
低温胁迫对马铃薯幼苗抗寒相关生理指标的影响*
2022-10-05 12:46段景秀侯禄晓暴会会汪雪梅郑昊吉祝光涛郭磊
云南师范大学学报(自然科学版) 2022年5期
段景秀, 侯禄晓, 暴会会,汪雪梅, 郑昊吉, 祝光涛, 郭磊
(云南师范大学 生命科学学院,生物能源持续开发利用教育部工程研究中心,云南省马铃薯生物学重点实验室,云南 昆明 650500)
马铃薯(SolanumtuberosumL.)属于茄科茄属一年生草本植物,是世界第四大粮食作物,其营养价值高,适应力强且产量大.马铃薯性喜冷凉不耐寒,低温胁迫是影响马铃薯生长发育的重要因素,严重制约着马铃薯产业的健康稳定发展[1].在2016年鄱阳湖地区的霜冻天气中,马铃薯幼苗冻死率达75%~95%,减产幅度为40%左右,造成严重损失[2];2018年,由于受到寒潮影响,广西玉林的马铃薯未能达到商品薯标准,造成了严重的经济损失[3].
野生种马铃薯要比普通栽培种马铃薯更耐寒,能够进行冷驯化[4].马铃薯普通栽培种出苗后在小于7 ℃ 的低温下植株停止生长,-0.8 ℃ 时遭受冷害,-1.5 ℃ 时遭受冻害,-3 ℃ 时死亡;野生种S.acaule一般在 -4 ℃ 处理12 h后才出现严重冻伤或死亡[5].大田生产马铃薯主要是利用秸秆、培土、熏烟和地膜覆盖等措施来提高地温预防冻害,这些措施虽然有一定的效果,但在面积大或低温灾害频繁的情况下,这些方法具有一定的局限性,难以使用,即使在小面积范围内实施这些措施,也会因投入大量的人力和物力而增加成本,严重影响马铃薯种植的经济效益和产业发展.鉴于马铃薯霜冻抗性等重要性状一直难以得到改良,甚至成为世界性的难题[6],进一步研究马铃薯的抗寒性及其作用机制尤为重要,而探究冷害和冻害胁迫下马铃薯生理指标的变化就是其中的一个研究内容,对选育马铃薯抗寒种质具有重要的意义.
Solanumcommersonii(CM)是已报道的马铃薯资源中较抗寒的野生种[7],选取云南师范大学马铃薯科学研究院提供的5份抗寒野生种CM的材料和1份不抗寒栽培种DM进行抗寒鉴定以及4 ℃和 -4 ℃ 1 h、3 h、6 h和12 h的冷害和冻害处理,分析其中可溶性糖含量、可溶性蛋白含量、超氧化物歧化酶(SOD)活性以及过氧化物酶(POD)活性等抗寒相关生理指标的变化.以期为解析马铃薯抗寒性提供研究基础,为农业生产上选育马铃薯抗寒品种提供指导.
1 材料与方法
1.1 试验材料
供试马铃薯种质为云南师范大学马铃薯科学研究院前期研究筛选的抗寒野生种CM中的559、801、538、674和803,以及不抗寒栽培种Solanumphureja(DM).
1.2 试验设计
选取生长状态良好,长势整齐的苗期马铃薯材料559、801、538、674、803和DM进行低温胁迫处理.试验在光照培养箱与冷室中进行.试验设置6组处理:25 ℃(对照组),4 ℃,-4 ℃ 1 h、3 h、6 h和12 h(植株-4 ℃处理前进行为期6 d的4 ℃低温驯化),每组三株.试验设置三次重复.低温胁迫处理后,取第3和第4片功能叶混合后按需称重,用锡纸包裹放进液氮中速冻后置于-80 ℃超低温冰箱中备用[8].另取每种材料的马铃薯各9株在相同的处理条件下进行抗寒表型鉴定.
1.3 生理指标测定
采用考马斯亮蓝法[9]测定可溶性蛋白含量;采用苏州科铭生物技术有限公司提供的试剂盒测定可溶性糖含量;参照李玲《植物生理学模块实验指导》进行游离脯氨酸含量测定;采用愈创木酚法[10]测定POD活性;采用苏州科铭生物技术有限公司提供的试剂盒测定SOD活性.
1.4 数据处理
采用Excel 2010进行数据统计和处理,采用GraphPad Prism 9 制作柱状图和折线图,并利用SPSS软件进行差异显著性分析.
2 结果
2.1 马铃薯抗寒性鉴定
不抗寒栽培种DM在-4 ℃处理12 h后,叶片萎蔫,茎秆瘫软[11],全株死亡(图1B);野生种CM(以674为例)在-4 ℃处理12 h后,叶片稍卷,仅部分叶片有损伤(图1D),但还能恢复生长.CM中的其余4份材料与674表现一致.

25 ℃ -4 ℃ 12 h
2.2 低温胁迫对马铃薯生理指标的影响
2.2.1 低温胁迫对SOD活性的影响
SOD是一种自由基清除剂,可有效提高植物的抗寒能力.对低温胁迫下的6份马铃薯材料SOD活性进行检测,结果显示,抗寒种CM中的5份材料在低温胁迫下SOD活性呈现先降低后升高又降低的趋势,在-4 ℃处理6 h下达到峰值且显著高于其他时间点(图2A-E),其中559、801、538和674在4 ℃驯化后,SOD活性没有显著增强(图2A-D).而803在低温驯化后,SOD活性显著增加(图2E),推测低温驯化可能对803抗寒能力的提升有一定作用.不抗寒栽培种DM整体的SOD活性都低于其他材料,低温驯化后SOD活性也没有被诱导增加.说明SOD活性较高是CM具有较强抗寒能力的原因之一.
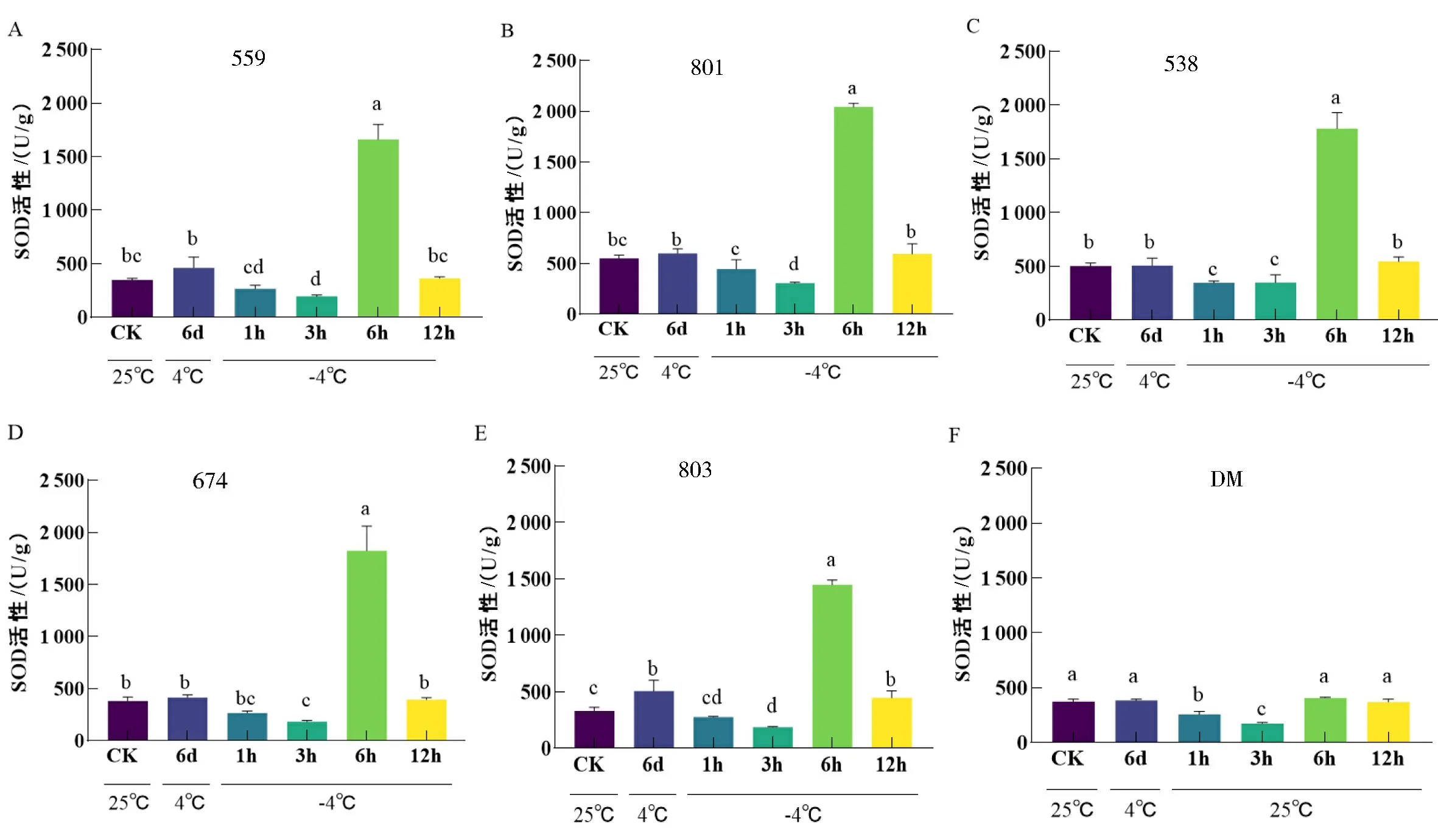
不同小写字母表示差异显著(P<0.05),下同.
2.2.2 低温胁迫对POD活性的影响
由图3可知,低温驯化后,抗寒种CM中的801、538和803以及不抗寒栽培种DM的POD活性都有不同程度的增加,559和674没有显著变化;在-4 ℃处理时,各材料POD活性总体呈上升趋势,其中803的POD活性增加最为显著,推测POD活性对803的低温抗冻性发挥了重要作用.

图3 低温胁迫下马铃薯POD活性
2.2.3 低温胁迫对可溶性糖含量的影响
可溶性糖作为一种渗透调节物质,在植物抗寒能力调节中发挥着重要作用.从图4可看出,抗寒种CM559、801和538在低温胁迫下,可溶性糖含量显著增加;而674、803和 DM无显著变化,但674和803的可溶性糖含量明显高于栽培种DM,推测高浓度可溶性糖含量能够提高马铃薯抗寒能力,也是抗寒材料的重要生理特征之一.
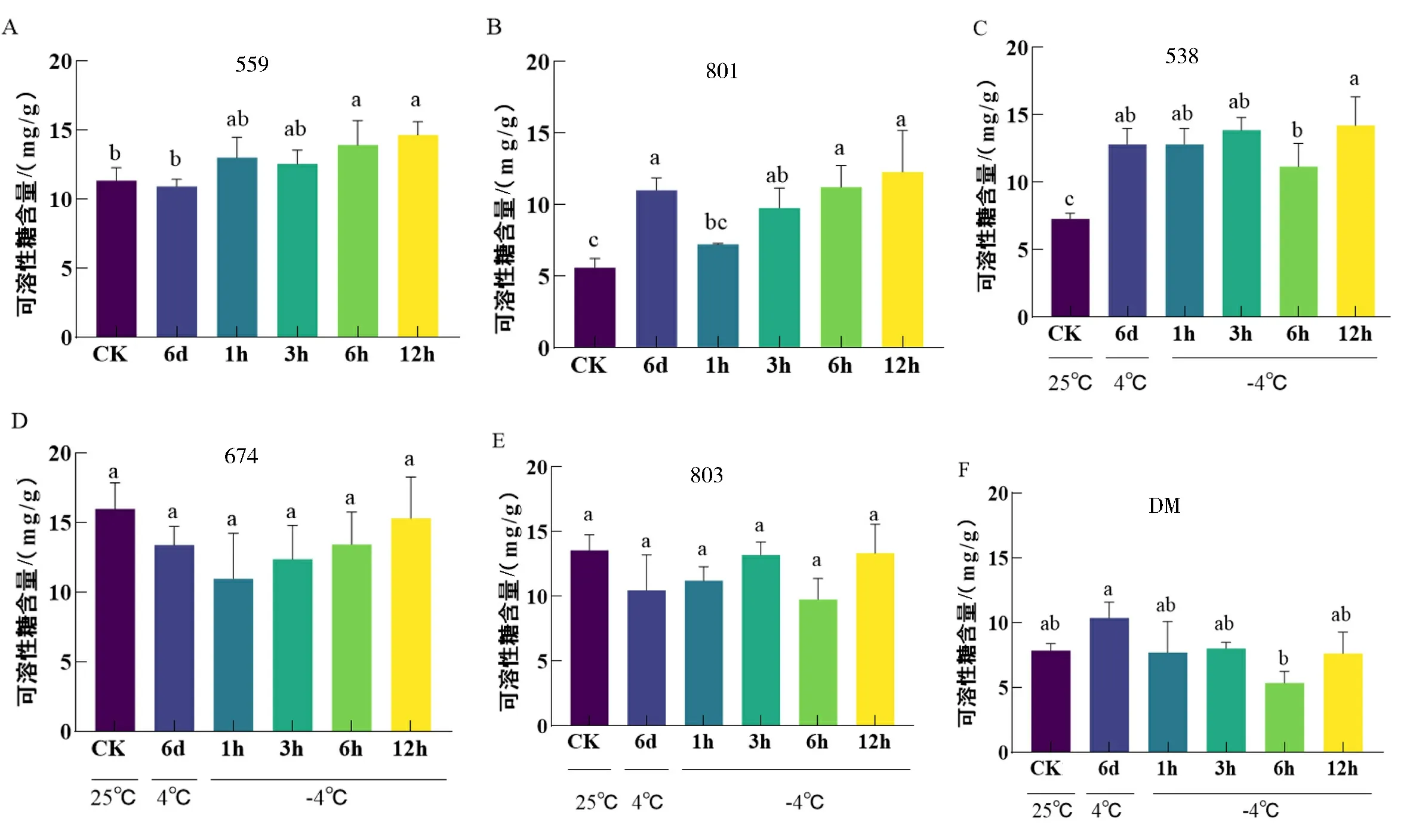
图4 低温胁迫下马铃薯可溶性糖含量
2.2.4 低温胁迫对可溶性蛋白含量的影响
可溶性蛋白有助于细胞维持渗透压平衡,增强抗寒性.抗寒材料559在4 ℃驯化后蛋白含量显著上升,-4 ℃胁迫后呈现先下降后上升的趋势(图5A),而其余材料在4 ℃驯化后可溶性蛋白含量显著下降或无变化(图5B-F).CM中除559外的材料大多在-4 ℃处理后的3 h、6 h或12 h时可溶性蛋白含量显著升高,特别是803,含量增幅最大(图5A-E).而不抗寒栽培种DM在低温胁迫后,可溶性蛋白含量显著降低(图5F).综上,抗寒种CM在低温胁迫下可溶性蛋白含量均增加,而不抗寒品种DM在低温胁迫下可溶性蛋白含量下降.
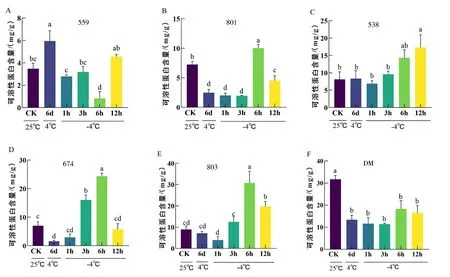
图5 低温胁迫下马铃薯可溶性蛋白含量
2.2.5 低温胁迫对脯氨酸含量的影响
作为最重要的有机渗透调节物质之一,植物组织中脯氨酸含量的增加可以提高植物的抗寒性.由图6可知,与室温对照组相比,低温驯化后各材料的游离脯氨酸均有一定程度的增加;总体上,游离脯氨酸含量呈现一个先增加后降低的趋势,其中抗寒种CM559和801在-4 ℃处理1 h游离脯氨酸增加最为显著,且低温胁迫下,CM脯氨酸含量显著高于DM.
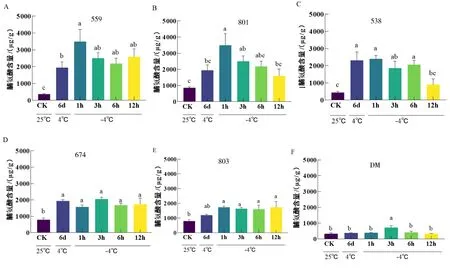
图6 低温胁迫下马铃薯脯氨酸含量
2.3 马铃薯抗寒种质综合评价
在所有材料中只有803在4 ℃低温驯化后,SOD活性显著增加,其他材料在冷驯化后均无显著变化.在低温胁迫条件下,各材料SOD活性呈现先下降后上升再下降的趋势,并且在-4 ℃处理6 h时达到峰值.其中,抗寒种CM 中801在处理前后SOD活性都高于其他材料,推测高SOD活性是其具有较强抗寒能力的原因之一(图7A).
在室温下,DM的POD活性相当于抗寒材料中的平均水平,并在低温诱导后呈现出与抗寒材料相似的变化趋势,只有在-4 ℃下12 h时活性下降,推测POD活性高低不是CM比DM更耐寒的原因之一.在低温胁迫下,抗寒种CM中803的POD活性最高(图7B).
在室温下674、803和559的可溶性糖含量较高,超过10 mg/g,在低温胁迫后变化不显著,而801、538和DM在未处理之前的可溶性糖含量较低,低于10 mg/g,其中抗寒材料801和538在低温胁迫后可溶性糖含量显著增加,而不抗寒材料DM可溶性糖含量在-4 ℃处理6 h时含量显著下降.以上结果说明CM中既有通过自身的高可溶性糖含量来提高抗寒性的材料,又有低温诱导可溶性糖含量增加来提高抗寒性的材料,且可溶性糖含量也是CM抗寒性强的原因之一(图7C).
低温胁迫后,除了559和538外所有材料的可溶性蛋白含量均在-4 ℃处理6 h时达到峰值,其中以抗寒种CM803增加最为显著.总体上,低温胁迫下CM与DM的可溶性蛋白含量相比无显著优势,推测可溶性蛋白对抗寒种CM的抗寒能力影响较小(如图7D).
低温胁迫下,6份马铃薯材料脯氨酸含量都呈现一个先增加后降低的趋势.且每种材料脯氨酸含量达到峰值的时间不同,如559在-4 ℃处理1 h时脯氨酸含量达到峰值,超过3 000 μg/g,801在-4 ℃处理3 h达到峰值,说明不同材料的脯氨酸含量对低温胁迫响应的模式有所不同.并且抗寒种CM中脯氨酸含量显著高于DM.推测高脯氨酸含量是影响抗寒种CM抗寒能力的重要因素之一(图7E).
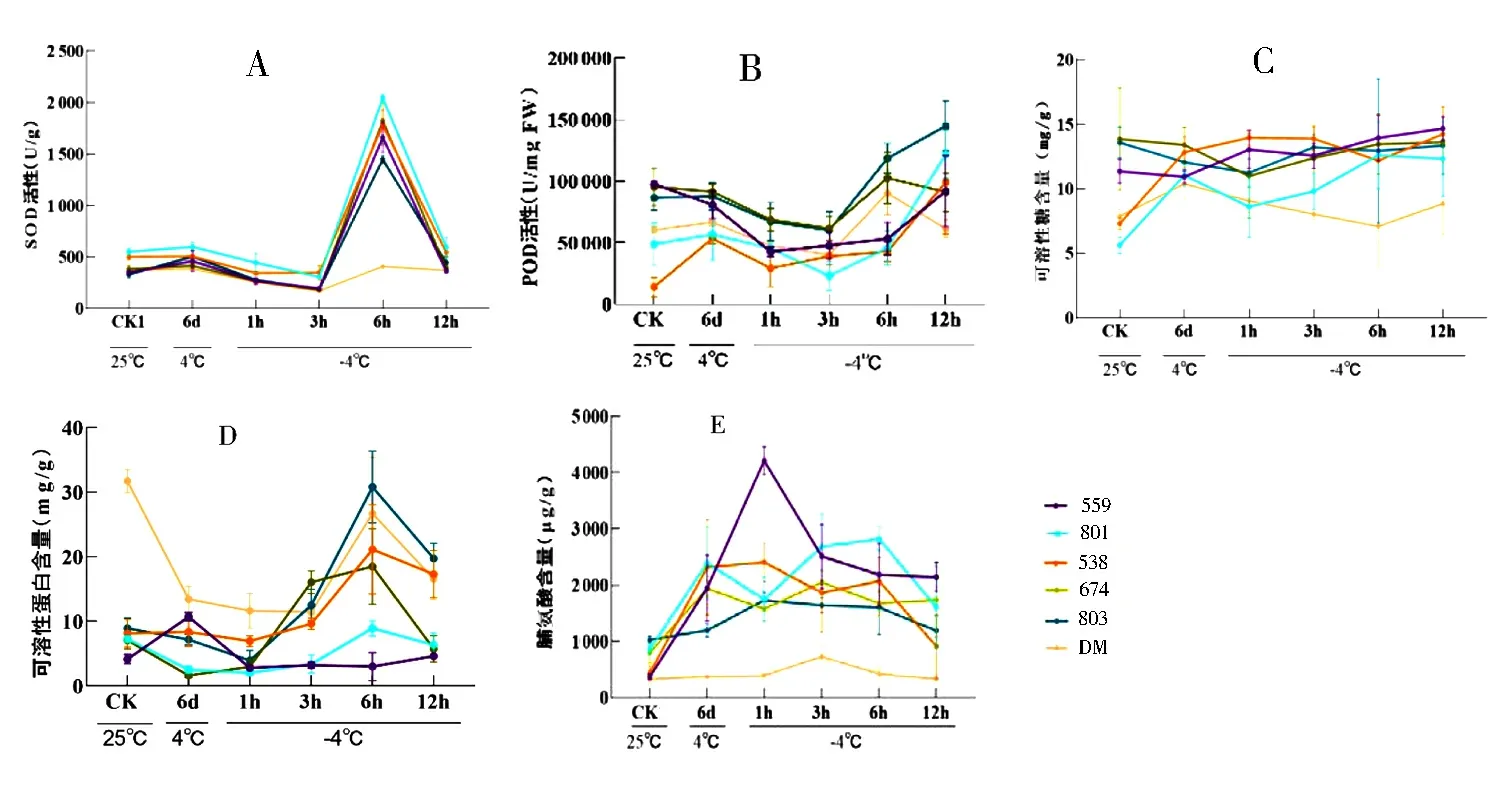
图7 低温胁迫下马铃薯各生理指标变化
3 结语与讨论
低温胁迫会对大多数作物产生伤害,影响产量,甚至造成死亡.然而,作物也会产生一系列反应抵御低温胁迫,在这个复杂的调控过程中,各个生理物质之间相互作用,相互影响[12].
植物面临低温胁迫时可通过产生大量的渗透调节物质来维持渗透压平衡,使其维持正常的形态和生理功能.研究结果显示,在4 ℃低温胁迫时,CM559可溶性蛋白含量升高最为明显.在-4 ℃低温胁迫时,随着胁迫时间延长,大部分材料中可溶性蛋白含量整体呈先上升后下降趋势,且在6 h或12 h时达到峰值,说明在该时间点可溶性蛋白响应最为强烈.5份CM材料的可溶性糖含量随低温胁迫时间的延长均有不同程度的增加,脯氨酸含量呈先增加后减少的趋势,说明5份马铃薯材料均会通过产生和积累渗透调节物质可溶性糖和脯氨酸来抵御寒冷.4 ℃驯化时,801、538和DM中可溶性糖含量增加,而6份材料的脯氨酸含量也均增加,说明低温驯化也能够提高马铃薯的抗寒性.材料559、674和803室温下就含有较高的可溶性糖含量,达到10 mg/g以上,材料801、674和803室温下的脯氨酸含量也较高,说明CM中有通过自身的高可溶性糖含量和高脯氨酸含量来提高抗寒能力的材料.不同材料马铃薯对低温胁迫的响应有所不同,所以脯氨酸和可溶性糖积累量的峰值出现的时间不同[8].低温胁迫后,马铃薯植株内的脯氨酸含量和可溶性糖含量变化趋势与白淼等[13]研究结果一致.
植物细胞内具有抗氧化酶保护系统,其中SOD和POD等能有效清除细胞内的活性氧物质,起到保护细胞的作用.实验表明,在4 ℃低温胁迫时,538中POD活性上升最为明显.在-4 ℃低温胁迫时,随着胁迫时间的不断延长,559、801、538和803中POD活性均呈上升趋势,表明细胞中POD活性不断增强以清除有毒物质活性氧,与孙佳平[14]等对甜菜以及郭扬[15]对玉米的研究结果一致.但674和DM中POD活性呈先上升后下降的趋势.低温驯化后803的SOD活性有明显的升高,推测低温驯化对提高803材料的抗寒性有一定的作用.低温胁迫6 h时,6份材料的SOD活性达到峰值,而且DM的SOD活性低于CM中的5份材料,说明SOD高活性是CM具有抗寒性的原因之一.
综上,不同材料马铃薯幼苗的生理物质对低温胁迫的响应模式不同,但多数材料在低温处理6 h 和12 h 时响应最强烈.CM既可通过自身的高可溶性糖含量和高脯氨酸含量来提高抗寒能力,又可通过诱导累积可溶性糖和脯氨酸及提高SOD活性来抵抗低温胁迫,从而提高其抗寒能力.
猜你喜欢
中老年保健(2022年3期)2022-11-21
蔬菜(2022年10期)2022-11-10
热带作物学报(2022年5期)2022-06-01
今日农业(2021年20期)2021-11-26
安徽化工(2021年3期)2021-05-29
山西农业科学(2020年10期)2020-10-16
棉花学报(2020年3期)2020-08-08
科学与财富(2019年15期)2019-10-21
园艺与种苗(2015年10期)2015-02-27
岷峨诗稿(2014年3期)2014-11-15
