种群自然更新中的作用尚无人报道.本研究以武夷山国家级自然保护区内的香果树种群为研究对象,比较实生苗和根萌苗的年龄结构、数量特征、生长特征、自然更新能力以及对树荫的忍耐力,旨在阐明原生生境中香果树不同起源幼苗的生存状况及香果树种群的更新策略,为香果树种群恢复提供理论依据和实践指导.
1 研究地自然概况
研究地位于武夷山国家级自然保护区(27°33′~27°54′N,117°27′~117°51′E),是世界同纬度现存面积最大、保存最完整的中亚热带森林生态系统,区内自然资源丰富,保存有完好的地带性常绿阔叶林群落,是东南亚大陆生物多样性最丰富的地区,也是具有全球生物多样性保护意义的地区.本区气候属于典型的亚热带季风气候,平均气温为12~13℃,年降水量1 480~2 150 mm,年蒸发量1 492 mm左右,相对湿度78%~84%,无霜期253~272 d.土壤类型主要有红壤、黄红壤、黄壤和山地草甸土[23].香果树在研究地主要星散分布于沟谷水溪旁的阔叶林中,其伴生种主要有银鹊树(Tapiscia Sinensis)、杉木(Cunninghamia lanceolata)、红脉钓樟(Lindera rubronervia)、盐肤木(Rhus chinensis)、野鸭椿(Euscaphis japonica)等.
2 研究方法
2.1 样地调查
在对武夷山自然保护区进行全面调查的基础上,在香果树分布比较典型的区域设置4个样地,面积为20 m×20 m,自然概况如表1所示.

表1 香果树样地概况Tab.1 Genernal situation of the habitats of E.henryi
调查样地生境:①确定生境的地貌地形、土壤、坡向、坡位、光照、温度、土壤含水量等生态因子,使用ZDS-10型光照计测定光照强度,使用DHM2型通风干湿温度计测定大气温度和湿度,在距地面0.5 m处进行测定[13].②确定不同起源的幼苗,观察种子萌发和幼苗根系情况以确定幼苗来源,标记实生苗和根萌苗.
2.2 不同起源幼苗的种群结构及生长特征
2015年11月,将样地内所有幼苗按照起源分类,采用幼苗主茎芽鳞痕的数目、节间的长短规律以及茎干表皮与周皮的纹路确定其年龄,每增长1 a设置1个年龄级,即1 a、2 a、3 a,以此类推.统计林内1~8 a幼苗中每级个体株数,作为该种群动态分析的基础数据.根据每级个体数的统计绘制年龄结构图.根据种群各龄级株数编制生命表,根据生命表中的标准化存活数以及死亡数绘制存活曲线和死亡曲线[24].记录每株幼苗的高度、基径、叶数,并利用活体叶面积仪(LA-S,美国CID公司)测定其叶面积,摘取叶片自然晾干,测定其干重并计算比叶重,以了解不同起源香果树幼苗的生长动态.
2.3 不同微生境中幼苗的生长特征
随机标记林内香果树母树或其他乔木的树冠下、冠缘、林窗以及林缘空地的香果树实生苗和根萌苗各10株,记录并测定其1年生侧枝的基径、枝长、叶数、节数、叶面积以及叶厚.利用打孔器于每个新生枝条上随机选取20个健康成熟的叶片,在主脉两侧各打3个圆片,其中30个圆片测其鲜重,自然晾干后再测其干重,以计算叶片含水量,另外30个圆片放入冰箱后迅速带回实验室进行叶绿素含量的测定[25].
2.4 不同微生境中幼苗的光合作用特征
于2015年8月,选择晴天,利用便携式光合仪(CI-340,美国CID公司)对所标记香果树幼苗的叶片进行光合作用测定,从8∶00—16∶00每隔2 h测定1次,测试采用开放式气路,测定植物的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)以及蒸腾速率(Tr).为了使2种幼苗的光合作用具有可比性,均选取1年生枝条由上到下的第3对叶片,每株选择6片,每次测定时间为30 min.
2.5 数据处理
采用SPSS19.0对数据平均值进行单因素方差分析、多重比较以及相关性分析,采用Origin9.0制图.
3 结果与分析
3.1 香果树幼苗的组成
武夷山香果树种群幼苗相对较少,2015年11月份平均每个种群中仅有32.5株1 a幼苗存活,其中根萌苗约占38%,不同起源幼苗的具体年龄结构如图1所示.由图1可以看出,随着年龄的增加,香果树幼苗的数量持续下降,至5 a后种群数量逐渐稳定在2.3株左右,幼苗的密度约为56.2株/hm2,期间3 a、5 a和7 a实生苗缺失.
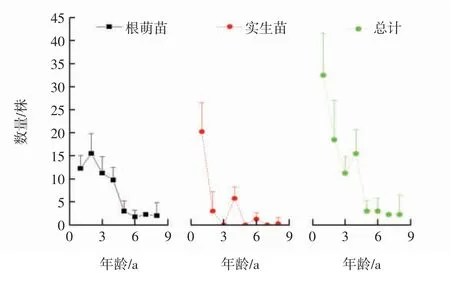
图1 香果树幼苗的年龄结构Fig.1 Age class structure of the saplings of E.henryi
3.2 香果树幼苗的生命表及存活曲线
根据香果树种群各龄级幼苗的数量编制生命表,结果如表2所示,绘制幼苗存活曲线结果如图2所示.由表2中的死亡率和消失率可知,香果树幼苗总体在3 a时出现负值,根萌苗在1 a和6 a时出现负值,表明此年龄段幼苗数量不足.香果树幼苗介于Deevey-Ⅱ型和Deevey-Ⅲ型之间,经数学模型检验可知,香果树幼苗总体和实生苗的存活曲线经幂函数拟合所得F值与R2均大于线性函数的数值,故两者更接近于Deevey-Ⅲ型;根萌苗存活曲线的拟合方程中,幂函数拟合方程的F值较大而线性函数的R2较大,从而表明根萌苗的存活曲线介于Deevey-Ⅱ型和Deevey-Ⅲ型之间.
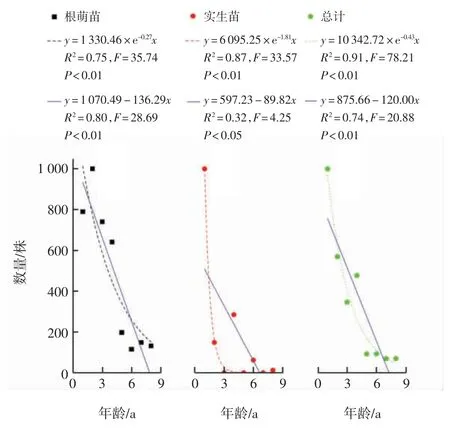
图2 香果树幼苗存活曲线Fig.2 Survival curve of the saplings of E.henryi
3.3 香果树幼苗的生长特征
武夷山香果树原生生境中根萌苗的生长较早,每年3月中旬开始展叶,而实生苗较晚,4月初香果树种子露白萌发,4月中旬出现真叶.观察测量实生苗和根萌苗的形态指标,结果如表3所示.由表3可以看出,香果树根萌苗的高度、基径、叶面积、比叶重均大于实生苗的数值,叶数小于实生苗的数值,两种幼苗在1 a、2 a时的形态指标差距较小,随着年龄增加差距逐渐增大.5个形态指标中,两种幼苗的苗高差距最大,根萌苗苗高为实生苗的2~3倍.

表3 香果树幼苗生长特征Tab.3 Growth characteristics of the saplings of E.henryi
3.4 香果树幼苗的耐阴性
对位于原生生境中不同位置的香果树幼苗的新生侧枝进行形态观测,结果如表4所示.由表4可以看出,冠下幼苗新生侧枝的基径和叶厚最小,叶面积最大;位于冠缘幼苗新生侧枝的枝长、节间长度、叶面积较大,叶厚相对较小;位于林窗的幼苗新生侧枝的基径、叶厚较大;同类型幼苗相比,位于林缘空地幼苗新生侧枝的基径、枝长和叶厚均最大,林缘空地下实生苗的叶片面积最小,而根萌苗的叶面积普遍高于实生苗.即相同微生境中实生苗的各项指标均显著小于根萌苗,其中根萌苗的叶面积、节间长度以及枝长为实生苗的4~5倍,基径和叶厚约为实生苗的1.4倍.
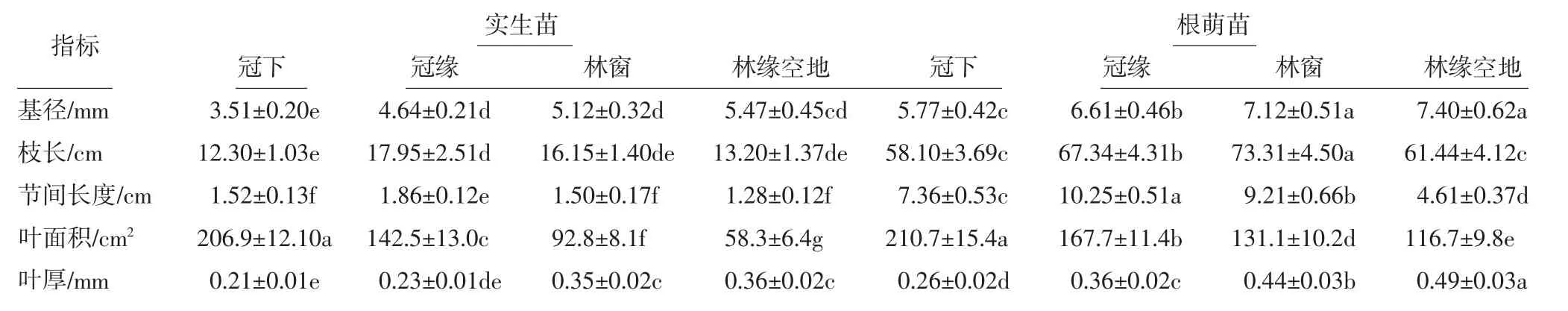
表4 香果树新生苗侧枝形态特征Tab.4 Morphological characteristics of annual branches of E.henryi
不同微生境中香果树幼苗新生侧枝上成熟叶片的各项生理指标差异较大,如表5所示.其中,冠下的幼苗叶片含水量和胞间CO2浓度显著大于其他微生境中的数值,林缘空地处的幼苗叶片含水量最低,林窗下幼苗叶片的胞间CO2浓度最低;叶绿素含量、Ca/Cb、净光合速率以及气孔导度则以冠缘或林窗生境中最大,其他微生境较小;蒸腾速率与叶片含水量相反,冠下幼苗最低,林缘空地幼苗最高.香果树根萌苗新生侧枝成熟叶片的含水量均显著小于同生境中实生苗的数值,除冠下幼苗外根萌苗的蒸腾速率均显著大于实生苗的数值.
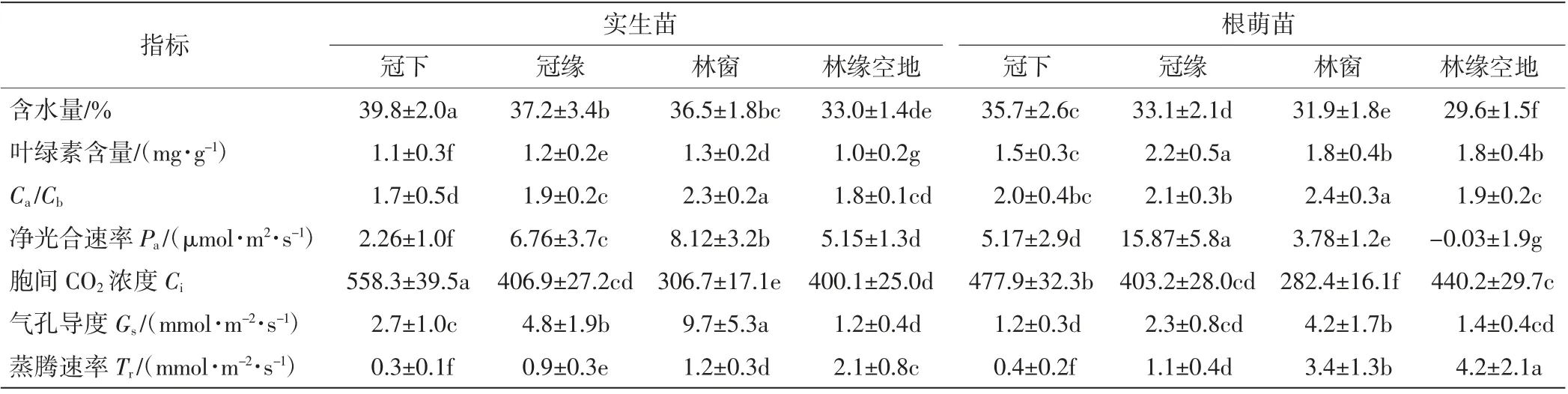
表5 香果树新生苗侧枝光合生理指标Tab.5 Photosynthetic physiological index of annual branches of E.henryi
4 讨论与结论
4.1 香果树根萌苗和实生苗的种群结构
分析种群的年龄结构是揭示种群动态变化和预测种群发展趋势的重要手段[26],武夷山香果树种群由于大多生长于溪流边,林内存在大量砾石,幼苗很难存活,其存活曲线整体呈Deevey-Ⅲ型,种群呈衰退型[23].本研究发现香果树幼苗较少,尽管1 a实生苗的数量较多,但大多数个体在生长过程中死亡,所调查的4个样地中仅存活了1株8 a实生苗,3 a、5 a和7 a实生苗缺失,这是由于香果树每2~4 a开花结果一次[27],其种子寿命极短,常温下仅能存活10个月[21,28],次年存留于土壤中的种子无法萌发,导致实生苗的年龄不连续.根萌苗的数量较多,占幼苗总体的65.44%,1 a苗个体每个样地约有12.25株,但8 a苗则减少到2株左右,由此可见香果树幼苗数量极低,无法维持种群稳定,种群呈衰退型.拟合曲线显示1~8 a香果树幼苗的存活曲线更接近于Deevey-Ⅲ型,这表明其低龄幼苗死亡率极高.
4.2 香果树根萌苗和实生苗的生长特性比较
有研究表明,在植物自然更新过程中,根萌苗比实生苗更具优势[29-30],但也有学者认为某些植物(如喜光先锋物种Cordia alliodora、Casearia gossypiosperma)的实生苗和根萌苗一样具有较高的生长速率[31].本研究结果发现,香果树根萌苗萌芽时间早,生长迅速,在基径、苗高、叶数、叶面积、比叶重等方面均高于实生苗的数值,二者各年龄(1 a、2 a、4 a、6 a、8 a)的平均比值依次为2.45、2.37、1.10、1.85、1.32,且两种幼苗的基径、苗高和叶数随着年龄增加差距逐渐增大,这表明香果树的根萌苗通过获取母树的营养,在短时间内地上和地下部分功能发育完善,随着年龄的增加,其竞争优势更加突出.
4.3 香果树幼苗自然更新能力和对树荫的忍耐力
自然界中通过种子进行更新的物种较少[31],这可能是由于种子被捕食率高、种子活力较低、实生苗死亡率高等原因导致.香果树开花量大,结实较少,虽然果实中含种子数量较多,但由于种子微小、寿命极短、幼苗生长缓慢,导致种子活力、种子萌发过程及幼苗极易受到环境影响;而根萌苗生长迅速,抵抗力强,不易受环境影响[32].这使得利用根、茎的萌蘖力增加幼苗数量成为香果树自然更新的重要途径.本研究发现香果树幼苗主要存在于4种微生境中,即树冠下、冠缘下、林窗以及林缘空地中.通过对香果树根萌苗及实生苗的新生侧枝进行观测,发现香果树根萌苗在这4种微生境中的生长均优于实生苗,在相同微生境中两种幼苗同一形态指标的数值差异均具有统计学意义,且冠下根萌苗侧枝基径和苗高的生长速率高于其他生境,这表明香果树根萌苗比实生苗更适合生存于阴暗环境中.光合生理指标则显示根萌苗叶片含水量低、蒸腾作用强、胞间CO2浓度低,且冠下的叶片叶绿素含量高,光合有效辐射高,这些均表明根萌苗在阴暗环境中利用光能的能力比实生苗强,在香果树种群更新中优于实生苗.
4.4 香果树自然更新建议
香果树原生生境中根萌苗数量较多,在其自然更新中具有不可替代的作用.本研究发现根萌苗在冠缘和林窗下的生长显著优于冠下和林缘空地,实生苗也有类似规律,这表明冠缘和林窗是香果树幼苗生存较适宜的环境,这可能是因为光照及该微生境的水热组合较有利于幼苗生存.林缘空地的幼苗节间较短,生长缓慢,长势较差,根萌苗的光合有效辐射甚至为负值,表明光照太强会抑制幼苗的形态建成,不利于其光合作用,故应对此生境幼苗进行遮荫处理.而冠下幼苗所处位置光照较弱,应清理地被物,增加透光率,以利于其进行光合作用,促进快速生长.
猜你喜欢
浙江农业科学(2024年2期)2024-02-27 15:03:24
农业科技通讯(2022年1期)2022-01-17 06:06:40
四川林业科技(2019年5期)2019-07-07 07:14:30
山东林业科技(2019年2期)2019-06-03 10:10:54
安徽农学通报(2019年1期)2019-02-14 02:25:08
湖北林业科技(2019年6期)2019-01-06 02:14:29
——以新源县为例
新疆农业科技(2018年5期)2018-05-07 02:25:08
农村农业农民·B版(2018年2期)2018-03-22 08:47:22
农村.农业.农民(2018年4期)2018-01-23 02:07:28
湖北林业科技(2017年2期)2017-05-25 18:44:32
