
BRs生物活性研究进展
2022-10-14 11:36刘金娜
现代农业科技 2022年19期
刘金娜
(1杨凌职业技术学院,陕西杨凌 712100;2西北农林科技大学生命科学学院,陕西杨凌 712100)
油菜素甾醇类化合物(brassinosteroids,简称BRs)是一类从植物中分离获得且具有多羟基结构的甾体化合物,其中活性最好的芸苔素内酯(brassinolide,BL)于1970年被发现,1978年Mandava等将其纯化并命名为brassinolide,1979年Grove等确定了其结构[1-3](图1)。 1982 年,Yokota 等[4]从栗瘿蜂的组织中获得了BL的合成前体,其B环含有酮型结构,被称为栗甾酮(castasterone,CS)。随后在一些低等、高等植物的花粉以及幼嫩器官中发现了BL、CS类似物,且该类化合物均表现出了相似的促进植物生长的活性,研究者将这些物质命名为BRs。在第16届国际植物生长物质学会会议(IPGSA)上,BRs被认定为继生长素、细胞分裂素、赤霉素、乙烯、脱落酸后的第六大植物激素[5]。至今已有80余种BRs被发现,它们可促进植物生长,提高植物抗性,缓解重金属毒害,调节各大激素平衡,同时还具有一定的保鲜作用和药理活性,是一种环保型的植物生长调节剂。本文总结了BRs的生理效应、保鲜作用和药理活性,展望了其在农业中的应用前景,以期为新型植物生长调节剂的研发提供一定参考。
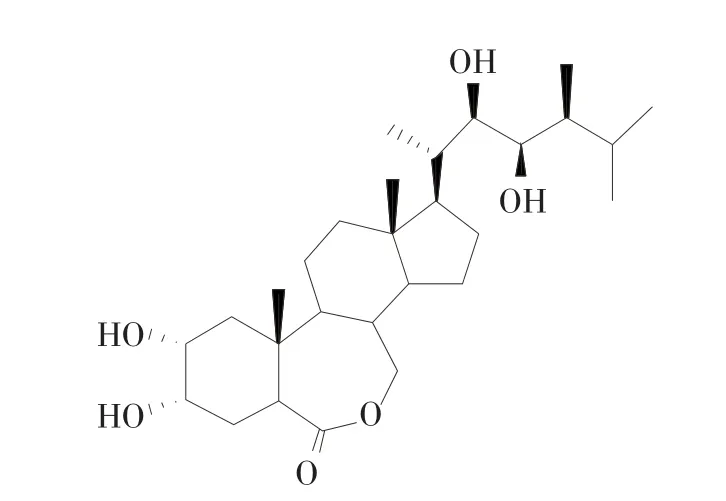
图1 油菜素内酯结构式
1 生理效应
1.1 促进植物生长
BRs最重要的生理功能就是促进植物生长。BRs可促进细胞分裂,引起植物茎节的膨大、伸长、开裂以及弯曲,最早采用大豆第二节间法验证了其类似的活性[6]。此外,BRs还能促进花粉管的伸长,促进受精,进而提高作物的坐果率与结实率,增加千粒重[7]。前人研究发现,外源施用纳摩尔级的BL就能使菊芋导管物质的分化速度提高10倍[8]。植物的衰老受植物激素调控,细胞分裂素可以延缓植物叶片的衰老,而24-表油菜素内酯能加速衰老,特别是在高浓度条件下,可通过提高植物体内过氧化物酶的活性、降低蛋白酶的活性来实现[9]。BRs还可促进种子的萌发和幼苗的生长。BRs浸种可提高陆地棉和黑吉豆的发芽率(分别提高了39%和23.3%),促进2种植物幼苗根的伸长、干重和鲜重的增加以及叶面积的增大[10]。BRs也能促进果实的成熟。Symons等[11]研究表明,6-去氧栗甾酮和栗甾酮可调控葡萄的成熟,且具有一定的促进作用。
油菜素内酯能提高植物光合速率,调控类囊体膜的形态建成以及光系统Ⅱ的作用[12]。外施0.1 mg/L 24-表油菜素内酯可以增加黄瓜中蔗糖、可溶性糖以及淀粉的含量,还能增强卡尔文循环下二磷酸核酮糖羧化酶的活性,进而增加CO2的同化量,提高产量[13]。胡文海等[14]研究表明:24-表油菜素内酯可以防止黄瓜低温光抑制的发生,处理1 d后,黄瓜净光合速率明显提高;此外,24-表油菜素内酯还能通过加强光系统Ⅱ反应中心的开放程度提高光电子传递量子的能力,但不影响天线色素的光能转换效率。
BRs可以调控植物的农艺学特征。28-高油菜素内酯浸种可提高扁豆硝酸还原酶的活力,增加其固氮能力,1×10-8mol/L下浸种8 h可使种子千粒重增加1.2 g[7]。喷施0.1 mg/L BRs可以显著增加西瓜产量,改善西瓜品质,尤其是可以提高西瓜可溶性总糖和番茄红素的含量[15]。从天竺葵中提炼出的精油具有杀虫、抗菌等药理学活性,喷施3 μmol/L 28-高油菜素内酯能显著增加精油的含量,还能促进天竺葵生长[16]。此外,BRs还能调控一些菌类物质的生长,古巴裸盖菇喷施 0.01 mg/L(22S,23S)-28-高油菜素内酯后菌丝生长速率增加了2~3倍,一些BRs在双孢蘑菇和平菇(糙皮侧耳)工业生产中有应用前景[17]。
1.2 提高抗逆性
BRs除了能调控植物生长以外,还在植物逆境胁迫下发挥着重要的作用。研究表明,BRs能够促进植物DNA、RNA和蛋白质的合成,从而增强酶的活性,提高植物的抗旱性、抗寒性、抗盐性、抗病性以及增强植物的耐高温能力。
1.2.1 抗旱性。研究表明:用一年生刺槐根部浸蘸BRs,可有效提高幼苗移栽至大田的存活率,以浓度为0.2 mg/L时处理效果最好;待到幼苗叶片长出后叶面喷施BRs,可降低干旱胁迫下叶片蒸腾速率,并通过提高叶片含水量、清晨水势、可溶性糖含量、脯氨酸含量及超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)的活性来增强幼苗耐逆性[18]。因此,施用BRs对于干旱半干旱地区造林具有重要的现实意义。DI-31(BRs类似物)可诱导植物的氧化迸发,通过增强干旱胁迫下植物的生物量和叶面积阻碍相对水含量的下降,还可通过调节AREB/ABF转录因子控制的逆境应答基因来控制气孔的开闭,进而增强植物在干旱条件下的适应性[19]。
土壤盐渍化属于非生物胁迫之一,可使作物减产44.4%~65.4%。24-表油菜素内酯可使盐胁迫下豌豆豆荚种子数量增加28%~38%,千粒重增加18%~35%[20]。 董亚茹等[21]研究发现,0.01 μmol/L 24-表油菜素内酯处理可缓解盐胁迫下桑树叶片和根系的过氧化损伤,提高桑树的抗盐性。高浓度的NaCl处理可以延迟植物根尖细胞的有丝分裂,引起染色体异常。24-表油菜素内酯无胁迫条件下会促进细胞分裂,也会引起染色体异常。然而,在盐胁迫下,其作用于植物,可克服染色体畸变[22]。高温胁迫下植物生物量的积累速度降低,0.05 μmol/L 2,4-表油菜素内酯处理可提高水稻种子萌发效果,实现干物质的快速积累,且能通过调节抗氧化物酶(SOD、POD、CAT)的活性缓解高温伤害[23]。
1.2.2 耐温性。BRs可提高植物的耐热性和耐冷性。将24-表油菜素内酯水溶液施于生长12 d的大麦第1片和第2片叶上,3 d后将其置于高温环境(42℃持续3 h)下,结果表明,24-表油菜素内酯处理能改善叶片光系统Ⅱ(PSⅡ)的功能,起到一定的保护作用[24]。芒果果实经10 μmol/L BL处理后,对5℃低温表现出更高的耐受性[25]。
1.2.3 抗病性及耐药性。BL表现出了一定的抗病性,可以抵抗烟草上的花叶病毒(TMV)、细菌病原体丁香假单胞菌(Pst)以及真菌病原菌,增强水稻对稻瘟病和水稻黄单胞菌的抵抗能力,还可以预防黄瓜枯萎病[26]。油菜素内酯能够缓解由于过量使用农药而产生的药害。喷施BRs后,作物蒸腾量下降,减少药剂的进入量,从而缓解杀虫剂、除草剂等农药引起的药害[27]。于高波[28]研究发现,BRs能够诱导谷胱甘肽巯基转移酶基因的表达,加速谷胱甘肽途径对作物体内百菌清的降解。
1.2.4 缓解重金属毒害。目前,缓解和抑制重金属对植物的毒害作用已经成为研究热点。研究发现,外施低浓度的BRs能够显著增加铝胁迫下绿豆根系干重和叶片叶绿素含量,降低绿豆叶片丙二醛含量,从而增强植物对重金属胁迫的耐受性[29]。尹 博等[30]研究表明,外施24-表油菜素内酯可以调控番茄体内铜、铁、锌的化学形态和亚细胞分布水平,降低铜对番茄的生物毒性,从而保证番茄在铜胁迫下正常进行生理生化代谢活动。
1.3 调节激素的平衡
拟南芥突变体是研究基因功能的重要工具。赤霉素矮化拟南芥突变体试验结果表明,油菜素内酯可通过GPA1基因调控赤霉素的合成,可通过促进β-1,3-葡聚糖酶的释放来促进赤霉素矮化拟南芥突变体种子的萌发[31]。Clouse和Kauschmann从拟南芥矮化突变体(cbb1,cbb2和cbb3)中发现油菜素内酯通过调控meri5和TCH4基因的表达,在植物细胞伸长的过程中发挥着重要作用。种子成熟过程伴随着淀粉、蛋白质和油脂等物质的合成积累,种子脱水干燥后生命活动减弱[32]。遗传分析发现,BRs作为信号物质可下调AGL15转录因子的表达,抑制拟南芥种子的成熟[33]。
BRs作为重要的植物激素,与其他激素交叉协调着植物的生长发育过程,其诱导的生理现象也同时受到其他激素相似或者相反的调控。BRs与赤霉素在细胞伸长的响应中具有类似的作用,与生长素在调控植物生长上具有协同作用,与脱落酸和细胞分裂素在多种生理过程中具有相反或拮抗的效果。
一直以来,赤霉素与BRs对植物下胚轴伸长的促进效应被认为是相互独立的。但是,最近的研究证实,赤霉素的这种效应依赖于BRs激活的BZR1转录因子。生长素与BRs之间的关系研究较多,两者能协同调控植物的生长,这可能与同一个目的基因调控信号传导途径有关。BRs促进植物胚轴伸长的启动较生长素缓慢,但在后期的生长中会比生长素更有效。植物的生长往往伴随着植物内源激素的显著变化,不同激素在光照或黑暗下的作用机理也不尽相同[34]。在胚轴伸长试验中,BL表现出很强的活性,而生长素表现的促进作用并不明显;但是将温度提高7℃后,生长素的促进效应是原先的1.8倍。这主要与两者调控路径中的基因有关。温度升高后,bri1突变体的缺陷使得生长素的反应依赖于BL的转导途径[35]。低浓度下,BL能够促进主根的生长和根的向地性弯曲,这种效应是由于BL加强了生长素中转运突变体auxl-7和pin2的活性,从而加强了根对重力的敏感性[36]。此外,外源施加BL可以促进植物下胚轴生长素的极性运输,但并不能在整体上提高生长素的含量。
BRs促进植物生长的特性与生长素相似,大豆第二节间法(BSIB)中,菜豆幼苗喷施BRs后再喷施生长素可以协同促进乙烯的产生;施用顺序颠倒后,这种效应有所加强。BRs还能诱导生长素协调基因AuxIAA、GH3和SAUR-AC的表达。由此可知,BRs能调控自身信号传导途径以及其他激素的生物合成,例如拟南芥中顶端钩的形成[37]。
乙烯与BL在根向地性生长中也表现出一定的协同效应。高浓度的乙烯能够促进根向地性生长,而BL能够增强ACC合酶的活性,进而提高乙烯的生物合成。同时,乙烯还能加强BL条件下根向地性的响应能力[38]。
BRs与脱落酸之间有一定的拮抗关系。长时间的脱落酸处理能增加BZR2/BESI的磷酸化程度,进而影响BRs的信号传导。研究表明,细胞分裂素能促进光形态建成和叶绿体发育,与BRs调控相反,但是均能促进细胞分裂,两者在分子水平上的关系尚需更进一步研究[39]。
2 保鲜作用
BRs能够延长保鲜时间,提高果品质量。无核小蜜橘外施5 mg/L 24-表油菜素内酯后贮藏在温度为12~16℃、湿度为90%~95%的条件下,果品病害的发生率可由18.7%降至4.0%,这可能与H2O2含量的增加和应激相关代谢产物的积累有关[40]。
3 药理活性
BRs除了在植物上具有调节作用外,还被证实对动物具有一定生理活性作用。研究表明,24-表油菜素内酯可显著缓解有毒物质对鲟鱼、黑海鲑鱼、鲤鱼、欧洲鲫鱼和美国银鲤等鱼类的伤害[17]。低浓度的24-表油菜素内酯还能有效增加梨形四膜虫的数量和提高其对外界的适应度。
Khripach等研究发现,用24-表油菜素内酯饲喂小鼠不仅可以降低正常小鼠体内的胆固醇含量,而且可以显著降低高胆固醇小鼠血液中的甘油三酯含量。BRs具有一定降低胆固醇、抗癌以及抗病毒的作用,因而有望成为研制新型心脑血管疾病药物的备选材料。
细胞试验结果表明,BRs(化合物A和B)可以阻止人体前列腺癌细胞的增殖,同时还能促进癌细胞的凋亡[41]。化合物C从褐藻中分离获得,该化合物表现出一定的抗肝癌细胞和结肠癌细胞的作用(IC50为12.38 μmol/L)[17]。 化合物 E 和 F 具有消毒、杀菌、抗病毒的作用,可能成为治疗角膜炎以及疱疹的新药。此外,Ismaili等[42]还发现,BRs具有一定的保护神经作用,可能会改善老年帕金森症状。具有药理活性的BRs结构如图2所示。
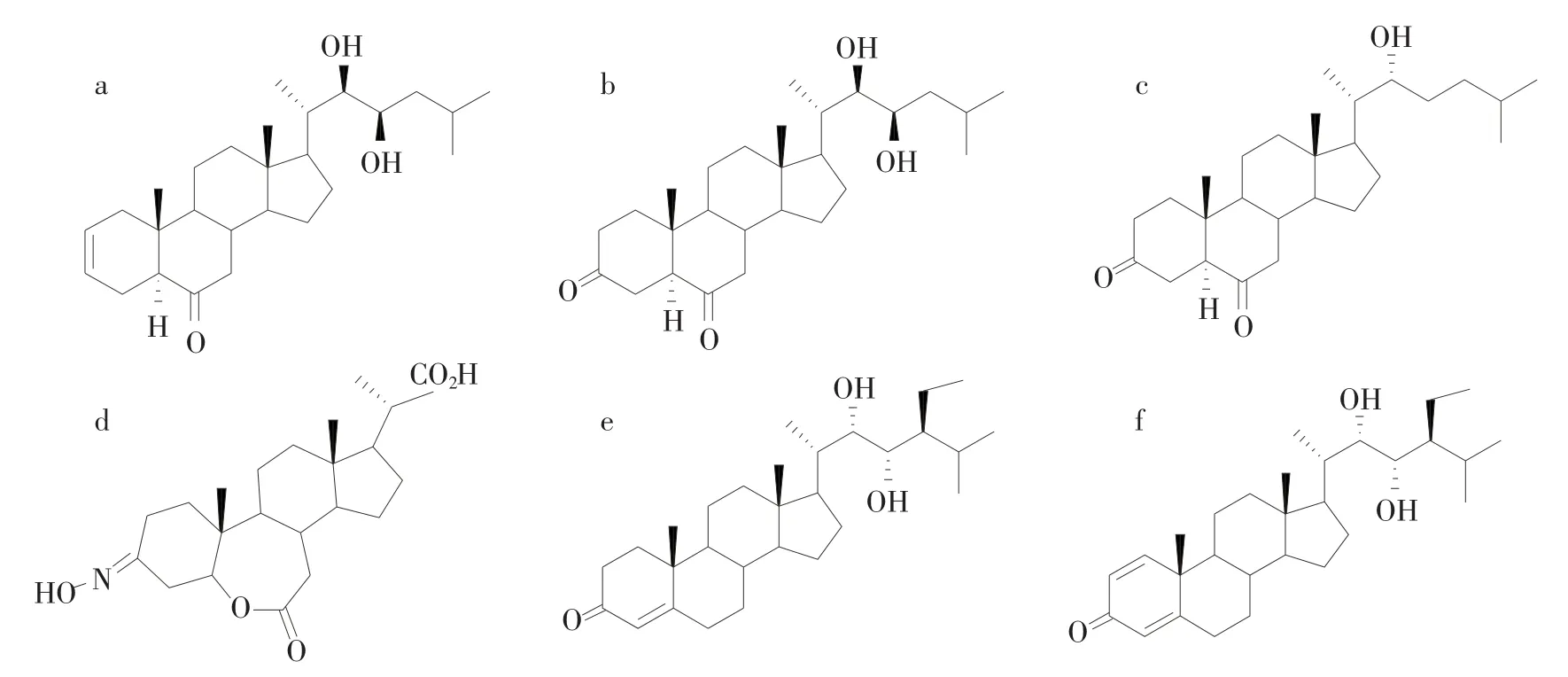
图2 具有药理活性的BRs结构
4 展望
2010—2016年,我国生长调节剂市场需求逐年递增,年均用量为3.65万t,若使用不规范会污染土壤,影响人们健康[43]。BRs作为国际上公认的新型、高效、环保的植物源生长调节剂,已经被开发成多种产品并应用于农业生产中,可使作物增产20%~30%,如芸苔素内酯(brassinolide)、云大 120(28-homobrassinolide)、八仙丰产素(24-epibrassinolide)等。 它们在自然界中含量极低,虽然在花粉和未成熟的种子中含量高,但是也是纳克级别。人工合成是快速获得大量BRs的一个重要途径,但6-氧-7-内酯型BRs合成工艺过程复杂,合成步骤繁琐,产率不高,生产成本极高,合成5 mg的24-表油菜素内酯就需要耗费90美元;加之多羟基结构的存在导致其作用不稳定,大田需要喷施4~5次才能发挥较好的效果。这些因素限制了BRs在农业中的应用[1]。
基于BRs的构效关系研究发现,一些化合物也表现出一定促进植物生长的活性。第一类化合物结构特殊,含有芳香环,表现出了一定的BRs活性,可通过豆甾烷来制备。另一类是螺甾烷类化合物,其结构类似于薯蓣皂苷元,不同的是C6位具有类似栗甾酮的活性官能团,在促进植物生长中表现出稳定性和持效性,能促进生长,增加作物产量,改善作物品质,提高耐逆性,还能阻碍昆虫发育;此外,连接一些糖类物质后还具有一定的抗癌性。最重要的是,该类化合物合成步骤简单,生产成本远远低于6-氧-7-内酯型BRs,有望成为继BRs之后的新型环保植物生长调节剂。
猜你喜欢
中国药学药品知识仓库(2022年5期)2022-04-11
科学导报(2021年33期)2021-06-07
小天使·六年级语数英综合(2020年3期)2020-12-15
三农资讯半月报(2020年14期)2020-08-09
中学生物学(2018年1期)2018-01-19
中国医药导报(2018年28期)2018-01-18
湖北农业科学(2016年24期)2017-03-18
小小说月刊·下半月(2015年11期)2015-05-14
英语学习(2015年12期)2015-02-01
