
基于积分投影模型预测濒危植物景东翅子树的种群动态
2022-10-13 05:14袁春明杨国平耿云芬张珊珊
生态环境学报 2022年8期
袁春明,杨国平,耿云芬,张珊珊
1.云南省林业和草原科学院,云南 昆明 650204;2.国家林业和草原局云南珍稀濒特森林植物保护和繁育重点实验室,云南 昆明 650204;3.云南省森林植物培育与利用重点实验室,云南 昆明 650204;4.中国科学院西双版纳热带植物园,云南 勐腊 666303
种群动态模型在回答有关生态、进化和保护有关的问题方面是一个有价值的工具。虽然可以从个体数量或种群密度估计种群的趋势,但了解驱动这些趋势的机制需要对基本生命率或种群统计过程(生长、存活和繁殖)进行量化(Metcalf et al.,2013)。种群的矩阵投影模型,由于其结构相对简单且能提供有价值的信息,已成为种群动态分析的主要方法(Caswell,2001)。种群矩阵模型的普及应用在20世纪80年代得到了快速的发展,同期引入的弹性分析(Caswell,1984;de Kroon et al.,1986),至90年代已成为种群统计学和保护生物学中扰动分析的主要方法(Crouse et al.,1987;Silvertown et al.,1993)。
种群矩阵模型包括基于年龄的Leslie矩阵模型(Leslie,1945)和基于大小或生活史阶段的Lefkovitch矩阵模型(Lefkovitch,1965),因为人为划分的生活史阶段的数量和投影矩阵的维数的不同而影响到种群统计的结果(Enright et al.,1995;Ramula et al.,2005)。另外,矩阵模型中种群生活史被划分成离散的阶段,而划分生活史阶段的变量往往是连续的(如年龄、大小等)。这些缺陷对生长缓慢、寿命长的树种来说更是如此。为此,Easterling et al.(2000)提出了积分投影模型(Integral projection models),该模型无需对所有个体划分生活史阶段,而是通过反映个体的状态变量如大小变化、生存和繁殖的连续函数来探讨种群的动态。在此基础上,Zuidema et al.(2010)提出了一种新的积分投影模型的参数化方法并进行了比较验证,认为积分投影模型在分析树木的种群统计是十分合适的工具。积分投影模型功能强大,能够整合连续的和离散的变量(如种子库)、环境的变量,以及应用于具有复杂生活史的植物种群等,它们已经得到了迅速的推广应用(Ellner et al.,2006;Dahlgren et al.,2009;Miller et al.,2012;Dauer et al.,2013;Li et al.,2013;Zambrano et al.,2014)。
景东翅子树(Pterospermum kingtungense)属梧桐科翅子树属,为云南特有种,其分布区域狭窄、种群数量极少,按照IUCN濒危等级标准评价为“极危种”,被国家林业局列为国家重点保护野生植物名录(第一、二批)中二级保护植物及全国极小种群野生保护植物。根据第二次国家野生保护植物调查,该物种有12个空间或地理上分隔的种群,分布在不同大小的片断或连续森林中。本研究基于不同大小(5 hm2和15 hm2)片断和连续森林中景东翅子树种群统计参数的连续3年调查及实验研究,应用积分投影模型预测该濒危物种的种群动态。主要回答以下2个问题:(1)不同大小的片断和连续森林中景东翅子树的种群动态如何?(2)驱动景东翅子树种群动态变化的关键因子是什么?目的是为该濒危物种的保护和管理策略的制定提供科学依据。
1 材料与方法
1.1 研究地点与物种概况
研究地点位于云南景东彝族自治县川河流域。研究区域属典型山地季风气候,年平均气温18.3 ℃,最冷月平均气温 10.9 ℃,最热月平均气温23.2 ℃;年平均降水量1086.7 mm,5—10月为雨季,其降水量占全年降水的84.1%。研究区域自然植被主要为松林和山地常绿阔叶林。常绿阔叶林主要分布在山体的下部,由于人为干扰破坏如开垦、修路等,造成了这些常绿阔叶林的片断化。
景东翅子树为乔木树种、高达25 m,具两性花,花期1—3月,果期10—11月。景东翅子树生长缓慢、寿命长,属所在群落演替后期的物种,主要分布在海拔1400 m左右的山地常绿阔叶林中。
1.2 样地设置与调查
选择3种类型的生境,包括5 hm2和15 hm2的森林片断各1个,以及2个大片的连续森林。在每一种生境中分别建立3个50 m×100 m的固定样地(共9个),其中片断生境中的样地尽可能靠近片断中心,连续森林中的样地则设置在远离森林边缘的位置。在样方四角埋设PV管固定样方位置,以便进行种群动态研究的连续调查。
2018年1—2月对样地内景东翅子树的所有个体进行全面调查并挂牌编号,记录胸径(DBH,位置用油漆进行标注)和高度,其中幼苗和幼树测定其地径。2018年繁殖季节调查样地内所有个体是否开花结实,对开花个体调查其单株果实数、每果实的种子数等繁殖产出参数。
2019、2020和2021年重复进行上述调查,对每一个体跟踪记录其生长状况的变化如胸径(或地径)、高度,是否开花结实、是否死亡,对开花个体调查其繁殖产出参数(单株果实数、每果实的种子数),以及新增幼苗个体数。样地概况及调查详见Yuan et al.(2021)及杨国平等(2021)。
1.3 积分投影模型(IPM)的建立
IPM对种群动态的表述与矩阵投影模型相似,不同的是其中的转移矩阵由一个积分函数来代替(Easterling et al.,2000;Metcalf et al.,2013):

式中:
n(y,t+1)和n(x,t)——时间t+1和t时的种群个体大小的分布;
L和U——IPM模型中个体大小的下限和上限(本研究中将胸径或基径的大小作为个体的状态变量);模型核k(y,x),可分解为存活生长P(y,x)和繁殖F(y,x)二部分:
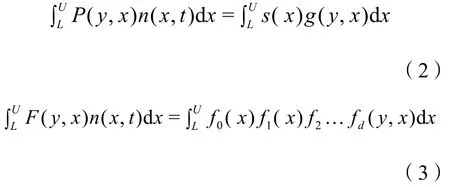
s(x)——存活概率;
g(y,x)——生长(时间t+1时的个体大小,即大小为x的个体成长为大小为y的个体);
f0(x)——开花概率;
f1(x)——每株果实数;
f2——每果实的种子数(常量);
…fd(y,x)——代表与繁殖有关的其它函数如幼苗建立的百分比、幼苗的生存率等。其中存活概率s(x)、开花概率 f0(x)通过逻辑回归建立、生长即时间t+1时的个体大小g(y,x)通过线性回归建立、每株果实数f1(x)通过泊松回归建立,其它常量函数如每果实的种子数、幼苗建立的百分比、幼苗的生存率等则通过野外调查或试验取得,其中部分结果见Yuan et al.(2021)。
1.4 数据分析
景东翅子树为极小种群物种,个体数量较少,参照杨国平等(2021)依据种群中每个个体的跟踪调查相关资料,建立3种不同生境(5 hm2片断、15 hm2片断和连续生境)3个时间段(2018—2019、2019—2020、2020—2021年间)的综合IPM模型9个,所谓综合种群模型是指将同一生境如连续生境中的3个样方的数据合并在一起而建立的模型;生命率回归模型的筛选采用AIC即赤池信息量准则(Akaike information criterion),它是衡量统计模型拟合优良性的一种标准,取AIC值最低者;运用中点法则将模型核 k(y,x)转化为100阶大转移矩阵(Easterling et al.,2000;Metcalf et al.,2013),计算种群增长率、弹性值等。数据处理使用R软件(v.3.4.3),调用R 共享包 IPMpack(v.2.1;Metcalf et al.,2013)编制的R代码进行修订实现。
2 结果与分析
2.1 种群生命率
3个时间段不同大小片断和连续森林中景东翅子树的种群生命率回归模型及参数如表1。从图1—3可知,3种生境中景东翅子树死亡个体数为0—2 plant·a-1·hm-2,年开花个体数为0—4 plant·a-1·hm-2,新增个体数为0—1.3 plant·a-1·hm-2(新增个体数及每株果实数函数未显示)。2020—2021年间5 hm2片断种群存活概率随个体大小的增加而减少,其它种群均表现为随个体大小的增加而增加;种群的开花率除2018—2019年和2019—2020年间5 hm2片断未出现开花外也随个体大小的增加而增加。

表1 不同生境中景东翅子树的种群生命率回归模型及参数Table 1 Vital rate regression models and its parameters of P.kingtungense populations in different habitats
2.2 种群增长率
15 hm2片断和连续森林中景东翅子树种群的增长率等于或趋近于1(表2),说明其种群是稳定的,其中15 hm2片断种群的增长率3个时间段均≥1表明种群是稳定或增长的,连续森林中种群的增长率在2018—2019年和2019—2020年间略有下降,而在2020—2021年间以每年0.036%的速度呈几何级数增长。5 hm2片断森林中景东翅子树种群的增长率在2018—2019年和2018—2019年间趋近于1,但在2020—2021年间以每年9.133%的速度呈几何级数减少。

表2 不同生境中景东翅子树种群增长率Table 2 Population growth rate of P.kingtungense in different habitats
2.3 弹性分析
3个时间段不同大小片断和连续森林中景东翅子树种群的存活和生长对种群增长率的贡献占99.2%以上,而繁殖对种群增长率的贡献不超过1%(表3)。景东翅子树种群的存活和生长对种群增长率的贡献占绝对优势,说明种群的存活和生长是驱动种群增长、维持种群稳定的关键因子。
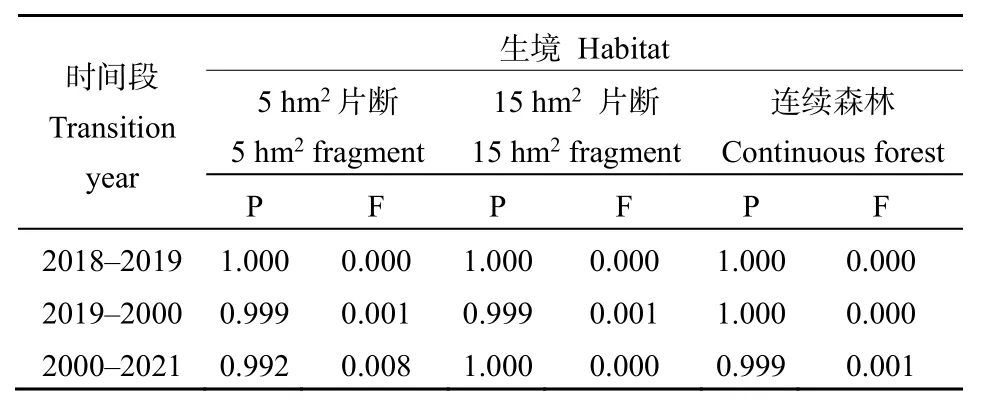
表3 不同生境中景东翅子树种群生命率参数的弹性值Table 3 Elasticity values of vital rate of P.kingtungense populations in different habitats
3 讨论
本研究表明,不同生境不同时间段景东翅子树种群生命率(或统计过程)存在一定差异。3种生境中景东翅子树死亡个体数一般为0—2 plant·a-1·hm-2,其中死亡的个体主要为幼苗或幼树,这主要是由于干旱造成的;只有一个例外,即5 hm2片断2020—2021年间的1个样方中死亡了3株(图3a)、是因维护电线线路由人为砍伐造成的。不同生境中景东翅子树年开花个体数为0—4 plant·a-1·hm-2,15 hm2片断中2018—2019年间3个样方中开花个体数最多为6株(图1h),5 hm2片断中 2018—2019、2019—2020年间未发现开花现象,2020—2021年间3个样方中只有1个个体开花(图1g、2g、3g)。5 hm2片断中景东翅子树种群数量及大树少,降低了景东翅子树的开花率、甚至造成多年才开花的现象,因而导致幼苗补充率下降(Yuan et al.,2021)。15 hm2片断和连续森林中幼苗的补充限制则可能受微生境因子(如森林枯落物、森林动物及真菌病原体对种子的侵害等)的影响。

图1 2018—2019年间不同生境中景东翅子树种群生命率函数Figure 1 Vital rates’ functions of P.kingtungense populations in different habitats for transition year 2018-2019

图2 2019—2020年间不同生境中景东翅子树种群生命率函数Figure 2 Vital rates’ functions of P.kingtungense populations in different habitats for transition year 2019-2020

图3 2020—2021年间不同生境中景东翅子树种群生命率函数Figure 3 Vital rates’ functions of P.kingtungense populations in different habitats for transition year 2020-2021
积分投影模型揭示出景东翅子树种群如果没有较大的人为干扰和环境的波动(如严重的干旱等),3种生境下景东翅子树种群可维持稳定。此结果与应用Lefkovitch矩阵投影模型对该研究物种预测的结果相似(杨国平等,2021),与墨西哥东南部韦拉克鲁斯州的Los Tuxtlas森林中一个演替后期的树种预测结果也一致,即生境片断化并没有降低所研究物种的种群增长率(Zambrano et al.,2014)。15 hm2片断和连续森林中景东翅子树种群的增长率等于或趋近于1(表2),说明其种群是稳定的,这是因为景东翅子树种群的死亡率和幼苗的补充率均较低(分别为0—2 plant·a-1·hm-2和0—1.3 plant·a-1·hm-2),二者相互抵消使种群处于相对平衡状态。5 hm2片断景东翅子树种群动态在年际间是波动的,3年间幼苗未得到补充,相反2018—2019年间3个样方中死亡了2株(图1a)造成种群增长率的下降,2019—2020年间未发现死亡植株(图2a)其种群维持稳定,2020—2021年间1个样方中人为砍伐了3株(图3a;个体胸径分别为7.5、6.7和46.3 cm,其中前2个个体大小相近在图中重合为1个点),造成种群增长率的较大下降。
弹性分析表明,景东翅子树种群的存活和生长是驱动种群增长、维持种群稳定的关键因子。弹性分析主要用于评价和比较种群特定生活史阶段生存、生长和繁殖等种群参数对种群增长率的相对贡献(de Kroon et al.,2000;Heppell et al.,2000)。3个时间段不同大小片断和连续森林中景东翅子树种群的生长和存活对种群增长率的贡献均占绝对优势、繁殖的贡献不到1%(表3),这可能是因为观测的3年期间正处在景东翅子树结实小年的原因,并且幼苗建立的百分比也较低(0.091%—0.405%)。
濒危物种一般具有个体数量少、繁殖率低等特点。积分投影模型能够更好地利用较少的数据(Ramula et al.,2009),通过生命率回归模型预测濒危物种种群的动态,相比矩阵投影模型具有较大的优势。但是就本研究来说,为期3年对景东翅子树种群动态观测的时间还是短了一些。景东翅子树为极小种群物种,其繁殖力较低。项目研究开始前的2017年为结实丰年,而3年观测研究期间还未见第2个结实丰年的出现;项目开展前期还认为5 hm2小片断生境的个体不开花,观测到第3年(2020年)才发现1株树开花结实。这些都说明对濒危物种种群动态的研究应进行较长时期的定位监测才能获取“理想”的结果。
4 结论
3年期间景东翅子树种群生命率(或统计过程)在不同时段不同生境中存在一定差异,但是除 5 hm2片断在2020—2021年间受人为影响导致较大下降外,3种生境中景东翅子树种群的增长率等于或趋近于1,说明其种群是稳定的。3种生境中景东翅子树种群的存活和生长对种群增长率的贡献占99.2%以上,说明种群的存活和生长是驱动其种群增长、维持种群稳定的关键因子。研究认为尽管景东翅子树种群个体数量少繁殖力较低,但是如果没有较大的人为干扰和环境的波动如严重干旱等,3种生境中景东翅子树这种长寿命物种的种群可维持稳定。因此,除去人为干扰外,对于这些濒危物种的保护关键在于对其生境的保护以维持系统抵抗环境扰动的能力。
猜你喜欢
农业与技术(2022年15期)2022-08-15
东北林业大学学报(2022年1期)2022-03-28
种子(2022年2期)2022-03-21
风景园林(2022年1期)2022-03-07
风景园林(2022年1期)2022-03-07
意林·少年版(2020年17期)2020-10-12
发明与创新·中学生(2020年4期)2020-04-17
农民致富之友(2019年11期)2019-05-23
环球时报(2017-01-26)2017-01-26
海外星云 (2014年21期)2015-01-14
