
山白树潜在适生区对气候变化的响应
2022-10-08 10:46杨黔越叶兴状徐毅枫覃名扬黄秋良张国防
福建农林大学学报(自然科学版) 2022年5期
杨黔越, 叶兴状, 徐毅枫, 覃名扬, 刘 宝, 黄秋良, 张国防
(福建农林大学林学院,福建 福州 350002)
气候是决定生物群落分布的主要因素,其变化不仅会改变物种的适应性,而且会对自然界的生态系统功能造成很大干扰[1].然而,近年来人类在工业生产中排放了大量的温室气体,使得自然生态系统功能受到严重影响[2],如群落水平稳定性[3]和植被生产力[4]等.同时,也造成了生物多样性下降以及植物空间分布格局的改变[5],引起生物带和生物群落空间分布的巨大变化.此外,在适应气候变化时,许多孑遗植物可能无法做出及时的转移而面临灭绝[6].因此,研究未来气候变化对孑遗植物分布产生的影响并制定出科学的保护策略,对物种多样性保护具有重要意义.
基于数据共享化以及空间分析技术,最大熵(maxium entropy, MaxEnt)模型成为目前使用最为广泛的生态位模型[7-8],在物种地理分布数据和环境因子数据的基础上实现对物种在该区域生存的预测,并对未来气候变化下植物潜在分布区进行预测,在入侵生态学[9]和保护生物学[10]中有广泛的应用,特别是在预测物种的现实分布和潜在分布的研究中,表现出较高的预测准确率[11].
山白树(Sinowilsoniahenryi)是金缕梅科(Hamamelidaceae)山白树属(SinowilsoniaHemsl.)的唯一树种,常为落叶灌木或小乔木[12],其被列入《中国高等植物受威胁物种名录》[13].山白树结构细密且材质较硬,是北方不可多得的用材树种[14];其根系具有较强的固土保水能力,也是河岸防护林常用的树种之一[15],还可防止水土流失.此外,山白树的花粉和种子含有丰富的脂肪和蛋白质,既可制成药品,又可开发出营养价值丰富的食品[16-17].山白树是我国特有的孑遗植物,然而其生境破碎化日趋严重,加之该树种为单性花,不仅结籽少,而且种子也缺乏媒介传播[18],使得山白树成为濒危植物.目前山白树研究主要聚焦于地理谱系[19]、生殖生物学[20-21]、遗传多样性[22]和系统发育[23]等.虽然周天华等[24]利用MaxEnt对其适生区分布变化进行了研究,但未对未来气候变化下山白树的空间迁移格局以及制约山白树地理分布的具体环境因子进行研究.近年来许多学者如朱耿平等[25]发现,调整了参数后的MaxEnt模型比默认参数的MaxEnt模型有着更好的预测结果,其ROC响应曲线更加平滑且更能准确反映出环境因子对物种分布的影响.2020年公布的第六次国际耦合模式比较计划(CMIP6)的共享经济路径(shared socioeconomic pathways, SSPs)气候数据的模拟效果较CMIP5的通用气候系统模式(CCSM4)[26]好.因此,利用最新的CMIP6气候数据及优化的MaxEnt模型对山白树适生区进行预测具有重要意义.
本研究采用Enmeval数据包优化后的MaxEnt模型对山白树当前和未来的适宜生境进行模拟,并分析影响山白树地理分布的气候因子以及在气候变化下山白树的空间迁移格局,以期为我国山白树野生资源的保护和利用研究提供依据.
1 材料与方法
1.1 数据来源及处理
通过检索中国数字植物标本馆、全球生物多样性信息平台和40个山白树的地理分布数据[27],并通过实地调查获得了19个山白树的分布点.ArcGIS 10.4中需要的地图数据来自国家基础地理信息系统中的中国行政区划矢量图(1∶400万).当代、2050s和2090s的环境数据是在世界气候数据库CMIP6模式下,通过共享经济路径提取的19个生物气候因子(表1).首先对收集的数据进行整理,即删除掉分布信息不全、分布重复记录以及人工栽培的分布点;然后将分布记录进行地理校正,并运用经纬度查询进行地理坐标转化,共得到45条山白树经纬度分布的点数据.此外,为降低群集效应所产生的误差[28],将经纬度分布的点数据导入ArcGis中,以10 km范围内的多个样本只保留一个点为原则进行缓冲区分析,最终确定了41个山白树的有效分布记录点,地理空间分布见图1.

图1 山白树的41个分布点

表1 影响山白树适生区分布的9个主导环境变量
1.2 优化后MaxEent模型的有效性评价
利用ENMeval语言包对模型进行优化,并将MaxEnt模型提供的6种函数即阈值函数(T)、片段函数(H)、交互函数(P)、线性函数(L)、二次项函数(Q)进行组合[25,29],得到的11个特征组合分别为L、H、HP、PT、QT、QH、QHP、LPT、LQH、LQPT以及LQHPT.再对参数进行测试,最终选取最佳的参数组合进行MaxEnt模型建模[30-31].在MaxEent3.4.1软件中分别导入山白树的地理分布数据(csv格式)和19个环境变量(asc格式).选取75%的分布数据建立模型,25%的分布数据用于检验模型,将预测结果设置为logistic,输出格式为asc格式,其余参数设置为默认值,并重复10次;再通过刀切法(Jackknife)计算变量的贡献率和置换重要性.采用受试者工作特征曲线(ROC)来评价模型的预测精度,ROC的AUC值越大说明预测的准确性越好.其取值为0~1,AUC值为0.9~1.0,表示预测结果优秀[32-33].
为了进一步分析山白树的生态适宜性,将MaxEent模型模拟出的结果导入ArcGIS 10.4软件中进行可视化处理,再利用Manual的分级方法将山白树的适生区分为4个等级[34],分别为高度适生区(0.7~1.0)、中度适生区(0.5~0.7)、低度适生区(0.2~0.5)和非适生区(0~0.2),并在ArcGIS10.4软件上生成山白树的适宜性分布等级图.
1.3 山白树的潜在适宜区分布格局
将山白树适宜区的空间迁移格局分为保留区、丧失区和新增区,其中保留区表示当前和未来均存在的区域;丧失区表示在当前存在、在未来消失的区域;新增区表示在当前不存在、未来存在的区域.根据山白树适宜生境预测结果,将适宜区(分布存在概率值大于0.5)和非适宜区(分布存在概率值小于等于0.5)分别赋值为“1”和“0”,以此来区分山白树的地理分布是否存在;再进一步分析山白树在2050s和2090s的气候情境下适宜分布区的变化过程,并定义保留区(1→1)、丧失区(1→0)以及增加区(0→1)[35];最后将结果导入ArcGIS 10.4中,绘制出山白树的适生区迁移格局.
2 结果与分析
2.1 模型的准确性评价
基于山白树的41个分布数据,利用刀切法分析19个环境变量对山白树适宜分布区的贡献大小,得到9个贡献率较大的环境变量,结果见表1.将筛选出的9个环境变量导入优化后的MaxEnt模型建模,并对山白树的当代与未来潜在地理分布区进行模拟.最终从MaxEnt模型得到ROC曲线,在10次重复模拟后得到的平均AUC值均大于0.987.根据AUC的评价指标可知优化的MaxEnt模型具有很好的预测能力和稳定性.
2.2 制约山白树地理分布的主要环境因子
贡献率最大的6个环境因子与山白树存在的概率关系如图2所示.存在概率大于0.7说明环境适合山白树生长.
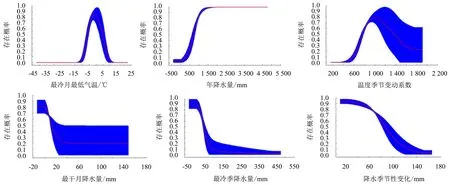
图2 山白树主导环境变量的响应曲线
2.3 不同时期山白树潜在适生区及其面积变化
MaxEnt模型预测结果表明,山白树当代适生区面积为37.87×104km2,约占国土面积的3.94%,其中,高、中、低度适生区面积分别为1.81×104、13.09×104和22.97×104km2,分别占国土面积的0.18%、1.37%和2.39%(表2).从图3可知,山白树的适宜生境以四川安县、重庆城口、湖北房县、河南焦作、甘肃天水、山西垣曲、陕西眉县为边界,形成了一个较为连续的适宜分布区.其中,高度适生区主要分布于秦岭一带,中度适生区分布于陕西南部,低度适生区分布于河南西部,并延伸至山西南部至甘肃东部及湖北北部至四川北部.

表2 山白树在不同时期的适生区面积
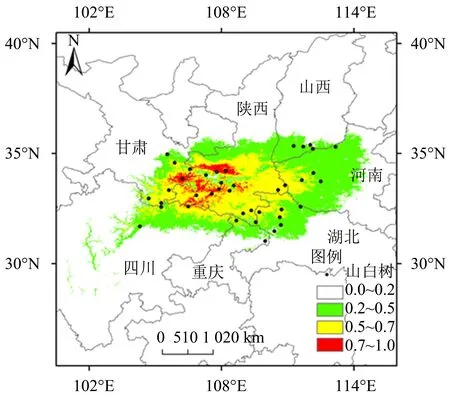
图3 山白树当前气候条件下的适生区分布
在未来气候变化条件下,山白树的适宜生境与当前相比向西移动的趋势如图4所示,并且在2050s时山白树的总适生面积均有减小的趋势,与当前相比分别减少2.58×104、0.91×104、4.17×104km2(表2);在2090s时,山白树的的适生面积在SSP126的气候情景下减少了0.52×104km2.但在气候情景为SSP245和SSP585的情况下其适生面积均有所增加,分别增加了0.61×104、0.60×104km2.

a:2050s—SSP126;b:2050s—SSP245;c:2050—SSP585;d:2090s—SSP126;e:2090s—SSP245;f:2090s—SSP585.
2.4 未来气候变化下山白树适宜生境的空间迁移格局
将山白树未来的空间迁移总变化与其当代的中度适生区和高度适生区的面积之和进行比较,表明在2050s时,SSP585排放情况下丧失面积最大,为2.79×104km2,丧失率达到了12.13%,面积的总变化减少了1.41×104km2(表3).其丧失面积主要分布在湖北的西北部、陕西南部以及四川东北部地区(图5c).在 2050s时,SSP245排放情况下增加面积最大,为2.72×104km2,增加率达到了11.85%,面积的总变化增加了1.11×104km2(表3).其增加面积主要分布在甘肃东南部、河南西部以及湖北西北部.在2090s年代的3种浓度排放情景下,山白树适生面积均呈增长趋势,即新增面积均大于丧失面积,在SSP126、SSP245、SSP585下面积的总变化分别增加了0.47×104、1.11×104、0.43×104km2(表3).说明在未来两个年代中2090s更适合山白树的生存和繁殖;同样,在3种气候情景下,SSP245的排放浓度可为山白树提供相对最适宜的生长条件.

a:2050s—SSP126;b:2050s—SSP245;c:2050—SSP585;d:2090s—SSP126;e:2090s—SSP245;f:2090s—SSP585.
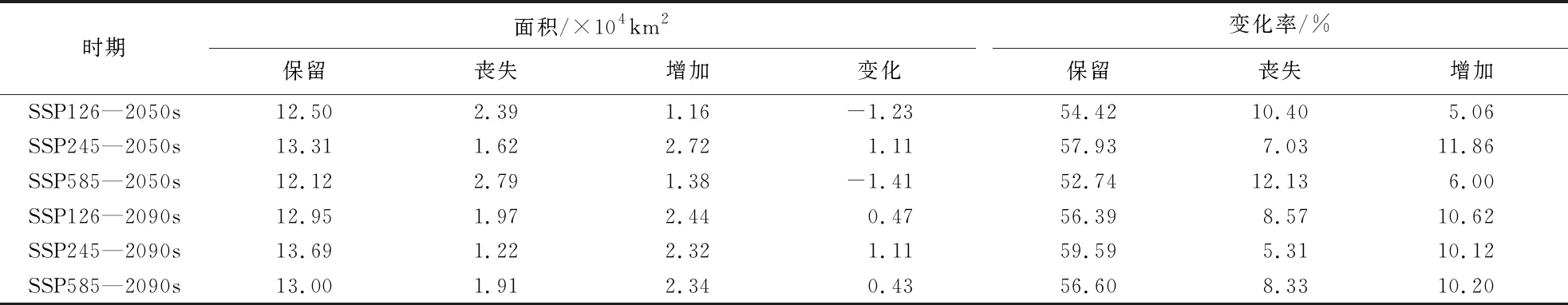
表3 山白树在不同时期的空间分布变化
3 讨论
本研究结果表明,山白树当代的适宜生长区主要分布在陕西南部、山西南部、河南中西部、湖北西北部以及重庆、四川东北部,甘肃东南部,与山白树在当代的主要分布范围重合度较高,同时也与周天华等[24,19]的预测结果相一致.说明本研究结果可靠,也表明山白树近20 a来的分布格局变迁不大.这可能是因为山白树对生存环境的要求比较严格,其生长环境对林地的郁闭度、相对空气温度、水分条件以及日照时间等的要求都较高,而秦岭、巴山以及中条山这一带正好满足其对生长环境的需求.此外还发现,山白树的高度适生区主要位于秦岭、九嵕山和大巴山[36].山白树的适宜生长区在2050s的3种气候情景下均有减少的趋势,并且其高度适生区在逐渐地向西迁移,这与周天华等[24]的模型预测结果不太一致.原因可能有以下几方面:其一、模型的构建不同.本研究采用了通过ENMeval数据包优化后的MaxEnt模型,即对MaxEnt模型的调控倍频和特征组合参数分别进行了调整,通过降低模型的复杂性提高模型的预测精度[27];而周天华采用的则是以默认参数来构建的模型.研究[32,37]发现,基于默认参数构建的MaxEnt模型与优化后的MaxEnt模型相比,其预测的结果可能存在着严重的拟合偏差[38].其二、本研究中当代数据采用30arc-second的空间分辨率,未来数据采用的是2.5arc-minutes的空间分辨率,均比周天华等[24]采用的精度要高.其三、气候因子的选择不同.本研究与周天华等[24]的研究分别有9个和7个气候因子参与建模,其中重复的气候因子只有2个,这可能是造成两研究预测结果有所不同的主要原因.
MaxEnt模型预测结果显示,影响山白树未来适生区分布的气候因子主要为温度和降水.且在2090s年代山白树的适生区范围有扩大的趋势.这与周天华等[24]利用最大熵模型对山白树适生区变化的研究结果一致.其中,影响山白树最适宜的最冷月最低气温为-4.08~4.12 ℃,当温度低于-4.08 ℃或高于4.12 ℃时,其生存概率的下降趋势明显增大.影响山白树最适宜的年降水量为815.48~2 749.32 mm,当年降水量低于815.48 mm时,其生存概率急剧下降,表现出山白树喜水的特性,这与岳志宗[39]对山白树分布与生境的研究结果一致.冬季的低温使山白树能够进行种子休眠,降水量的增加则为山白树来年的种子萌发提供了充足的水分条件,有利于其种子萌发[40].研究[41]表明,山白树常生于河边、干河床中或沟谷旁,能抵抗间歇性的短期水浸泡.此外,江志红利用IPCC-AR4模式预测在21世纪末期年降水量最高将提高11%[2].章杰[42]研究秦岭地区未来的气候变化趋势时指出,在未来温室效应加剧的情况下,可能导致降水频率和降水量增大.未来降水的增多是山白树在2090s时适生区范围扩大的重要原因之一.
水分条件和热量条件均为植被地理分布的重要影响因素,且两者之间通常为互补关系[43].结合山白树适宜的水热因子推测,最冷月最低气温和年降水量限制了山白树越过黄土高原和中温带南界分布;温度季节变化系数限制了山白树越过大巴山、武当山.这3个气候因子可能是限制山白树集中分布在我国暖温带地区及亚热带北界地区的重要原因之一.
未来3种气候情景下,2050s—SSP585情景时的极端条件下山白树的适生区面积最小,为33.69×104km2.与当代山白树的分布数据相比较,其丧失区面积为2.79×104km2,丧失率达到了12.13%,空间分布格局总体上也呈减少趋势.这可能是因为在2050s—SSP585的情景下,降水和温度增幅较大,当增幅超过山白树适宜的生长范围时,山白树的生境破碎化更严重,使得其适生区范围收缩幅度较大.山白树对生长环境的要求比较严格.张立军[44]研究发现气候变迁和人类过度干扰使得山白树生境片段化日趋严重,加剧其地理隔离,降低了山白树居群之间的基因交流,使得各居群间呈现“孤岛”化分布,这与本研究得出的高度适生区破碎化程度较高相一致,同时也可能是分化出变种秃山白树(S.henryivar.glabrescens)的主要原因之一.此外,山白树通常采用实生繁殖,傅志军等[41]利用湿沙层积法对其催芽后再培育,结果表明出苗率高达了95%以上.
4 结论
本研究结果表明,制约山白树适宜生境变化的气候因子主要有最冷月最低气温、年降水量、温度季节变动系数、最干月降水量;其适生区分布主要在鄂西和秦岭山地的中低山地区.在3种气候条件下,2050s时山白树的总适生区总体上呈减少趋势,平均减少2.56×104km2;而2090s时,其总适生区在总体上呈增加趋势,平均增加0.23×104km2.山白树在未来的保留区主要集中在秦岭、九嵕山和大巴山,这些区域内的自然保护区均可作为山白树在未来气候变化条件下的避难所和生长地.
猜你喜欢
中国中医药信息杂志(2022年7期)2022-07-17
中国野生植物资源(2022年3期)2022-04-19
云南农业大学学报(自然科学)(2022年2期)2022-04-07
福建农林大学学报(自然科学版)(2022年1期)2022-01-18
中国农村水利水电(2021年11期)2021-11-29
风景园林(2021年2期)2021-04-24
落叶果树(2021年6期)2021-02-12
南方农业·下旬(2016年9期)2017-04-27
江苏农业科学(2017年5期)2017-04-15
科技与创新(2016年18期)2016-11-04
