
连栽对杉木人工林土壤表层真菌群落结构及功能的影响
2022-10-08 10:50:18彭丽鸿崔朝伟王佳琪马东旭任正标何宗明林开敏
福建农林大学学报(自然科学版) 2022年5期
彭丽鸿, 崔朝伟, 王佳琪, 马东旭, 任正标, 何宗明, 林开敏
(1.福建农林大学林学院,福建 福州 350002;2.国家林业和草原局杉木工程技术研究中心,福建 福州 350002;3.大伊山旅游度假区管委会,江苏 连云港 222200)
杉木人工林具有速生丰产、适应性强、经济效益高等优点,在我国南方广泛种植.但伴随着杉木人工林纯林化、针叶化、集约化和多代连栽现象的日益严重,林分生物多样性降低、水土流失、地力衰退、生产力下降等生态问题严重影响其可持续发展[1-2].大量研究[2-5]表明:杉木连栽会造成土壤理化性质恶化或衰退,比如土壤容重增加、土壤通气以及容蓄能力下降、土壤养分含量下降以及酶活性降低等.土壤微生物作为土壤生物中最重要的组成部分之一,在植物生长、土壤肥力形成和发展、营养元素循环等方面发挥着重要作用[6-7].土壤微生物群落以土壤真菌为优势群落,土壤真菌是调节生态系统功能的重要驱动力[8],如菌根等真菌在提高植物生产力和抗病性以及缓解其他生物和非生物胁迫方面起到重要作用[9-10].腐生真菌占土壤真菌群落的50%以上,通常被认为是主导土壤中有机质降解和养分循环的主要分解者[11-12].以往的研究主要集中于连栽对人工林土壤理化性质的影响[13-15],连栽对于杉木人工林表层土壤微生物群落结构与功能类群影响的研究[16-17]较少,尤其是对理化性质和土壤真菌功能类群影响的研究鲜有报道.鉴于此,本研究以福建省南平市峡阳国有林场的杉木林为研究对象,研究杉木连栽对土壤真菌群落结构及功能的影响.利用ITS rRNA高通量测序技术揭示连栽对杉木人工林土壤真菌群落结构、功能及土壤环境因子的影响,旨在为杉木人工林地力衰退的防治提供参考.
1 材料与方法
1.1 研究区概况
样地设置在福建省南平市峡阳国有林场,属丘陵地带,年平均气温20 ℃,海拔350~700 m,坡度20°~30°;年均降水量1 500~1 600 mm,年均蒸发量1 300 mm左右,雨季3~6个月;年均相对湿度75.2%,无霜期290 d左右;土壤以红壤为主,土层中厚[18].林下植被主要有粗叶榕(Ficushirta)、毛冬青(Ilexpubescens)、毛柄短肠蕨(Allamtodiadilatata)、中华里白(Dicranopterischinensis)、观音座莲(Angiopterisfokiensis)、江南短肠蕨(Allantodiametteniana)、狗脊(Woodwardiajaponica)等.
1.2 试验设计
为了保证试验的准确性,尽量选取坡度、坡向、坡位、立地条件等较为一致的杉木人工林,2020年10月将峡阳国有林场中1967年实生苗造林形成的1代杉木人工林(CK,林龄53 a)和1997年对1代杉木人工林进行采伐后采用传统杉木栽培技术进行连续栽植的2代杉木林(TP,林龄23 a)设置为试验林.样地设计位置相邻,立地条件较好,林分地位指数均为18.
1.3 土壤采集方法
不同代数杉木林分的不同坡位(上坡、中坡、下坡)分别设置3个20 m×20 m的样地.在每一个样地内,按照梅花型挖取5个(0~10 cm)样地土壤剖面,均匀取土后进行等量充分混合;随机采集1 kg左右土样存放于自封袋中,带回实验室.土样分2批保存,部分放入-80 ℃冰箱保存,用于测定土壤真菌多样性、可溶性碳及硝态氮、铵态氮含量;另一部分进行烘干处理,过2 mm筛及0.149 mm筛,用于测定土壤化学性质等.
1.4 数据测定与分析
1.4.1 土壤全碳、全氮含量的测定 用碳氮分析仪测定过0.149 mm筛风干土的全碳、全氮含量.
1.4.2 全磷、全钾含量的测定 采用高氯酸—硝酸—氢氟酸消煮法,消煮液用电感耦合等离子发射光谱仪(PE OPTIMA 800)测定.
1.4.3 土壤可溶性碳的测定 采用硫酸钾浸提法测定过2 mm筛的新鲜土壤.
1.4.4 硝态氮、铵态氮的测定 采用氯化钾浸提—扩散法测定过2 mm筛的新鲜土壤.
1.4.5 微生物群落多样性分析 从新鲜土样中提取DNA后,利用带有barcode的特异引物扩增rDNA的保守区.然后将PCR扩增产物切胶回收,用QuantiFluorTM荧光计进行定量分析.将纯化的扩增产物进行等量混合,连接测序接头,构建测序文库,Hiseq2500 PE250上机测序.采用FLASH软件对原始数据进行质量过滤和双端序列的拼接,得到有效数据[9].采用Uparse软件,对相似度≥97%的序列归为1个操作分类单元,即OTU.运用Qiime软件对所得优质序列近物种进行分类分析,对OTUs进行丰度、Alpha多样性指数(Sob指数、Chao1指数、Ace指数、Shannon指数以及Simpson指数)等分析;采用FUNGuild数据库完成对真菌分类信息的注释[11];运用R语言进行土壤微生物NMDS分析以及相关性分析.使用Excel对数据进行整理以及图形绘制,采用SPSS 19.0软件对土壤养分和主要优势微生物的数据进行单因素方差(ANOVA)分析,采用独立样本T检验比较不同处理间的显著性(P<0.05),利用Canoco5.0软件做冗余分析[19].
2 结果与分析
2.1 杉木连栽对土壤养分的影响
对不同林分样地表层土壤进行理化性质分析,结果见表1.从表1可知:2代林中土壤养分含量整体呈下降趋势,而全钾含量有所上升,但并不显著;全碳、全氮、铵态氮含量与有机碳含量表现为2代林显著低于1代林(P<0.05);全磷含量无明显变化.说明2代连栽会降低土壤养分含量.

表1 不同代数杉木人工林土壤的理化性质1)
2.2 杉木连栽对土壤表层真菌多样性的影响
对所得优质序列按≥97%的相似度进行OTU聚类分析后得到了3 820个OTUs,数据有效率为91.03%~93.92%;CK和TP得到的有效序列分别为93.67%、92.57%,说明原始数据利用率较高,绝大多数都可用于后续数据分析.对不同林分样地表层土壤真菌进行Alpha多样性分析(表2),Alpha多样性指数表现为TP略高于CK,但独立样本T检验分析结果表明Alpha多样性分析指数在连栽前后差异不显著(P>0.05),说明TP可能会提高杉木人工林表层土壤真菌丰富度及多样性,但影响并不显著(P>0.05).

表2 Alpha多样性分析指数1)
2.3 连栽对土壤表层真菌群落相对丰度的影响
通过高通量测序得出,真菌群落在门和种水平物种数量上的相对丰度表现为TP
在属水平上真菌群落TP排名前十(从高到低)依次是木霉属(Trichoderma)、圆孢霉属(Staphylotrichum)、青霉菌属(Penicillium)、篮状菌属(Talaromyces)、Saitozyma、水曲霉属(Clonostachys)、拟青霉属(Purpureocillium)、大青属(Clerodendrum)、Leohumicola、弯颈霉属(Tolypocladium).主要优势群落属在TP与CK上表现为相同属,但相对丰度存在差异,主要优势群落木霉属、圆孢霉属、青霉菌属在TP及CK中相对丰度占比分别为12.14%与18.89%、6.52%与7.03%、4.75%与13.31%.经独立样本T检验,发现主要优势真菌群落的相对丰度差异性并不显著(图1b).
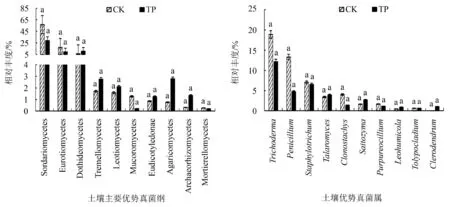
图1 土壤真菌群落在纲、属水平上的相对丰度
2.4 土壤表层真菌群落在纲、属水平上的多维尺度
为了解连栽对土壤真菌群落组成的差异性,分别对纲(图2a)、属(图2b)水平上土壤真菌群落进行非度量多维尺度(non-metric multidimensional, NMDS)分析.微生物群落组成的差异性是以组间距离来呈现的,差异越大距离越远,差异越小距离越近.由图2可知,不论在纲、属水平上,TP与CK间距离都相近,说明两者群落组成差异较小,即2代连栽对于表层土壤真菌的群落组成及结构的影响并不明显.

a.纲水平;b.属水平.
2.5 土壤真菌群落的功能
对TP及CK土壤真菌群落OTUs进行营养类型功能注解分析(图3),共检测出7种营养类型,分别是病原营养型(pathotroph)、病原—腐生营养型(pathotroph-saprotroph)、病原—腐生—共生营养型(pathotroph-saprotroph-symbiotroph)、病原—共生营养型(pathotroph-symbiotroph)、腐生营养型(saprotroph)、腐生—共生营养型(saprotroph-symbiotroph)和共生营养型(symbiotroph).相对丰度排前三的分别是腐生营养型、病原—共生营养型和病原—腐生营养型真菌;连栽前后腐生营养型真菌丰度无明显变化,除病原—腐生—共生营养型真菌丰度有所上升外,其他营养类型真菌丰度均出现一定程度的下降.独立样本T检验结果表明2代杉木林中腐生—共生营养型真菌的平均丰度极显著低于1代林(P<0.01).对其进行进一步的分类,发现该功能类群真菌主要优势类群是内生菌—凋落物腐生菌—土壤腐生菌—未定义的腐生菌和外生菌根—未定义腐生菌.这两类真菌类群的功能主要来源菌属为Mortierella属和Myxotrichaceae科.
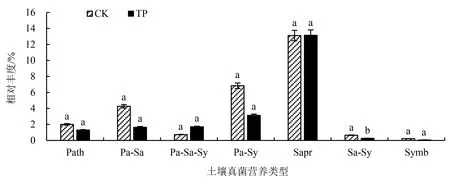
Path:病原营养型.Pa-Sa:病原—腐生营养型.Pa-Sa-Sy:病原—腐生—共生营养型.Pa-Sy:病原—共生营养型.Sapr:腐生营养型.Sa-Sy:腐生—共生营养型.Symb:共生营养型.
2.6 土壤真菌与理化性质的相关性
对Alpha多样性指数(图4)与土壤化学性质进行RDA分析,结果显示:第一、二主成分轴分别解释Alpha多样性的94%、5.5%.由图4可知,Sob指数与全碳、全氮、可溶性有机碳、硝态氮、铵态氮含量呈极显著负相关;Chao1和Ace指数与全碳、全氮、可溶性有机碳、硝态氮、铵态氮含量呈极显著负相关,与全磷含量呈显著负相关;Shannon和Simpson指数与全磷呈极显著负相关,与全碳、全氮、可溶性有机碳、硝态氮、铵态氮呈显著负相关,与全钾呈显著正相关.
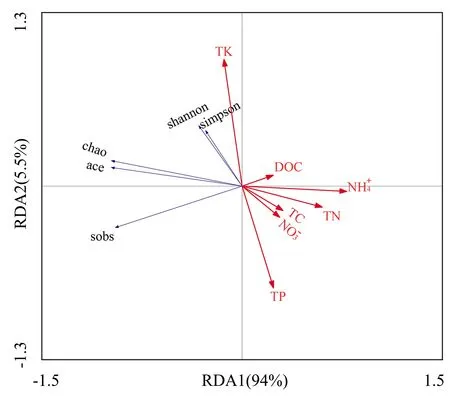
图4 Alpha多样性指数与土壤理化性质冗余分析
以土壤化学性质作为解释变量, 以不同纲、属水平上TP前6位的优势真菌群为响应变量进行冗余分析(图5).结果(图5)显示,第一、二主成分轴分别解释纲水平真菌群落组成差异的88.2%、11.3%(图5a);解释属水平上的土壤真菌群落组成差异的61.7%、19.6%(图5b).由图5a可知,粪壳菌纲与全碳、全氮、硝态氮、铵态氮含量呈显著正相关,与全钾含量呈显著负相关;散囊菌纲与全磷含量呈显著负相关,与有机碳含量呈正相关;座囊菌纲与全碳、全氮、铵态氮、硝态氮含量呈极显著负相关;银耳菌纲与全钾含量呈极显著正相关,与硝态氮含量呈极显著负相关;伞菌纲与土壤全碳、全氮、铵态氮含量呈极显著负相关;锤舌菌纲与有机碳含量呈极显著负相关.如图5b所示,木霉属与全碳、全氮、铵态氮、有机碳含量呈显著正相关;拟青霉属与全磷含量呈极显著负相关,与有机碳含量呈显著正相关;圆孢霉属与硝态氮呈显著负相关;篮状菌属与硝态氮呈极显著负相关;水曲霉属与硝态氮呈显著正相关;Saitozyma与全钾呈极显著正相关,与全碳、全氮、铵态氮呈显著负相关.

a.纲水平;b.属水平. Sordario:Sordariomycetes;Eurotiom:Eurotiomycetes;Dothideo:Dothideomycetes;Tremello:Tremellomycetes;Leotiomy:Leotiomycetes;Agaricom:Agaricomycetes. Trichode:Trichoderma;Penicill:Penicillium;Staphylo:Staphylotrichum;Talaromy:Talaromyces;Clonosta:Clonostachys;Saitosta:Saitozyma.
以土壤化学性质作为解释变量,以不同营养类型真菌群落为响应变量进行RDA分析(图6).结果(图6)显示,第一、二主成分轴分别解释不同营养类型真菌群落差异的71.5%、21.1%.其中病原营养型、共生营养型以及腐生—共生营养型真菌与有机碳含量呈极显著正相关关系,与全磷含量呈极显著负相关;病原—共生营养型真菌与全磷含量呈极显著负相关,与有机碳含量呈显著正相关;病原—腐生—共生营养型真菌与全碳、全氮、全钾、铵态氮含量呈极显著负相关,与有机碳含量呈显著负相关,与全磷、全钾含量呈正相关;腐生营养型真菌与全钾含量呈极显著正相关,与全磷、硝态氮含量呈显著负相关;病原—腐生营养型真菌与硝态氮呈显著正相关,与全磷、全钾含量呈显著负相关.
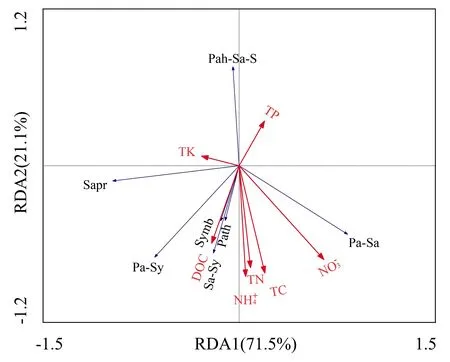
Sapr:腐生营养型.Path:病原营养型.Symb:共生营养型.Pa-Sa:病原—腐生营养型.Pa-Sy:病原—共生营养型.Sa-Sy:腐生—共生营养型.Pa-Sa-S:病原—腐生—共生营养型.
3 讨论与小结
本研究发现杉木人工林2代连栽会导致表层土壤全碳、全氮、铵态氮以及有机碳含量显著降低.连栽会导致土壤质量下降,对土壤养分含量及利用效率产生不同程度的影响.吕小燕等[20]和马祥庆等[13]认为土壤有机碳和全氮含量会随着连栽代数的增加而显著下降,这与本文研究结论一致.土壤微生物群落的丰富度及多样性对土壤状况的改变十分敏感,因此微生物群落变化可以作为反映土壤状况的指标之一[21].土壤微生物群落结构和组成在决定生态系统功能方面具有重要作用,尤其是有机质的转化和养分循环方面[22].本研究中,连栽对土壤表层真菌丰富度的影响与大多数研究结果[23-24]基本一致.2代杉木连栽可以在一定程度上提高表层土壤真菌丰富度,但影响程度并不显著.刘丽等[25]和王士亚[26]在对杉木人工林的研究中均发现3代连栽会造成土壤真菌多样性显著降低,而2代连栽对土壤真菌群落结构及多样性无显著影响,这与本研究结论一致.本研究结果表明连栽对表层土壤真菌alpha多样性指数无显著影响.前人在对杉木人工林的研究中发现,主要优势纲为散囊菌纲和粪壳菌纲[27].本研究发现杉木人工林1代林与2代林中的主要优势纲群落均为粪壳菌纲、散囊菌纲和座囊菌纲,主要优势属群落均为木霉属、圆孢霉属、青霉菌属.2代连栽对于优势菌群种类无显著影响,粪壳菌纲和散囊菌纲群落的相对丰度在连栽后有所下降,座囊菌纲群落丰度有所上升.粪壳菌纲真菌群落与散囊菌纲真菌群落均可以形成植物菌根,在帮助林木养分吸收方面具有重要作用[28-30],2代连栽在一定程度上可能会降低土壤养分的利用效率.独立样本T检验发现2代连栽对土壤真菌的组成及丰度影响并不显著,多维尺度分析表明2代连栽对纲、属水平上真菌群落组成与结构的影响并不显著.本研究结果表明2代连栽对表层土壤真菌在纲、属水平上优势菌群落组成、相对丰度以及结构的影响并不显著.
高尚坤[31]在马尾松不同营林措施土壤微生物群落特征影响的研究中发现腐生营养型真菌丰度在重度采伐及除灌后显著下降,而本研究发现2代连栽在一定程度上提高了腐生营养型真菌群落丰度,但影响并不显著.这可能与研究背景有关,高尚坤[31]的研究是在实施不同措施后第2年进行取样研究,除灌和采伐对土壤环境造成干扰,从而改变了腐生菌的营养来源.而本研究是2代林生长23 a后进行取样,采伐对土壤环境的干扰可能随着营林年限的增长而恢复;也可能因取样季节、林下植被、树种以及土地利用方式等不同而不一致[32-34].罗杨等[35]在对3代杉木连栽土壤根际镰刀菌属的研究中发现连栽会改变镰刀菌属的多样性,表现为先降低后升高的变化趋势,而本研究发现病原营养型真菌群落受2代连栽的影响并不显著,这可能与真菌功能类群划分差异有关.本研究发现,杉木人工林土壤2代连栽会导致腐生—共生营养型功能类群真菌群落的丰度极显著降低.腐生—共生营养型真菌的营养方式与土壤环境以及寄主植物的生长状态密切相关,通过寄主植物的健康状况和土壤环境选择不同的营养方式[36],连栽可能导致土壤养分发生改变,土壤环境的改变导致该功能类群真菌向腐生营养型转化.该功能群落以内生菌—凋落物腐生菌—土壤腐生菌—未定义的腐生菌(主要真菌来源是Mortierella属)和外生菌根—未定义的腐生菌—木材腐生菌(主要真菌来源是Myxotrichaceae科)为主.据报道,Mortierella属在降解芳烃和分解植物凋落物方面具有重要作用[37],Myxotrichaceae科真菌对纤维素以及可溶性酚类物质具有较好的降解作用[38].从土壤真菌功能类群变化情况来看,连栽可能会分解植物凋落物、纤维素等,导致真菌群落丰度降低,进而影响土壤养分循环速率和林木养分的吸收速率.
本研究结果表明:2代连栽对杉木人工林表层土壤真菌多样性的影响并不显著;对土壤主要优势菌群落种类、相对丰度以及群落结构的影响也不显著.共发现7种营养型功能类群,分别是腐生营养型、病原营养型、共生营养型、病原—腐生营养型、病原—共生营养型、腐生—共生营养型以及病原—腐生—共生营养型.其中腐生营养型真菌为主要功能类群,二代连栽对该营养类群真菌无显著影响,但会导致腐生—共生营养型功能类群真菌群落丰度极显著降低.土壤真菌多样性受土壤碳、钾元素含量的影响更大,丰富度受氮、磷元素的影响更大;腐生营养型真菌与全钾含量呈显著正相关,腐生—共生营养型真菌与有机碳含量呈极显著正相关,与全磷呈极显著负相关关系.2代连栽可能会降低植物凋落物、纤维素等分解速率,导致土壤养分循环速率降低.
猜你喜欢
南京林业大学学报(自然科学版)(2022年4期)2022-11-29 11:51:32
淡水渔业(2022年3期)2022-05-25 07:16:04
天津医科大学学报(2021年1期)2021-12-05 11:11:05
食品安全导刊(2021年32期)2021-11-27 15:31:06
食药用菌(2021年5期)2021-10-15 06:23:12
食品科学(2020年23期)2020-12-31 01:31:42
林业科学(2020年10期)2020-11-30 00:36:56
科学导报(2019年28期)2019-09-03 04:34:03
生物学教学(2017年9期)2017-02-18 07:01:53
中国医学影像技术(2017年11期)2017-01-16 12:54:05
