
22种枫属植物花粉形态及其分类学意义
2022-10-10 00:23祝志勇
福建林业科技 2022年3期
林 立,祝志勇
(1.宁波城市职业技术学院、宁波市园林植物开发重点实验室,浙江 宁波 315100;2.南京林业大学生物与环境学院,江苏 南京 210037)
花粉在植物生长发育过程中受环境因素影响小,常被用于植物科、属、种甚至种下分类和系统学研究[1-2]。尽管也有研究表明花粉形态在植物分类中存在局限性[3],但更多研究证实花粉形态在植物种属鉴定、种间(种下)关系推断中具有重要作用[4]。花粉许多重要特征,如大小、形状、外壁纹饰、萌发孔类型等,对于推断植物的起源、演化也具有重要意义[5-7]。
近年来,随着显微技术的快速发展,关于花粉形态的研究也越来越多。武海霞等[5]观察了20个桉树(Eucalyptusspecies)样品的花粉结构,指出花粉形态可作为桉树种间鉴别、杂交亲子代关系推断的重要指标。陈常颂等[6]发现不同茶树品种(Camelliasinensiscultivars)的花粉外壁纹饰能够稳定遗传,可作为种下分类依据。洪欣等[7]研究了报春苣苔属(Primulina)花粉形态,证实了将该属作为广义报春苣苔属的合理性。据统计,我国裸子植物、被子植物中分别已有三分之二、三分之一的科进行过花粉形态学研究[8]。
枫属(AcerLinn.)是一个分类和系统发育研究十分困难的属[9-12]。前人基于形态特征[10-12]、遗迹化石[13]、同工酶[14]、分子标记[15-16]等方面数据对枫属的分类和系统发育进行了大量研究,但该属的系统发育、属下分类以及部分种的系统位置仍未完全明确。2008年,徐廷志等在《Flora of China》(FOC)中对我国枫属进行了较大程度修订,将该属修订为14组99种[11]。但是,该系统还未得到有效验证,并且许多种的归并尚存在争议[17]。
国内外许多学者开展过枫属孢粉学研究[18-21]。Erdtman观察到枫属花粉粒具3沟或3拟孔沟[18]。几濑则发现枫属16种和2变种的花粉粒都为3孔沟型[19]。我国1960年出版的《中国植物花粉形态(第一版)》记载了3种枫属植物的花粉粒具三沟[20]。此后,王伏雄等[21]在增订版《中国植物花粉形态(第二版)》中记录了金钱枫(Dipteroniasinensis)为三孔沟型花粉。Biesboer[22]利用扫描电镜观察了41种枫树的花粉形态,发现花粉粒的表面雕纹在各组间具有差异性。Pozhidaev[23]用同样方法观察了枫属31种的花粉形态,发现花粉粒的孔径排列非常规律。张静敏等[24]观察到枫属花粉粒外壁多为条纹状雕纹,发现只有复叶枫具网状雕饰。曹伟等[25]观察了东北枫属植物的孢粉形态并将花粉分为两种类型。赵先贵等[26]基于花粉外壁特征探讨了枫属组间的亲缘关系。田欣等[27]发现枫属植物的花粉多为3沟型,外壁主要呈条纹状排列。由此可见,枫属花粉的一些特征,如外壁雕纹、萌发孔类型等,可作为属种间分类依据,并且对于枫属系统发育研究有参考意义。
1 材料与方法
1.1 研究材料
供试材料于2020年采自宁波溪口枫树基地、浙江天目山和杭州植物园等地,共22种(亚种)(表1),经中科院昆明植物园周元教授鉴定并借助中国数字植物标本馆(http://www.cvh.org.cn/)核对,种名按照《Flora of China》[11]。争议种如橄榄枫(A.olivaceum)仍根据《中国植物志》[12]命名。花粉材料用采样袋收集后带回实验室进行后续试验。
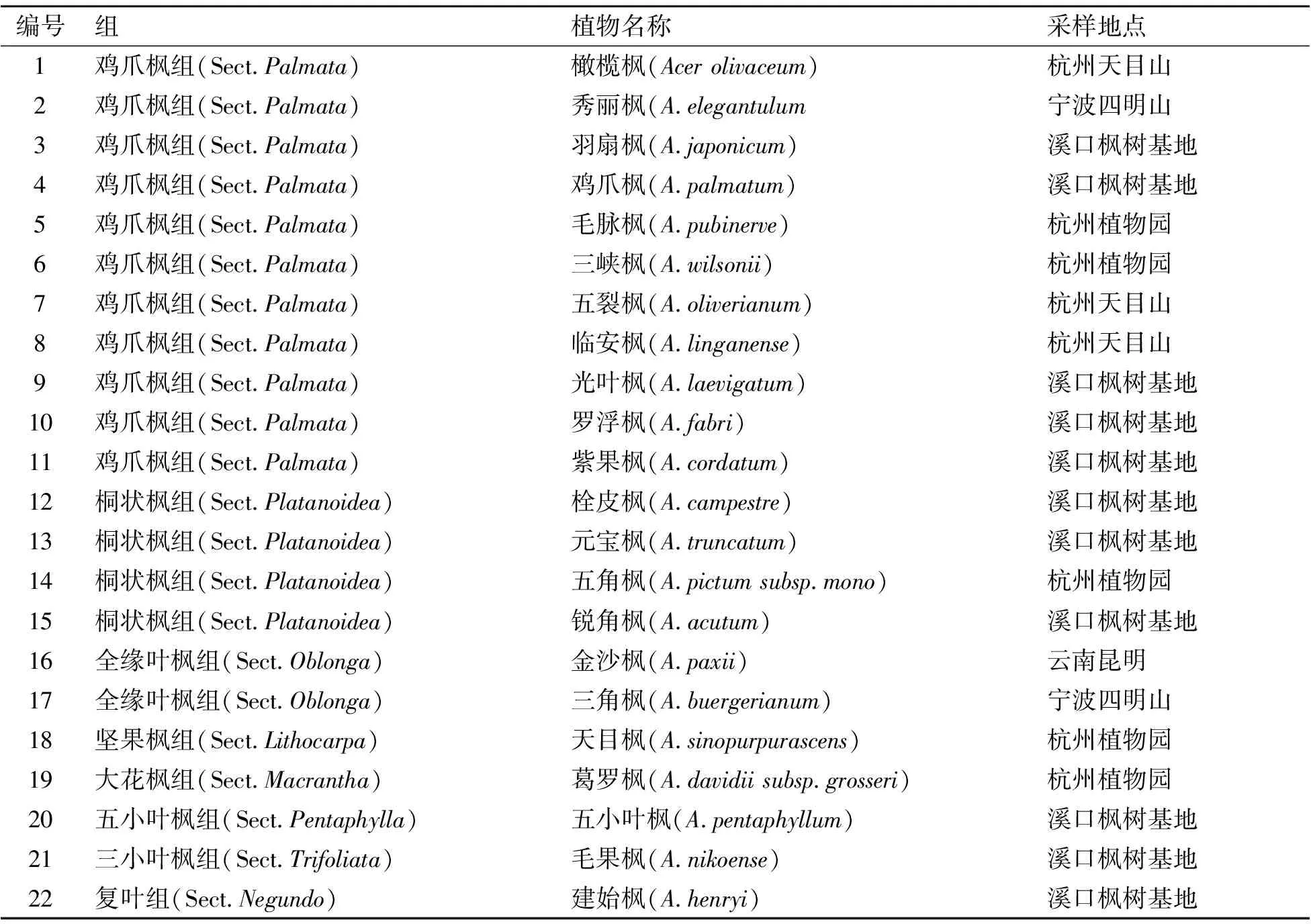
表1 22种枫属植物供试材料
1.2 研究方法
光学显微镜观察:采用Erdtman[1]醋酸酐分解法处理制片。在光学显微镜下测量[L3]枫属各种植物花粉大小,每种花粉取20粒,测最大值、最小值和平均值。
扫描电镜观察:将样品放在XD-1型CO2临界点干燥器(日本Eiko公司)中干燥,抖落花粉后撒在粘有导电胶的样品台上,用洗耳球吹扫粘在表面的样品,真空喷金镀膜后在JSM-840(日本JEOL公司)扫描电镜下观察。
1.3 分析方法
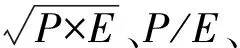
2 结果与分析
2.1 花粉形态特征
22种枫属植物的花粉形态特征见表2、图1。
2.1.1 花粉形状与大小 花粉形状从近球形到超长球形变化,赤道面观为近圆形或椭圆形至窄椭圆形。所有材料中,近球形有5种,分布于枫属4个组;近长球形8种,主要为鸡爪枫组类群(5种);超长球形1种,为鸡爪枫组的三峡枫;长球形种类最多(9种),分布于鸡爪枫组(3种)、桐状枫组(3种)、大花枫组(1种)、坚果枫组(1种)、三小叶枫组(1种)和全缘叶枫组(1种)。
花粉粒极轴长(P)在19.84~38.93 μm之间,其中建始枫极轴最短,毛果枫极轴最长。赤道轴长(E)在14.27~28.44 μm之间,其中葛罗枫赤道轴最短,毛果枫赤道轴最长。所有枫属植物中,葛罗枫花粉粒最小,体积大小指数为17.59;毛果枫最大,体积大小指数为33.35。
2.1.2 萌发孔类型 花粉萌发孔有3沟和3孔沟2种。3孔沟主要出现于鸡爪枫组和大花枫组。22种供试材料中,萌发沟最长的为毛果枫(32.87 μm),最短的为临安枫(14.42 μm),萌发沟长度与极轴长度比值(L/P)在0.55(元宝枫)~0.97(光叶枫)之间。萌发沟沟缘多为不整齐,长可达极面。
2.1.3 外壁纹饰 供试22种枫属植物花粉外壁都为条状纹饰,并且条纹大多平行排列,仅葛罗枫花粉条纹排列较不规则。葛罗枫属大花枫组,田欣等[27]对该组其它类群的观察也发现花粉外壁条纹排列不规则,表明该特征在大花枫组中具有特异性。
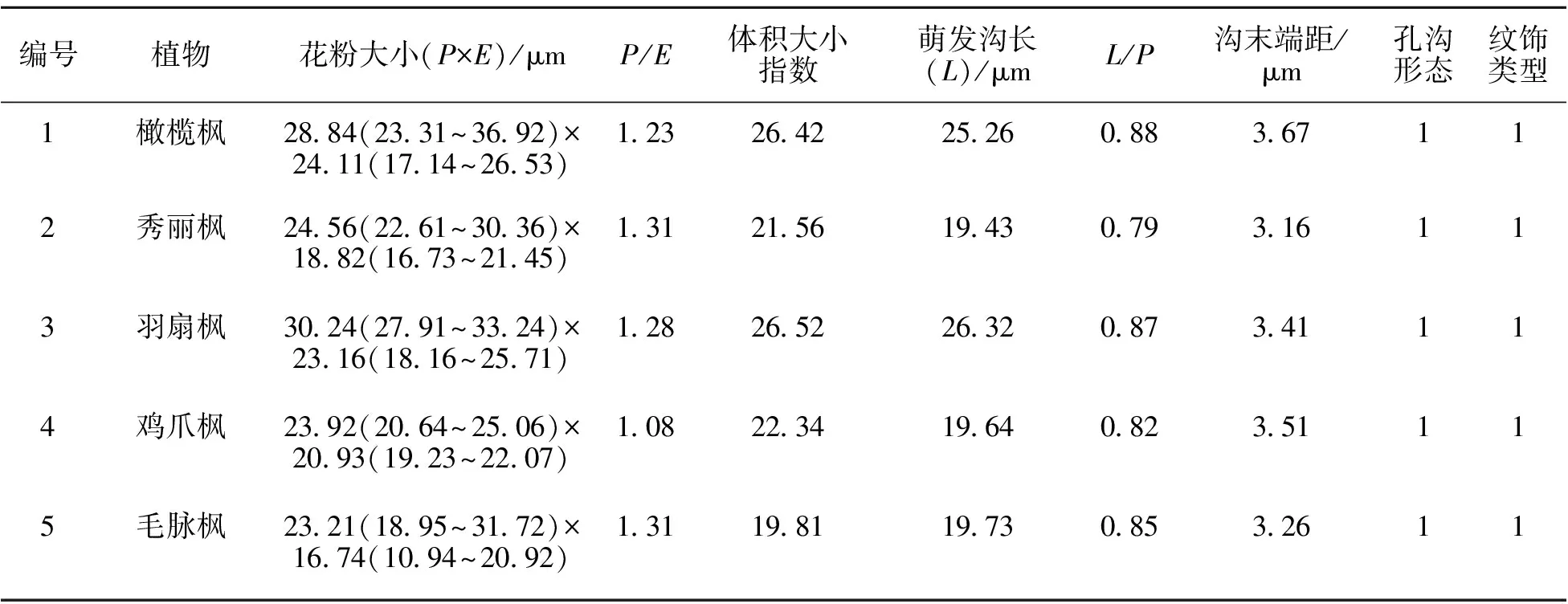
表2 枫属植物花粉特征

表2(续)
2.2 花粉形态的主成分分析
为研究供试花粉的主要特征差异,并获取枫属种间界别的花粉形态证据,将该属22个种(亚种)的花粉作为分析单位,对9个形态指标进行主成分分析(PCA),结果见表3、表4。
PCA结果表明,前4个主成分贡献率分别为36.668%、27.106%、14.081%、11.203%,累计贡献率为89.057%,表明前4个主成分可以代表原始变量绝大部分信息[28]。根据4个主成分得分系数(表4)可知:第1主成分主要受体积指数影响;第2主成分主要受P/E值影响;第3主成分主要受外壁纹饰影响;第4主成分主要受L/P值影响。总而言之,体积指数、P/E值、孔沟形态和沟末端距是影响较大的性状特征。

1为A.olivaceum 6000×;2为A.elegantulum 6000×;3为A.japonicum 6000×;4为A.palmatum 7000×;5为A.pubinerve 8000×;6为A.wilsonii 5000×;7为A.oliverianum 5000×;8为A.linganense 8000×;9为A.davidii subsp.momo 8000×;10为A.laevigatum 6500×;11为A.fabri 5000×;12为A.campestre 5000×;13为A.truncatum 7000×;14为A.acutum 5000×;15为A.paxii 7000×;16为A.buergerianum.6000×;17为A.sinopurpurascens 6000×;18为A.cordatum 8000×;19为A.davidii subsp.grosseri 5000×;20为A.henryi 6000×;21为A.pentaphyllum 7500×;22为A.nikoense 5000×。图1 22种枫属植物的花粉电镜图
2.3 花粉形态的聚类分析
根据PCA结果选择4个主成分指标(体积指数、P/E值、孔沟形态和沟末端距),采用平方欧氏距离法对22种枫属植物进行Q聚类分析。结果(图2)表明,在遗传距离为10时,22种枫属植物被划分为3个类群:类群Ⅰ,包含a、b、c 3个亚类群,共有14种枫树;类群Ⅱ,仅包含亚类群d,有5种枫树;类群Ⅲ,包含e、f 2个亚类群,共有3种枫树。
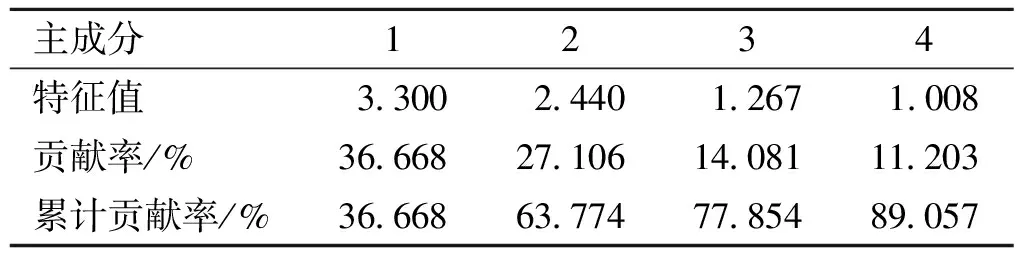
表3 22种枫属植物花粉形态前4个主要成分贡献率
亚类群a共包含4种枫树;其中3种为鸡爪枫组的橄榄枫、羽扇枫和三峡枫,该3种枫树花序类型、花粉形态相似,但其它诸如叶型、果型等特征则差异明显,并且分布区域也不尽相同,可能是应对不同生态条件的适应性辐射的结果。亚类群b包含鸡爪枫组5种、桐状枫组3种及坚果枫组1种;该9种枫树虽然在叶型、株型等方面差异较大,但均生活在温暖潮湿的低海波森林地带,表现出了与高湿度环境的适应性。亚类群c仅有三角枫1种。亚类群d包含5种枫树,分别属于鸡爪枫组(2种)、大花枫组(1种)、全缘叶枫组(1种)和复叶枫组(1种),5种枫树自然分布范围都较广,生境条件较为相似。亚类群e来源于桐状枫组,花粉形状较为相似。亚类群f由三小叶枫组的毛果枫构成,毛果枫花粉粒较大,与花粉粒同样较大的亚类群e构成了类群Ⅲ。可见,由代表花粉形状的4个主成分指标构建的聚类关系不能作为种间区分的标准,但可以在一定程度上反应花粉形态与生境条件的对应关系。

图2 22种枫属植物基于4个花粉形态指标的聚类图
3 讨论
3.1 枫属植物花粉形态特征
供试枫树花粉许多特征具有一致性,如:为辐射对称的单粒花粉;赤道面观为近圆形或椭圆形,极面观呈三裂圆形;萌发沟细长,沟缘多为不整齐,长可达极面。枫属大多组都只具有3沟而不具内孔,为3沟型,3孔沟只出现于鸡爪枫组和大花枫组,并且枫属物种的花粉外壁条纹具有较丰富的多样性。
3.2 孢粉学特征在枫属分类中的应用
枫属组间甚至组下类群的花粉形态具有异质性,其中花粉大小、形状、萌发沟长等特征最明显,如鸡爪枫组,花粉粒极轴最长的三峡枫(P=33.66 μm)为最短的光叶枫(P=20.16 μm)的1.67倍,最大体积(羽扇枫)为最小体积(光叶枫)的1.45倍,并且存在近球形、近长球形、长球形及超长球形等多种花粉形状。相比而言,花粉孔沟形态和外壁纹饰在枫属组(系)内具有遗传稳定性。因此,孔沟形态和外壁纹饰可作为枫属组(系)分类的依据,而花粉大小、形状等特征则可用于种级分类。
橄榄枫和秀丽枫性状相似,分布区域高度重叠,前人基于翅果角度、柱头形态等特征将两者处理为不同物种[10,12]。2008年,新版《Flora of China》将橄榄枫修订为秀丽枫。本研究发现两者的外壁纹饰和萌发孔类型相似,仅在花粉大小、萌发沟长等特征方面存在微小差异。结合分子证据[17],两者在5条DNA条形码序列上都高度相似,表明两者关系极近。但是,在杭州植物园实地观察表明橄榄叶片略小,柱头反卷,并且花期比秀丽枫晚14 d左右。鉴于此,建议将橄榄枫处理为秀丽枫的一个亚种。
3.3 基于孢粉特征推测枫属的系统发育
有研究认为现代被子植物花粉的一般演化规律为3沟到3孔沟[26]。本研究中,观察到枫属3个组为3孔沟类型。化石信息[13]和分子证据[15-16]证明金钱枫比枫属更为原始,并且无患子科其它属大多表现为3孔沟形态[29]。因此,认为枫属类群中花粉萌发孔(3孔沟)的消失是进化的表现。枫属类群既有3沟型也有3孔沟型的花粉,表明该属存在处于不同进化水平的2种类型,其中鸡爪枫组和大花枫组具3孔沟,属于枫属较原始类群。此外,花粉形态特征在推测鸡爪枫组的起源和演化上也具有重要参考意义。鸡爪枫组为枫属中物种数量最丰富的一组,同时也是唯一一个在北美第三纪地层中未发现化石记录的现存组[13]。从鸡爪枫组花粉具内孔可以推断该组较原始,但该组最早的化石记录直到早中新世才在欧亚中纬度地区出现[30],因此推测该组可能由早期扩散至亚欧大陆的枫属原始类群(如古穗花枫组)演化而来。同时,鸡爪枫组在花粉大小、形状等特征上表现出高度的种间差异性,结合该组物种分布区域(喜马拉雅地区-我国西南)的气候和生境异质条件进行分析,推测该组在历史上经历了辐射进化,导致该组物种“爆发式”发生并具有丰富的种间多样性。
猜你喜欢
植物研究(2021年5期)2021-03-02
音乐教育与创作(2019年12期)2019-04-11
时代英语·高一(2019年1期)2019-03-13
小学生导刊(2018年20期)2018-11-30
小溪流(画刊)(2018年7期)2018-11-28
散文诗(2017年18期)2018-01-31
农业与技术(2017年15期)2017-09-13
中学生阅读(初中版)(2016年11期)2017-01-13
海峡姐妹(2016年9期)2016-02-27
东北林业大学学报(2015年2期)2015-03-10
