
凝集素在甲壳类动物免疫学方面的研究进展
2022-10-14 08:30封永辉郭晓萌
中国动物检疫 2022年10期
封永辉,郭晓萌,邱 亮,董 宣
(1.新疆维吾尔自治区水产科学研究所,新疆乌鲁木齐 830000;2.中国水产科学研究院黄海水产研究所,青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室,农业农村部海水养殖病害防治重点实验室,青岛市海水养殖流行病学与生物安保重点实验室,山东青岛 266071)
甲壳类动物作为无脊椎动物中一员,不具备脊椎动物获得性免疫相关的B 淋巴细胞和T 淋巴细胞,但进化形成了完善抵御致病性生物的先天性免疫系统[1],包括由几丁质体表外壳、呼吸器官和消化器官组成的物理防御,血细胞组成的细胞免疫以及由各种免疫因子组成的体液免疫[2-3]。参与体液免疫的许多成分都存储在血淋巴细胞中。甲壳类动物的体液免疫必须依赖血淋巴细胞参与,启动依赖宿主模式识别受体(pattern recognition receptors,PRRs)来识别病原相关分子模式(pathogen-associated molecular patterns,PAMPs)[4-5]。
凝集素作为识别PAMPs 的重要成员,在甲壳类动物细胞免疫识别病原过程中发挥巨大作用[6]。通过甲壳类动物凝集素的深入研究可以提升对甲壳类动物免疫系统的认识水平,对发展绿色健康渔业生产具有重要理论和现实指导意义,有助于从根本上提升渔业生产的防病治病水平。本文从凝集素定义、分类和功能等方面,总结凝集素在甲壳类动物免疫学方面的研究进展情况,并对其中研究较多的两种重要类型——C 型凝集素(C-type lectins,CTLs)和纤维蛋白原样结构域免疫凝集素(fibrinogen-like domain immunolectins,FBGLs)作出总结。
1 凝集素介绍
1.1 定义
1888 年,俄国Hermann Stillmark 研究蓖麻籽毒性时发现提取物可以凝集血细胞,并将该提取物命名为蓖麻蛋白(ricin)。1954 年,Boyd 建议使用拉丁文legere(意为选择)命名,后因其能凝集红血球(含血型物质),将其命名为凝集素。凝集素是从各种植物以及无脊椎动物和高等动物中提纯的一种糖蛋白或结合糖的模式识别分子蛋白。1988年,Barondes 等将凝集素进一步定义为非酶和抗体的结合蛋白[7]。凝集素在自然界广泛存在,植物、动物、微生物中都存在多种类型的凝集素。
1.2 分类及功能
根据物种来源,凝集素被分类为植物凝集素、动物凝集素、原生生物凝集素、细菌凝集素和病毒凝集素。
植物凝集素主要存在于豆科(Legumes)、石蒜科(Amaryllidaceae)、桑科(Moraceae)、觅科(Amaranthaceae)、大戟科(Euphorbiaceae)、茄科(Solanaceae)、唇形科(Labiatae)、尊麻科(Urticaceae)以及谷类(cereals)等植物中。植物凝集素参与植物体抵御病毒和昆虫的免疫过程[8]。
1906 年,在牛血清中首次发现动物凝集素。目前根据其不同结构特征及特异性结合基序,动物凝集素被分为L 型、P 型(配体为甘露糖-6-磷酸盐)、C 型、F 型、M 型、I 型、R 型、S 型(依赖巯基)凝集素以及几丁质酶样凝集素(chitinaselike lectins)、半乳糖苷结合凝集素(galactosidebinding lectins,galectins)、钙连蛋白(calnexin,CNX)或钙网蛋白(calreticulin,CRT)、内凝集素(intelectins)、唾液酸结合凝集素(siglecs)、纤维胶凝蛋白(ficolins)、F-box 凝集素等[9-10]。动物凝集素能识别病原分子物质[11],并调节蛋白折叠和调控细胞信号通路转导等过程。
细菌凝集素分为可溶性凝集素、表面凝集素和菌毛凝集素。在功能方面,细菌凝集素拥有识别作用,与吞噬细胞结合可参与调节吞噬过程等[12],也可参与多种生命活动,如早期胚胎发育和先天免疫。根据分布部位不同,细菌凝集素发挥的作用也不相同。一般胞内凝集素主要用于合成和分泌蛋白,而在胞外或胞膜上的凝集素则可能有参与病原识别和细胞信号转导,触发一系列免疫反应的作用,如引发免疫系统内血淋巴细胞对病原的吞噬及消灭作用。常见的细菌凝集素包括来自革兰氏阴性菌的脂多糖(lipopolysaccharide,LPS),来自革兰氏阳性菌的肽聚糖(peptidoglycans,PGN)和脂磷壁酸(lipoteichoic acid,LTA),来自真菌的甘露聚糖(mannans)、β-1,3-葡聚糖和来自病毒的双链RNA(dsRNA)或糖蛋白[13]。
无脊椎动物体内普遍存在凝集素[14],其发现相对于植物凝集素要晚,相对于动物凝集素要早,可以追溯到1903 年。有研究[14]提到Noguchi 首次在无脊椎动物鲎(Limulus pohyphmus)体液中发现无脊椎动物凝集素,其主要发挥先天免疫功能。甲壳类动物凝集素相对于贝类动物凝集素结构更为复杂,是一种重要的免疫分子蛋白,在体液免疫中发挥识别PAMPs 的作用。甲壳类动物凝集素可以同时参与多项免疫反应,如参与病原识别、结合、凝集,激活酚氧化酶原激活系统(prophenoloxidase activating system,proPO)[15],作为调理素参与细胞吞噬等。Vasta 等[16]研究发现,昆虫、线虫、海胆、七腮鳗、鱼类和两栖类动物中的凝集素主要发挥先天性免疫功能,在细胞间、细胞与胞外基质间相互作用,以及在糖蛋白的转移、蛋白折叠、信号转导、受精和发育中发挥重要作用。
目前在甲壳类动物中研究发现的凝集素包括C 型、L型、M 型、CNX 或CRT、甘露糖苷结合凝集素(mannose-binding lectin,MBL)、galectins、革兰氏阴性菌结合蛋白(gram-negative binding proteins,GNBP)、含硫脂蛋白(thioestercontaining proteins)、清道夫受体(scavenger receptors)、丝氨酸蛋白酶同系物、唐氏综合征细胞粘附分子(down syndrome cell adhesion molecule,Dscam)、FBGLs[9-10,17-20](表1),其中C 型凝集素是甲壳类动物凝集素研究中的典型代表。
2 两种重要甲壳类动物凝集素研究概况
2.1 C型凝集素
C型凝集素是最早发现的动物凝集素之一。1983 年McFaelane 首先发现肝去唾液酸糖蛋白受体(hepatic asialoglycoprotein receptor,ASGPR)。该受体被鉴定为动物C型凝集素。C型凝集素被认为是一类配体依赖钙离子识别糖类型结合蛋白,因凝集需要活性钙离子(Calcium)而得名。它含有一个或多个糖类识别结构域(carbohydrate recognition domain,CRD)或C 型凝集素样结构域(C-type lectin like domain,CTLD),凡是含有与CRD/CTLD 相似的蛋白统称为C 型凝集素[97]。C 型凝集素超家族包括胶原凝素(collectins)、选择素(selectins)、胞吞作用受体(endocytic receptors)和蛋白聚糖(proteoglycans)。其主要特征是含有大约几十到几百个氨基酸序列。CRD/CTLD 结构域数目对其系统进化起了重要作用[9]。C型凝集素大多数是保守的,只包含1 个CRD;通过晶体分析揭示,CRD/CTLD 是一个特殊的压缩球状结构。目前研究发现,C 型凝集素也含有多个CRD,有的虽然具有CTLD,但不具备糖类结合能力。C型凝集素种类丰富,目前有1 000 多个。在脊椎动物中,根据结构和功能将C型凝集素分为Ⅰ—ⅩⅦ共17 组,几乎在所有的多细胞生物中都能被发现[98]。
甲壳类动物C型凝集素组织分布具有物种和组织表达特异性(表1)。多方面研究表明,表达最丰富的组织是肝胰腺、血细胞、鳃、胃、肠道、心脏等免疫相关组织,但在不同病原刺激下,表达量有很大差异,如在凡纳滨对虾(L.vannamei)中发现的LvLT 和斑节对虾(P.monodon)中发现的PmAV 和PmLT,在肝胰腺、血细胞中表达量较多[35,44-45],LvCTL 在胃、肠道中少量表达;在中国明对虾(F.chinensis)中发现的FcLec 仅表达于血细胞[38],FcLec1、FcLec2[33,39]在 胃、肠道中少量表达;在克氏原螯虾(P.clarkii)中提取的PcLec1 在鳃中的表达水平最高,而在血细胞中则非常低[63];日本沼虾(M.nipponensis)中的MnCTLDcp1 和MnCTLDcp2 在心脏中表达量最高,而MnCTLDcp3 在肝胰腺中高表达;罗氏沼虾(M.rosenbergii)中的MrLec3 在肝胰腺、鳃、胃和肠中表达量最高,MrCTL 和MrERGIC-53 在血细胞、肝胰腺、肠道、胃中的表达量最高[21]。阎德平等[80]对三疣梭子蟹(P.trituberculatus)C 型凝集素PtCTL5 研究发现,其在血细胞中的表达量最高,肝胰腺中次之。从不同组织中发现,C 型凝集素发挥着不同的功能,如在凡纳滨对虾(L.vannamei)中,参与了对虾神经系统发育的LvCTLD、LvLec在脑神经节中表达水平远高于在肝胰腺和血细胞中,分析认为其可能在维持脑稳态中发挥着作用,而LvLec1 和LvLec2 却在肝胰腺、肌肉、生殖腺、血液和鳃中都能表达[99]。王敏等[25]对秀丽白虾(E.modestus)研究发现,EmCTL-1 主要在血液中表达,EmCTL-2 主要在性腺、血液和胸神经节中表达,而EmCTL-3 和EmCTL-4 则主要在精巢中表达。中华绒螯蟹(E.sinensis)的EsLecA 主要在精巢和胸神经节中表达,而EsLecG 则主要在胸神经节中表达[70]。
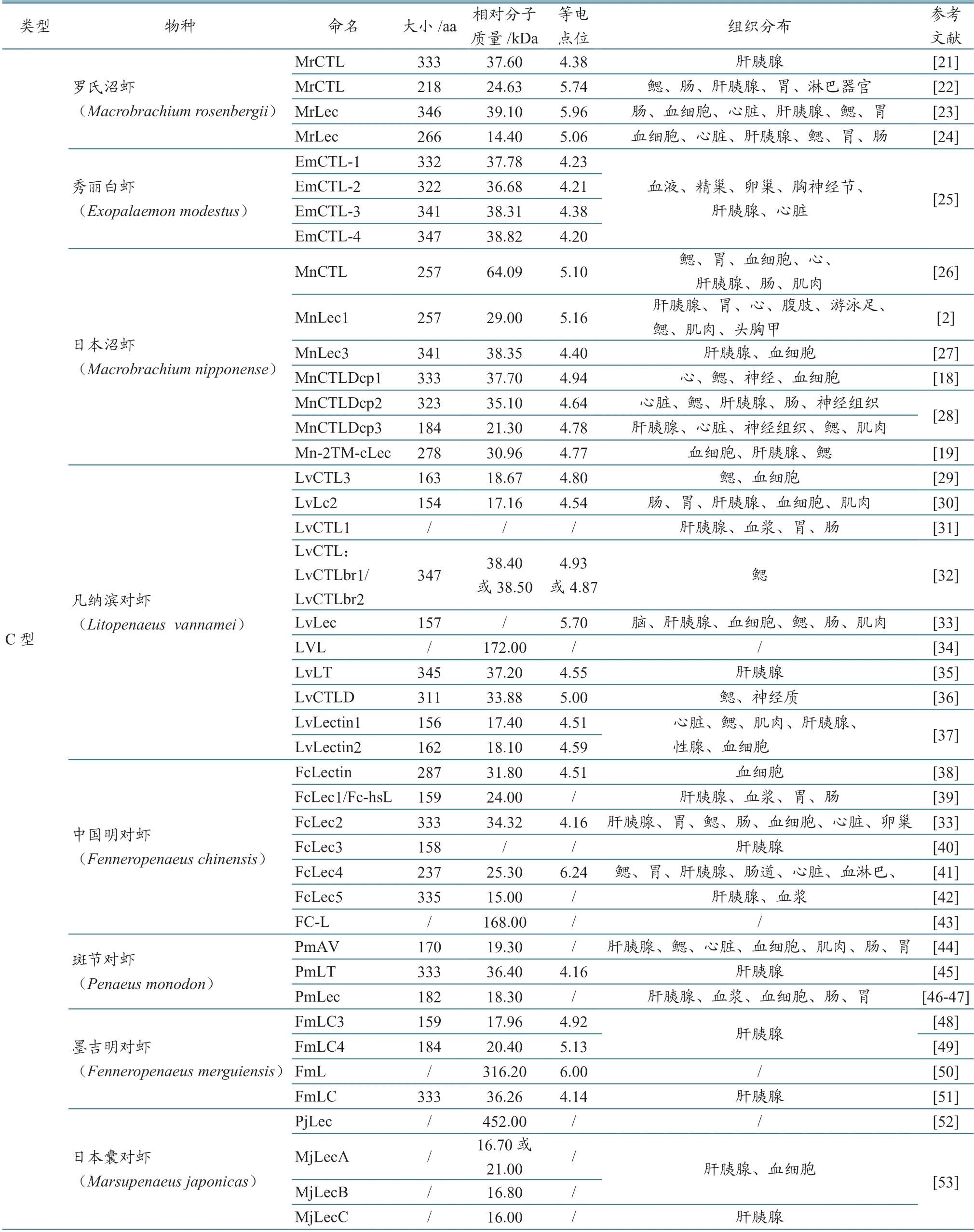
表1 甲壳类动物凝集素种类及蛋白分子大小

表1 (续)

表1 (续)
研究发现,甲壳类动物C型凝集素除作为PRRs 外,还同时在免疫反应中发挥多种免疫功能,例如凝集病原微生物(AMP)[100]、抗病毒应答[31,35,44]、粘附细胞[101]、增强调理、激活酚氧化酶原激活系统、形成结节、参与细胞吞噬作用和包封作用(encapsulation)[9]等。在凡纳滨对虾(L.vannamei)中发现的LvCTLD和在斑节对虾(P.monodon)中提取的PmLT 可增强细胞包被反应,而LvCTLD 甚至可激活黑化作用[45,66]。在日本囊对虾(M.japonicus)中分离出的MjCTL、凡纳滨对虾(L.vannamei)中的LvCTL1、斑节对虾(P.monodon)中的PmAV,可以抑制病毒繁殖并引发细胞病变反应,同时还可参与抗病毒免疫,但研究发现其对白斑综合征病毒(WSSV)不起作用[53,102]。在墨吉明对虾(F.merguiensis)中分离的FmL 可增强抗菌活性[50]。冯锦玲等[23]在研究罗氏沼 虾(M.rosenbergii)中发现,MrLec 可以结合脂多糖和肽聚糖,参与抗病毒和抗菌反应。凡纳滨对虾(L.vannamei)、南美蓝对虾(P.stylirostris)、远洋梭子蟹(Portunus pelagicus)、中华绒螯蟹(E.sinensis)和拟穴青蟹(S.paramamosain)中的几种C 型凝集素缺少信号肽,说明它们可能在胞内发挥作用[36,56,103]。在凡纳滨对虾(L.vannamei)中共分离出LvCTL1、LvCLT3、LvLec 共3 种C 型凝集素,其中LvCTL1 具有抗病毒功能[31],而LvCTL3 可显著降低感染副溶血弧菌和WSSV 引起的死亡率[104],LvLec 则参与革兰氏阴性菌的免疫应答[99]。中国明对虾(F.chinensis)体内的FcLectin和Fc-hsL 具有2 个CRD[38-39];FcLec3 可与WSSV的衣壳蛋白VP28 结合,且具有抑制细菌及真菌的作用[40]。斑节对虾(P.monodon)中的PmAV 具有1 个典型的CRD,显示出抗病毒活性,但不具备凝集活性[44];FcLec4 作为调理素,可促进吞噬作用,清除弧菌[41];Fc-hsL 对一些细菌和真菌具有直接灭杀功能[39]。在日本囊对虾(M.japonicus)中获得3个C型凝集素(MjLecA、MjLecB 和MjLecC),这些凝集素可以结合囊膜蛋白,阻止WSSV 对血细胞的感染,从而降低WSSV 感染率,其中两种还可以体外降低感染效率[53]。在墨吉明对虾(F.merguiensis)中克隆得到1 个双CRD 凝集素FmLec,在受到WSSV 刺激后表达上调。在克氏原螯虾中发现,PcLec-1、PcLec-2、PcLec-3凝集素,可促进虾对鳗弧菌的清除,还可参与机体免疫反应,抵御细菌、病毒、寄生虫等病原体感染[63,105]。
三疣梭子蟹(P.trituberculatus)中也克隆得到一种C 型凝集素[79]。阎德平等[80]在人工感染试验中发现,CTL5 具有消除副溶血弧菌和WSSV 两种病原的作用,且发挥一定的免疫应答功能。Qiu等[69]在体外试验研究拟穴青蟹时发现,SpCTL6不能直接杀灭溶藻弧菌,但是能够明显降低溶藻弧菌内的毒素水平,从而减少因感染溶藻弧菌引起的死亡,提高蟹的成活率。
2.2 FBGLs 凝集素
FBGLs 是一类进化上非常保守的模式识别蛋白,主要包括纤维胶凝蛋白、tachylectins 和纤维蛋白原相关蛋白(fibrinogen-related proteins)。其主要参与非己识别和宿主免疫应答作用[106]。
在淡水螯虾(又名信号克氏原螯虾、淡水小龙虾,P.leniusculus)中发现两种胶原样凝集素蛋白,它们不仅可以参与细菌结合还对革兰氏阴性菌具有清除作用[86]。日本囊对虾(M.japonicus)中发现的MjFREP1 对革兰氏阳性菌具有凝集作用,同时能够结合肽聚糖、脂多糖以及WSSV 的VP28 蛋白,在抗细菌和抗病毒反应中发挥重要作用[88]。罗氏沼虾(M.rosenbergii)的两种纤维胶凝蛋白以及Dai 等[90]在克氏原螯虾体内鉴定的PcFLP1,均能够凝集细菌,增强虾体对细菌的清除能力,在抗弧菌免疫中发挥关键作用。
3 讨论
在美国国家生物技术信息中心(NCBI)数据库中检索到的中国明对虾(F.chinensis)凝集素数据共43 条,其中有已发表论文支撑的只有10 条,其他数据均为软件预测的功能结果,需要进一步开展试验研究来挖掘其相关的功能。
在信号通路的研究中,针对甲壳类动物虽然也开展了一些工作,如中国明对虾(F.chinensis)血细胞的酚氧化酶原系统包括脱颗粒和酚氧化酶(phenoloxidase,PO)的活性,是通过磷酸激酶C(phosphokinase C,PKC)通路来调节的[107];罗氏沼虾(M.rosenbergii)血细胞proPO 系统包括脱颗粒和PO 的活性,是ODN2006 通过PKC、TPK 信号通路来调节,且此过程与磷酸激酶A(phosphokinase A,PKA)信号通路无关[107];凡纳滨对虾(L.vannamei)中LvCTL4 能通过激活NF-KB,随信号通路参与抗菌免疫反应[108],但是否同一物种同时存在着多条类似脊椎动物的信号途径,如免疫缺陷同系物(IMD)途径、JAK/STAT途径、Toll 样受体(Toll-like receptors,TLRs)信号途径,仍缺乏系统性的研究数据来证明。从目前的报道来看,甲壳类动物信号途径的整体性及调控机制还不明确,如在凡纳滨对虾(L.vannamei)中,已报道的9 种C 型凝集素如何发挥免疫学调控作用,9 种凝集素是否具有协同或抑制机制,这些都需要进一步开展相关试验研究。
甲壳类动物凝集素的研究结果还需进一步整合,不同研究在同一物种分离到的凝集素是否是同一种物质,还需要进一步确认。如在罗氏沼虾(M.rosenbergii)研究中,Snigdha 等[21]、Huang 等[22]发现的MrCTL,以及Feng 等[23]和Huang 等[24]发现的MrLec 具有不同的氨基酸数目和相对分子质量,其是同一种功能物质,还是两种不同的物质,需要进一步比较确认。
4 结语
目前对甲壳类动物的免疫机制,主要以经济品种对虾、龙虾、鳌虾和蟹等动物为研究对象,取得了很多成果,在一定程度上揭示了甲壳类动物尤其是十足目动物免疫防御机制。然而受试验材料及研究手段的限制,甲壳类动物的免疫机制研究仍然滞后于其他无脊椎动物,处于相对初级阶段,对于病原体致病机制及虾蟹先天性免疫防疫机制的研究仍然具有很大的提升空间。而对先天性免疫认识水平的提高,可以在促进虾蟹自身的免疫能力方面,为疾病防治提供新的方法,从根本上提升防病治病水平。
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2022年8期)2022-09-20
当代水产(2022年5期)2022-06-05
中国药学药品知识仓库(2022年9期)2022-05-23
锦州医科大学报(2022年2期)2022-05-07
家庭科学·新健康(2022年1期)2022-02-02
百科探秘·海底世界(2020年4期)2020-04-24
食品与生活(2018年7期)2018-09-19
漫画月刊·哈版(2017年7期)2018-01-29
山东工业技术(2017年5期)2017-03-16
