
濒危植物东兴金花茶种子萌发及幼苗光合特性研究
2022-10-10 11:07秦惠珍柴胜丰唐健民
种子 2022年8期
秦惠珍,邹 蓉,柴胜丰,唐健民,韦 霄
(广西壮族自治区中国科学院广西植物研究所/广西喀斯特植物保育与恢复生态学重点实验室,广西 桂林 541006)
东兴金花茶(Camelliaindochinensisvar. tunghinensis)为山茶科山茶属常绿灌木[1],是我国极小种群野生植物,分布范围狭窄,现存面积仅52 hm2[2]。东兴金花茶枝条浓密、花繁叶茂、树型优美,是培育茶花新品种的优良种质资源。此外,东兴金花茶富含黄酮、多糖等有效成分[3-7],具有清热解毒、利尿消肿、止痢、收敛止血等功效[8-9]。由于东兴金花茶生境脆弱,分布范围小,种子自然更新困难,乱砍滥伐现象严重,已经列入国家二级保护植物[10-11]。此外,因其分布范围狭窄,被列为“极小种群野生植物”。
目前,关于金花茶组植物种质收集和保存的研究较匮乏,对东兴金花茶的种子繁殖鲜见报道。本课题组前期研究发现,东兴金花茶扦插成活率低且金花茶植物组培技术尚未得以攻破,推测有性生殖是其野外和引种栽培的主要繁殖途径。在东兴金花茶的原生境中,普遍存在花多果少的现象,结籽率非常低,且果实极易被动物采食,种子资源十分有限。东兴金花茶作为茶族“皇后”中的一种,在园林园艺和科研中具有重要价值,但因其基本属于野生状态,缺乏足够的苗木资源,使许多研究难以开展。种子萌发是植物天然种群更新的基础[12],种子萌发和幼苗的生长影响着植物种群的发展、生存和繁衍[13]。萌发阶段是种群更新最薄弱的环节[14],是种群生活亏损最严重的时期之一[15],幼苗的自然更新不足以补充衰老个体的数量,种群就会逐渐走向灭亡。因此,了解东兴金花茶自然种群种子萌发率和幼苗的生长特性可为揭示其濒危机制、制定保护策略提供理论依据。
1 材料与方法
1.1 材 料
东兴金花茶种子于2020年12月采集于广西防城金花茶自然保护区(108°06′41″E,21°44′48″N)。采集后及时去除果皮和杂质,挑选饱满、无病虫害的种子用于观察和测量,测量完成后于湿沙中保存。
1.2 种子形态特征观测及种子吸水率测定
观察并记录东兴金花茶种子的性状及表型特征,随机选取20粒种子,用游标卡尺测量种子的长和宽;随机选取100粒种子测其重量,设5次重复,用百粒重估算千粒重,以平均值±标准差表示;随机选取50粒种子,采用TTC染色法测定种子生活力,设3次重复;设计13个浸种时间段(0、2、4、6、8、10、12、14、16、18、20、22、24 h)测定种子吸水率,每处理20粒种子,重复3次,将种子浸泡在25 ℃恒温水浴中。
种子吸水率(%)=[(种子吸水后重量-吸水前重量)/种子吸水前重量]×100%。
1.3 种子萌发特性观测
选取无病虫害的种子,于2021年1月初播种。在遮阴度为50%的遮阴棚播种,播种前种子用0.1%的KMnO4溶液消毒30 min,清水洗净。播种容器为36 cm×28 cm的黑色营养袋,采用沙土∶黄土=3∶1(体积比)基质进行播种,每处理设置3个重复,每个重复播种30粒。以芽顶出沙层为种子萌发标准,观察到第1粒种子萌发后,每7 d统计一次萌发数,试验时长120 d。进行常规浇水,保持基质湿润。
1.3.1种皮对种子萌发的影响
以完整种皮(ck)、种皮开口、去掉少部分种皮(少破)和完全去掉种皮(全破)的种子作为试验材料。
1.3.2种子大小对萌发的影响
分别选取不同大小规格的种子进行试验,大、中、小3个处理,每个处理3个重复,每个重复播种30粒种子,大种子的平均横径、纵径、厚度和单粒重分别为3.14 cm、2.87 cm、2.78 cm和11.24 g;中等种子的平均横径、纵径、厚度和单粒重分别为2.65 cm、2.43 cm、2.31 cm和9.26 g;小种子平均横径、纵径、厚度和单粒重分别为2.54 cm、2.11 cm、1.89 cm和5.45 g。
1.3.3不同浸种时间对种子萌发的影响
分别将种子置于25 ℃恒温水浴中,浸泡0、2、4、6、8、10 h后进行播种。
1.4 萌发指标计算
选用发芽势(Germination Energy,GE)、萌发率(Germination Percentage,GP)、发芽指数(Germination Index,GI)、活力指数(Vigor Index,VI)等4个指标评价种子的活力,本试验限定为30 d。
GE=(规定时间种子发芽数/播种种子数)×100%;
GP(%)=(萌发种子数/播种种子数)×100%;
GI=∑(Gt/Dt),Dt为发芽时间,Gt为对应发芽时间的发芽粒数;
VI=地上部分鲜重×发芽指数。
1.5 幼苗生长参数测定
萌发试验结束后,在不同大小、不同种群、不同冠层、不同种皮处理和不同浸种时间的5个处理中,分别随机取10株早期萌发的幼苗,剪掉留存的种子,用清水洗净擦干,用电子天平、游标卡尺、直尺测量并记录株高、根长、基径、地上部分鲜重、地上和地下部分生物量,其中生物量为在105 ℃烘箱中烘0.5 h后,再在65 ℃下烘至恒重。
1.6 幼苗光合特性测定
1.6.1光合-光响应曲线
种子萌发试验结束后将幼苗单株移栽至20 cm×25 cm营养袋中,进行常规水肥管理。6月晴天09:00—11:30时测定光合-光响应曲线,测定时选取生长健壮的幼苗,相同叶位且完全展开的成熟叶片进行测量,每个株系进行3个生物学重复。测定项目包括气孔导度(Stomatal conductance,GS)、净光合速率(Net Photosythetic Rate,PN)、蒸腾速率(Transpiration Rate, TR)和胞间二氧化碳浓度(Intercellular carbon dioxide concentration, CI)。测定时光合有效辐射(Photosynthetically Active Radiation, PAR)设置为1 200、1 000、800、600、400、200、150、100、80、60、40、20、0 μmol/(m2·s),流量设置为500 μmol/s。光源由便携式光合作用测定系统Li-6400(USA)自带的红蓝光提供。
1.6.2光合-CO2响应曲线
测定时间和叶片选择同光响应曲线,测定时设置叶室温度为25 ℃,相对湿度为60%,根据光合作用光饱和点测定的结果,光源控制光强在600 μmol/(m2·s)以内[当光强大于600 μmol/(m2·s)时,植株会出现光抑制现象],光合测定仪的流速设定为500 μmol/(m2·s),通过安装高压浓缩CO2小钢瓶,设定CO2浓度梯度为0、50、100、150、200、300、400、500、600、800、1 000、1 200、1 600 μmol/mol。每个CO2浓度下控制测定时间为120 s,数据稳定后仪器自动记录叶片净光合速率、蒸腾速率、气孔导度、胞间CO2浓度。
根据得到的不同光合有效辐射和CO2浓度下的PN,在弱光状态(0~100 μmol/mol拟合得到表观量子效率Apparent quantum yield,AQY),采用叶子飘[16]光合计算软件分别拟合绘制光合速率的光合-光响应曲线、光合-CO2响应曲线,并得到最大净光合速率(Maximum net photosynthetic rate,Pmax)、光饱和点(Light saturation point,LCP)、光补偿点(Light compensation point,LSP)、CO2饱和点(CO2saturation point,CCP)、CO2补偿点(CO2saturation point,LSP)、暗呼吸速率(Dark respiration,Rd)、光呼吸速率(Photorespiration rate,RP)。
1.7 数据分析
种子萌发数据、幼苗生长数据和光合作用参数采用Microsoft Excel 2010软件进行数据统计,采用SPSS 19.0统计软件(OneWay ANOVA, Duncan post hoc test,p<0.05)进行不同处理间的多重比较分析。统计值以平均值±标准误表示。采用Origin 8.5软件绘图。
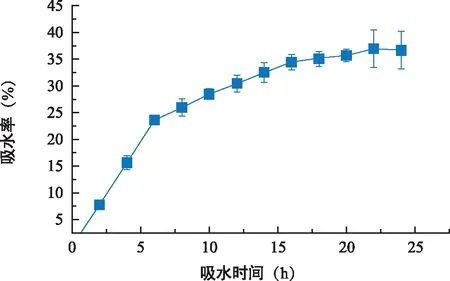
图1 东兴金花茶吸水特性曲线Fig.1 Water absorption property curve of C. indochinensis
2 结果与分析
2.1 种子形态特征和吸水特性
东兴金花茶种子近球形,横径(2.63±0.34)cm,纵径(2.45±0.28)cm,种皮黄棕色。种子单粒重(2.431±1.355)g,生活力(96.78±1.57)%,生活力较强。种子吸水特性见图1,随着浸种时间的延长,种子的吸水率提高,但随着时间变化,种子吸水率的增长速度发生变化前6 h,种子吸水率增长速度最快,6 h后增长速度减慢,在22 h之后基本不再增长。可见,东兴金花茶种子浸种早期吸水快,中期变慢,后期便达到吸水饱和。
2.2 种皮破损对种子萌发及幼苗早期生长的影响
种皮完整度对东兴金花茶种子萌发具有显著性影响(表1),种皮少破有助于加速种子萌发,发芽势、发芽率、发芽指数和活力指数显著高于对照和全破处理(p<0.05)。种皮少破时种子的发芽率达47.22%,是ck的1.60倍,而全破处理种子萌发率仅13.33%,为ck的45%。
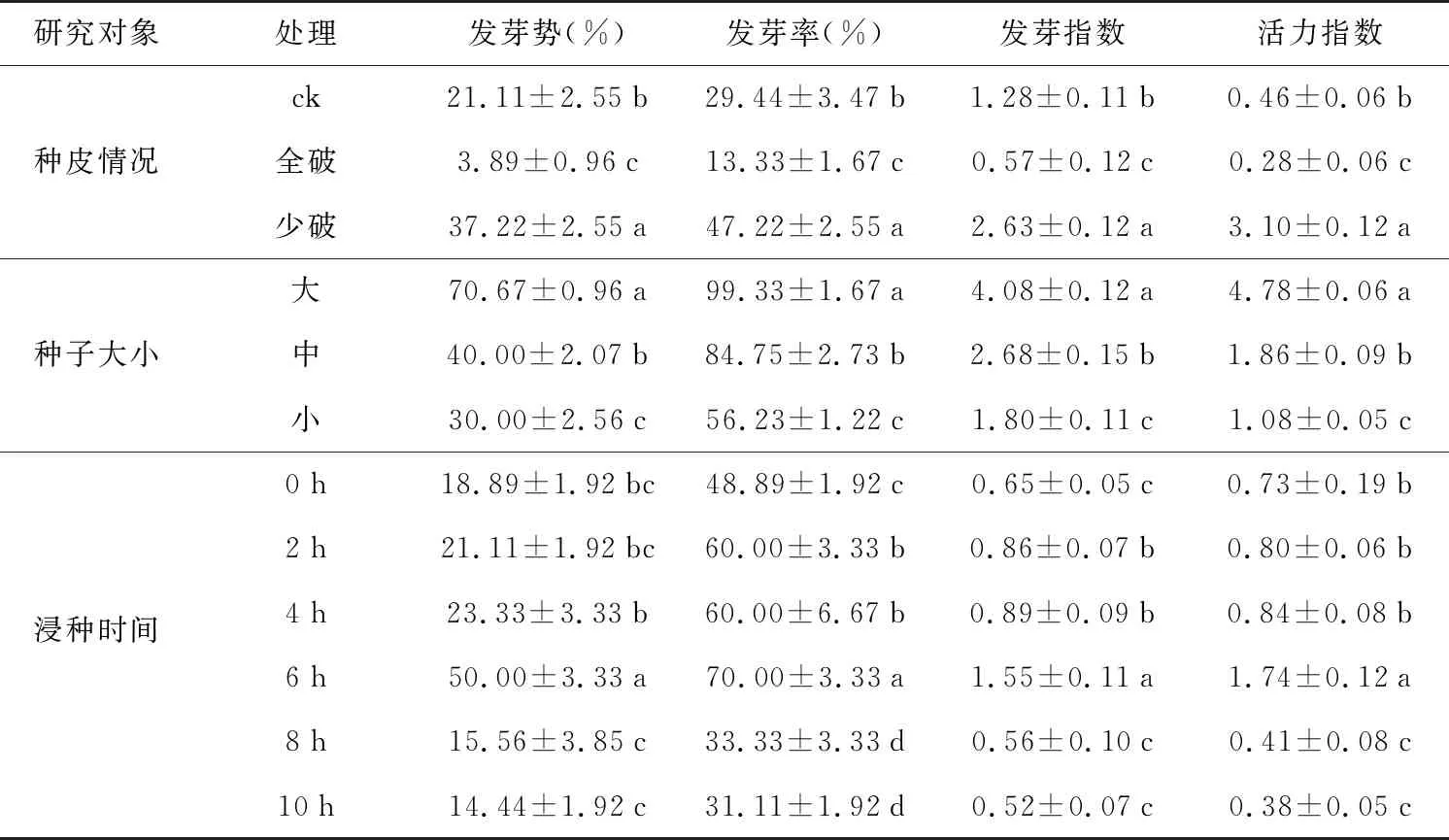
表1 不同处理条件下东兴金花茶的萌发参数Table 1 Germination parameters of C. tunghinensis seeds in different treatments
如图2所示,种皮少破处理幼苗的株高显著高于对照和全破处理,且幼苗的地上部分和地下部分生物量显著大于其他两个处理。因此,对东兴金花茶的种皮进行少破处理能显著加快其萌发进程和生长速度,但将种皮完全去除时会严重影响其种子萌发。
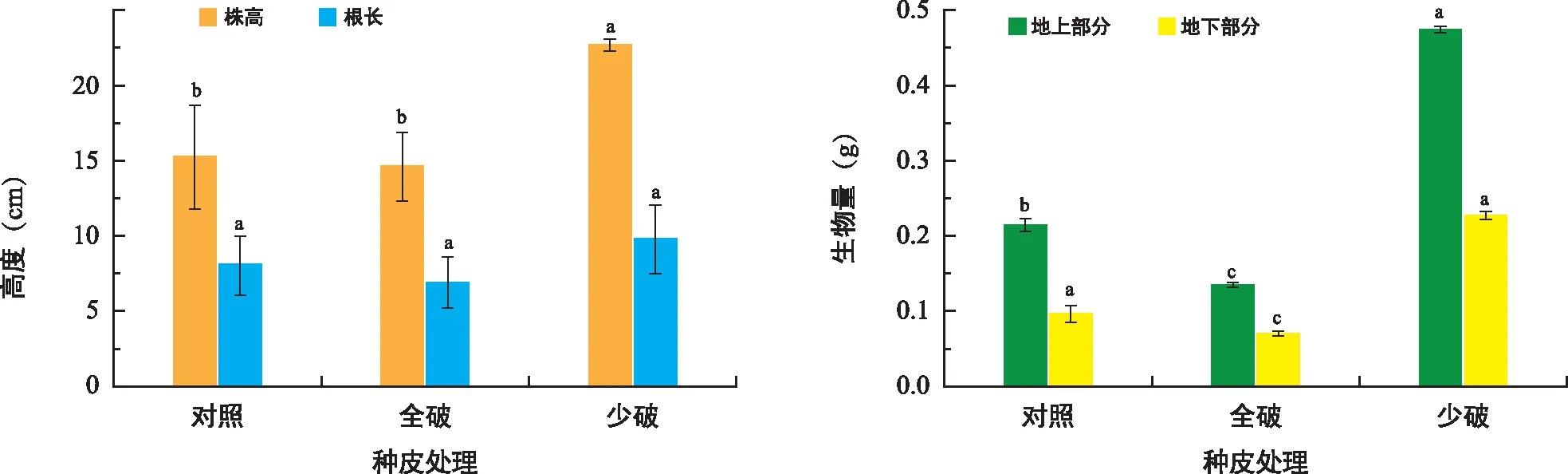
图2 种皮对东兴金花茶生长的影响Fig.2 The effect of seed coat on C. indochinensis growth
2.3 种子大小对种子萌发及幼苗早期生长的影响
在种子萌发的过程中,大小不同的种子萌发参数具有显著性差异,大种子的发芽势、萌发率、发芽指数和活力指数显著高于中种子和小种子(表1),其中大种子的发芽率接近100%,小种子的发芽率只有56.23%。
如图3所示,对于幼苗的生长,无论是株高、地上部分的生物量还是地下部分的生物量,大种子均显著大于中种子和小种子,而对于根长和地下部分的生物量而言,则具相反趋势,小种子的根长显著大于大种子和中种子,但地下部分的生物量与大种子并没有显著性差异,可见,小种子幼苗的生长更多养分集中在根等地下部分。
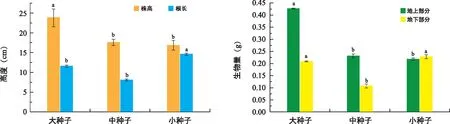
图3 种子大小对东兴金花茶生长的影响Fig.3 The effect of seed size on C. indochinensis growth
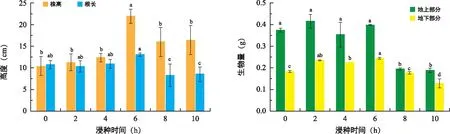
图5 浸种时间对东兴金花茶生长的影响Fig.5 The effect of soaking time on C. indochinensis growth
2.4 不同浸种时间对种子萌发及幼苗早期生长的影响
东兴金花茶的发芽势和发芽率因浸种时长不同而不尽相同,如图4所示,东兴金花茶的发芽势和发芽率随浸种时间的增加呈相似的趋势,均在浸种6 h时达最大值(发芽势和发芽率分别为50.00%和70.00%),显著高于其他浸种时长,超过6 h后东兴金花茶的发芽势和萌发率急剧降低(表1)。
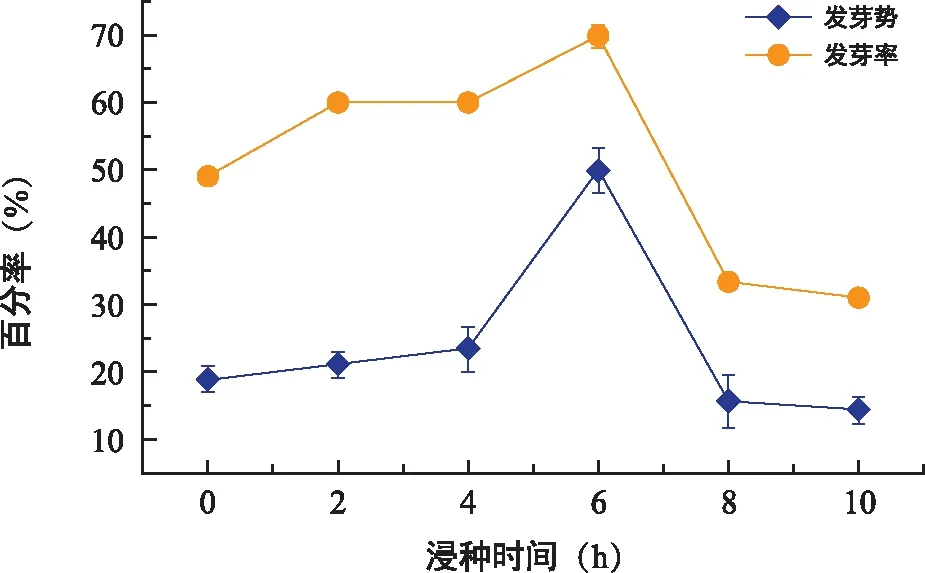
图4 不同浸种时间对东兴金花茶发芽势和萌发率的影响Fig.4 The effect of different soaking time on seed germination potential and germination rate of C. indochinensis
如图5所示,浸种时间为6 h的幼苗株高(21.97 cm)显著高于其他浸种时间(p<0.05),其根长与浸种时间为0、2、4 h的幼苗无显著性差异(p>0.05),与浸种时间为8、10 h具有显著性差异。浸种0、2、4、6 h幼苗的地上生物量显著高于浸种8、10 h幼苗的地上生物量,且浸种时间为6 h的地下生物量显著高于其他处理。可见东兴金花茶最佳的浸种时间为6 h。
2.5 幼苗光合生理特性
2.5.1光响应曲线
光合-光响应曲线反映了植物不同光照条件进行光合作用的能力。参照秦惠珍等[17]对东兴金花茶和长尾毛蕊茶成熟植株光响应曲线模型的研究,采用直角双曲线修正模型对东兴金花茶幼苗叶片的光响应曲线进行拟合,拟合程度较好(R2=0.99),从图6可以看出,在光照强度为0~250 μmol/(m2·s)时,净光合速率随光照强度的增强呈现直线上升的趋势;在250~600 μmol/(m2·s)时,随光照强度的增加净光合速率增长速度减慢,然后趋于稳定;光照强度大于600 μmol/(m2·s)时,净光合速率有下降的趋势,出现了光合抑制现象。根据叶子飘直角双曲线模型进行拟合得到东兴金花茶幼苗的Pmax、LSP、LCP、Rd、AQY等相关生理参数,东兴金花茶的最大净光合速率(Pmax)仅为2.67 μmol/(m2·s),光饱和点(LSP)、光补偿点(LCP)分别为484.25、2.68 μmol/(m2·s),可见东兴金花茶具有较低的光饱和点和光补偿点,且其表观量子效率为0.024,低于一般植物的实测表观量子效率(AQY=0.03~0.05),可见东兴金花茶的幼苗具有明显阴生植物的光合特性。此外,东兴金花茶暗呼吸速率较低,为0.35 μmol/(m2·s),说明其幼苗暗呼吸速率较小,有机物代谢效率高,这可能与植株处于幼苗阶段,不断积累有机物等特性有关。
2.5.2光合-CO2响应曲线
CO2响应曲线反映了植物利用CO2的能力。由图7可知,东兴金花茶幼苗的净光合速率随着CO2浓度的增大呈上升趋势,在所测定的范围内(50~1 600 μmol/mol )并没有呈下降或趋于稳定的趋势,表明东兴金花茶幼苗能利用较高浓度CO2进行光合作用。根据叶子飘直角双曲线模型对东兴金花茶光合-CO2曲线进行拟合,得到东兴金花茶幼苗的Amax、CSP、CCP及RP等相关生理参数,东兴金花茶的最大净光合速率(Amax)为12.24 μmol/(m2·s),CO2饱和点(CSP)、CO2补偿点(CCP)分别为2 317.00、3.83 μmol/(m2·s),可见东兴金花茶在CO2充足的情况下具有较高的净光合速率,其幼苗具有较高的CO2饱和点和较低的CO2补偿点,幼苗对CO2的利用范围较广。此外,东兴金花茶的光呼吸速率RP为0.04,光呼吸速率较小,表明其消耗有机物的能力较弱。
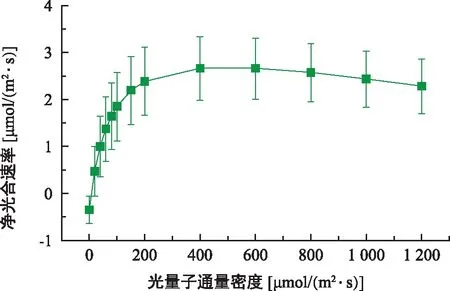
图6 东兴金花茶幼苗光合-光响应曲线Fig.6 Photosynthesis-light response curve of C. indochinensis seedlings
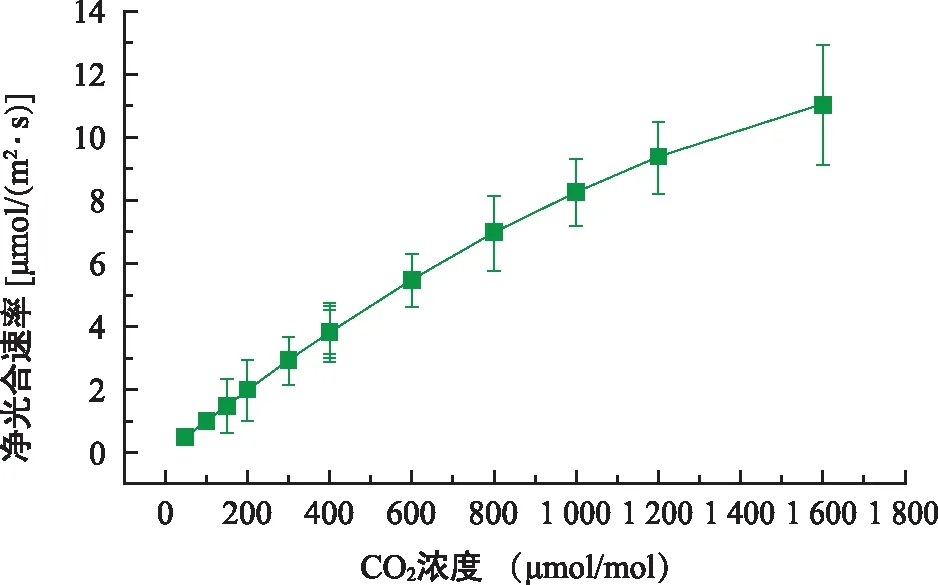
图7 东兴金花茶光合-CO2响应曲线Fig.7 Photosynthesis-CO2 response curve of C. indochinensis
3 讨 论
3.1 种皮对东兴金花茶种子萌发的影响
种皮是种子的保护屏障,同时也是种子萌发的阻碍,有些种子的种皮经过处理后能显著提高种子的萌发率和成活率[18-19]。东兴金花茶种皮蜡质,可能与其在不良环境下防止水分快速丢失和病原菌入侵的保护机制有关[20]。本研究结果表明,种皮对东兴金花茶种子萌发具有明显的束缚作用,去掉少部分种皮后,种子萌发时间缩短,发芽势和发芽率等各项指标显著提高,株高、根长、地上生物量和地下生物量显著增加,这与刘艳等[21]研究结论一致。东兴金花茶种皮经过开口处理后,发芽势显著提高,幼苗株高和根长等显著高于对照,说明其种皮对种子的萌发具有较强的抑制作用,去除少部分种皮后有利于水分和其他物质运输,从而促进了种子的萌发。另外,东兴金花茶完全去除种皮后,其发芽势、发芽率、株高和根长等显著降低,这可能是种皮破裂面积过大,使其种子极易受生物浸染腐烂,导致种子发芽率降低,这与赵东兴等[21]研究马槟榔种皮对种子萌发的影响结果相一致。此外,在采集东兴金花茶种子时发现,东兴金花茶果实成熟后果皮开始脱落,种皮易裂开,导致种胚多干燥失水或腐烂,这可能也是东兴金花茶自然繁殖率低的关键原因之一。
3.2 种子大小对东兴金花茶种子萌发的影响
种子大小决定种子内部储存营养物质的含量,一般而言,大种子具有更强的竞争优势和存活能力[23-24],而小种子在数量和传播方式等方面更具有优势[25]。有研究表明,种子大小与种子萌发率显著负相关[26],与幼苗成活率正相关[27]。也有研究表明,种子大小与萌发率之间没有直接的相关性[28]。本研究结果显示,东兴金花茶大种子的萌发率显著高于小种子,与张玲卫等[29]、朱雅娟等[30]研究得出的结果(草本植物中种子大小与萌发率呈现负相关)相反。一般来说,小种子在生态系统中更易形成土壤种子库,具有更高的萌发率和成活率。本研究结果并不支持此观点。东兴金花茶大种子比小种子具有更高的萌发率,原因可能是其胚乳能提供更多后期种子萌发过程需要的营养物质。
3.3 浸种时间对东兴金花茶种子萌发的影响
种子吸水量与浸种时间有关,但并不代表浸种时间越长对种子萌发越有利[31]。宋松泉等[32]指出,种子萌发过程中浸种处理有利于种子原生质膜的恢复和细胞内物质的转换,从而加速种子的萌发进程,促进幼苗生长[33]。本研究中不同浸种时间对东兴金花茶的发芽势和发芽率具有显著影响,浸种时间过长或过短都不利于种子萌发。研究表明,不同浸种时间对幼苗的生长影响不一致,但总的来说,浸种时间6 h,可以显著提高东兴金花茶种子的发芽势和发芽率,其幼苗的生长状况也最好。
3.4 光照强度和CO2浓度对东兴金花茶幼苗的影响
植物幼苗期是其生活史过程中最脆弱的阶段[34],特别是珍稀濒危物种幼苗的生存和生长发育关系到该物种的有效管理和保护[35]。光合作用是植物生长发育的基础,光照和CO2是影响植物光合作用的主要环境因子,对光照强度的适应幅度反映了其对生态环境的适应能力[36],研究濒危植物幼苗的光合特性是指导人工栽培的关键[37]。本研究结果表明,东兴金花茶具有较低的光饱和点及光补偿点,对光照的适应幅度较窄,且强光(光照照度大于600 μmol/(m2·s)出现了光抑制现象;而CO2饱和点较高,CO2补偿点较低,对CO2的利用能力较强。整体上看,强光对东兴金花茶幼苗的影响较大,在幼苗生长阶段应提供遮阴环境,避免阳光直射;同时可施用碳肥等促进幼苗进行光合作用,增加有机物的积累。
3.5 东兴金花茶种子和幼苗阶段的生态适应及致濒因素
种子形态特征和萌发方式是植物另一个关键的繁殖策略,关系幼苗的存活能力及个体未来的适合度[38]。尤其是濒危植物,种子阶段任何不利于种子萌发的因素都会对种群新个体的产生与补充产生直接影响[39],可能会加剧物种的濒危或灭亡。东兴金花茶种子平均单粒重为2.431 g,远高于乔木树种种子的平均质量0.328 g[40],大粒种子营养贮备更多,有利于萌发的幼苗在山林较厚的凋落物中成功建植[41]。东兴金花茶采取产生少量的大种子以便取得竞争优势的生殖策略,即K-对策[42],种子产出的周期长、成本高[43],种子成熟脱落后散落范围小,传播能力有限,种子难以扩张,东兴金花茶进入生殖阶段所需年限长,结实植株少,以至于林下幼苗数量少,加上前期的金花茶热潮,东兴金花茶大量成熟开花植株被砍伐,导致东兴金花茶种群难以恢复,使其濒危程度加剧。
东兴金花茶野外果实成熟期为12月,处于干旱少雨的季节,少部分种子在树上干枯死亡,部分种子成熟后脱落,随动物踩踏、雨水、凋落物分解等因素埋入土壤中作为潜在种群,翌年春天温度升高,雨水增多时萌发为种群补充后代。东兴金花茶的萌发特性是其自然种群更新的障碍所在,较长的果实成熟期和种子休眠期不利于种群空间资源的迅速占据,东兴金花茶从开花到果实成熟接近10个月,在这期间果实和种子要忍耐细菌、病虫害和动物捡食等各种环境压力。由实验可知,东兴金花茶的种子萌发需要一定的水分条件,水分过少或过多均不利于种子萌发。东兴金花茶果实成熟期在干旱少雨的季节,掉落时土壤水分少,种子难以萌发;翌年春季若雨量过多则导致土壤含水量高,种子易发霉变质,降低种子的萌发率;此外,东兴金花茶幼苗对强光特别敏感,强光下其光合作用受到抑制,光照适应范围狭窄,这也是导致东兴金花茶自然种群幼苗少、天然更新困难的重要因素。即种子萌发时对土壤水分的要求和幼苗对光照的要求难以适应多变的环境条件可能是其稀有的重要原因。因此,需加强东兴金花茶原生境的生态保护,注意保护其上层植被,为东兴金花茶提供一定的荫蔽度。此外,东兴金花茶幼苗的生长缓慢,植株矮小,与竞争能力强的植物竞争难以取得优势,也是其濒危的另一重要原因。种子萌发和幼苗阶段的生长受阻只是东兴金花茶致濒的一方面原因,应结合东兴金花茶的传粉生物学和分子遗传学等进行研究,从东兴金花茶的繁殖和遗传多样性水平了解其濒危机制。
4 结 论
东兴金花茶种子存在休眠期,萌发时间较长,蜡质种皮延长了种子萌发的时间,去掉少部分种皮可以促进其种子的萌发和幼苗的生长。东兴金花茶大粒种子贮存营养多,种子萌发和幼苗生长快。东兴金花茶种子萌发需要一定的水分条件,浸种可以加速种子的萌发。总体上看,东兴金花茶种子萌发过程长,幼苗生长慢,对光照要求严格,不利于种群空间资源的迅速占据,这可能是东兴金花茶种群衰落,形成极小种群濒危植物的重要原因。人工培育的东兴金花茶应及时采摘成熟果实,避免果实干枯失去活性,采摘后及时去掉外果皮留种皮,采用河沙等透气性较好的基质进行播种或沙藏。
猜你喜欢
文史春秋(2019年9期)2019-10-23
文史春秋(2019年9期)2019-10-23
文史春秋(2019年9期)2019-10-23
文史春秋(2019年9期)2019-10-23
学生导报·东方少年(2019年3期)2019-05-14
现代农业研究(2017年3期)2017-04-17
江苏农业科学(2016年4期)2016-06-14
江苏农业科学(2015年11期)2016-01-27
成长·读写月刊(2014年12期)2015-05-07
吉林农业(2014年5期)2014-07-09
