
醉马草种子萌发对增温和降雨变化响应的研究进展
2022-09-29 11:05:38田伟涛张耀丹李超锋李庆伟徐井润董乙强吴淑娟袁晓波
草地学报 2022年9期
田伟涛, 张耀丹, 夏 江, 李超锋, 李庆伟, 徐井润, 董乙强, 吴淑娟, 袁晓波*
(1. 新疆昌吉州草原站, 新疆 昌吉 831100; 2. 兰州大学草地农业科技学院, 甘肃 兰州 730020; 3. 新疆瑞昶设计院有限公司, 新疆 乌鲁木齐 830000; 4. 新疆农业大学草业学院, 新疆 乌鲁木齐 830052)
近年来,全球气候变化已成为各地政府和科研工作者所关注的重点问题之一[1-2]。联合国政府间气候变化专门委员会(Intergovernmental Panel on Climate Change,IPCC)第五次气候变化评估报告中强调,温度升高和降水变化是当前全球气候变化最为显著的驱动因子[3]。据统计,自工业革命以来,人类活动加剧导致全球平均气温升高了约1.5℃[4]。与之相比,我国气候变暖形势更为严峻,过去50年,我国地表温度增加幅度较大,同期增温幅度明显高于全球平均水平,且表现出在未来仍会持续增加的趋势[5-6]。相比于全球平均气温不断升高的趋势,降水变化具有较大的不确定性[7]。Folland等[8]研究发现,过去一个世纪全球总降水量呈现出增加的趋势,并且暴雨和干旱等极端气候事件的发生频率亦有所增加。胡婷等[9]通过模型模拟的研究表明全球降水变化总体表现为“湿者愈湿、干者愈干”。总之,以温度升高和降水变化为主要特征的全球气候变化趋势愈发明显。
种子萌发是植物幼苗生长和群落建植等不可或缺的阶段,受温度和水分等非生物因子调控[10-11]。温度对于植物种子萌发具有至关重要的作用,其影响植物种子萌发进程的可能原因有以下两点:一方面,温度会影响种子内部细胞器和生物膜系统等,进而影响细胞内部正常生命活动的有序进行[12];另一方面,温度的高低亦影响植物发芽过程中对各种生理生化反应起关键作用酶的活性,当温度过低时,较低的酶活性不足以催化各种代谢反应的有效进行,而温度过高会导致酶变性失活。因此,温度对植物种子萌发的影响取决于温度的高低,并且不同的植物具有适宜其种子萌发的最佳温度和耐受限度[13-14]。水分也是调控植物种子萌发的重要非生物因子[15-16]。通常,植物种子成熟后会极度脱水,吸涨是种子萌发的第一步,只有吸涨后种子才能启动萌发过程,而不同植物启动其种子萌发时对外界水分含量的需求或高或低,这可能是植物躲避恶劣气候的一种生态对策[17-18]。水分胁迫会促使植物种子内活性氧的产生,高浓度的活性氧可通过氧化蛋白质干扰种子内部正常的细胞新陈代谢活动,从而对植物种子萌发进程产生一定影响[19]。温度和水分通过影响植物种子萌发特性,进而对田间出苗等农牧业生产活动产生深远影响,因此研究种子萌发如何响应温度和水分等非生物因子的变化对于农牧业生产具有重要实践意义[11,16,20-21]。
醉马草(Achnatheruminebrians)为多年生冷季型草本植物,属于禾本科(Gramineae)芨芨草属(Achnatherum),分布海拔为1 200~4 300 m,尤其以亚高山和高山草原分布较为广泛,是我国西北地区天然草原常见的一种多年生有毒草本植物[22-25]。众多研究发现醉马草种子中内生真菌带菌率可达100%[26-28],内生真菌侵染致使家畜采食醉马草后表现出食欲不振、神情呆钝、蹒跚如醉等中毒症状,对畜牧业生产造成巨大经济损失[25,29]。另外,内生真菌与醉马草之间可形成互利共生的关系[30],醉马草可为内生真菌提供生活场所,而内生真菌不仅能够提高醉马草对干旱、高低温、盐碱、重金属等逆境的耐受程度,亦能促进醉马草在逆境条件下种子的萌发[31-35]。目前,关于增温和降水变化对醉马草种子萌发影响的基础性研究已有众多报道,但对于种子萌发响应增温和降水变化的作用机理和分子机制尚不明晰,关于禾草内生真菌在调控种子萌发对增温和降水变化响应过程中扮演何种角色仍具有较大不确定性。鉴于此,本研究针对该领域国内外相关研究进展进行梳理总结并展望,以期为进一步深入开展醉马草防控相关研究提供基础资料和理论依据。
1 醉马草的特性以及国内分布概况
醉马草是我国天然草地常见的一种有毒草本植物,分布范围广泛,主要集中于海拔为1 200~4 300 m的地带,包括新疆、青海、内蒙古、宁夏、甘肃以及西藏部分地区(表1)[26]。相关研究表明,新疆地区醉马草的分布面积在1987年为40.0万公顷,1992年分布面积达到了53.3万公顷,5年期间分布面积增加了33.25%[36-37],局部地区醉马草的覆盖率甚至高达85%[38]。近年来,醉马草分布面积和覆盖率在不同地区呈扩增趋势[39]。例如,近期有研究通过建立生态位模型预测了醉马草在青海省的潜在扩散区域,结果表明,醉马草在青海省的潜在可扩散生存区域达到青海省土地面积的50%[40]。
醉马草分布面积的增加主要与其自身较强的适应性和抗逆性密切相关。首先,醉马草具有株体高(平均高度约100 cm),叶片质地偏硬,圆锥花序紧密呈穗状且分蘖数多(约100)等特点[39],其对光能和资源的获取能力强,具有更强的生态位优势;其次,醉马草种子产量高,且种子成熟后易脱落,可附着于放牧家畜表皮并进行传播,有利于其在草地中定植[34,38,41];此外,众多研究发现醉马草种子中内生真菌带菌率可达100%[26-28],且后续研究已发现2种与醉马草共生的内生真菌,包括甘肃内生真菌(Epichloёgansuensis)和醉马草内生真菌(E.inebrians)[27],其可提高醉马草对干旱、高低温、盐碱、重金属等逆境的耐受程度[23,31-35]。
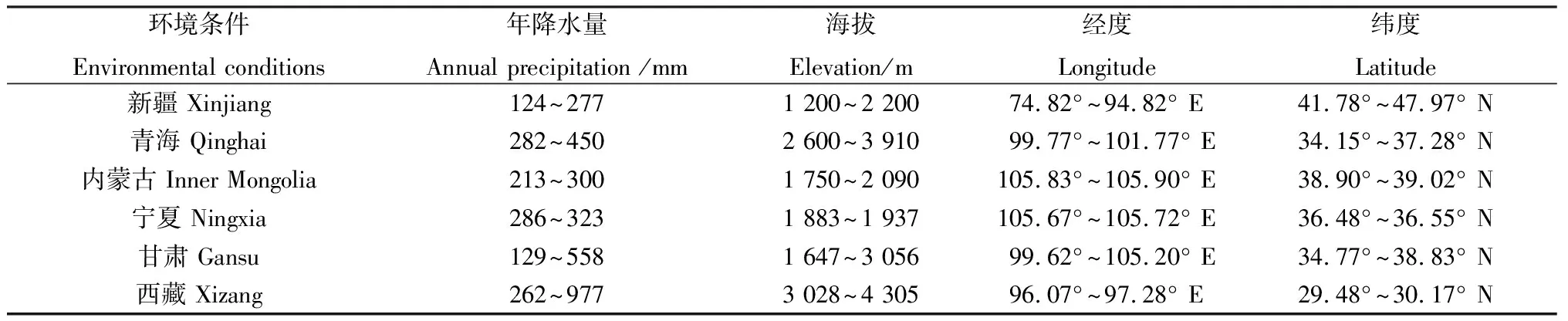
表1 醉马草在我国的主要分布区及环境条件[26]Table 1 The main distribution areas and environmental conditions of Achnatherum inebrians in China[26]
家畜采食醉马草后通常表现出食欲不振、神情呆钝和蹒跚如醉等中毒症状[25]。相关研究认为家畜采食醉马草后表现中毒症状与其芒刺刺伤家畜有关[42-43]。然而,有研究表明醉马草芒刺刺伤家畜多表现为伤口处形成小脓肿或刺伤眼角膜使得家畜失明,因此,物理性芒刺刺伤并非醉马草致毒的真正原因[44]。此外,相关学者通过旋转蒸发等方法从醉马草水提液中分离得到麦角新碱、麦角酰胺及异麦角新碱等多种生物碱,并确定醉马草大量产碱的原因是香柱菌属内生真菌的侵染,但这些研究并未进一步深入开展毒性试验,从而得到生物碱造成家畜中毒的机理[45-48]。后续有研究表明,麦角新碱及异麦角新碱这两种生物碱显著抑制了动物体内平滑肌细胞的正常生长,并且明确了麦角新碱及异麦角新碱对平滑肌细胞的半致死浓度[29]。至此,明确了家畜采食醉马草致其中毒的真正原因是醉马草与内生真菌共生后产生的生物碱对家畜的直接毒害作用[48-50]。
2 醉马草种子萌发对增温的响应
温度是植物种子萌发的重要调控因子,种子萌发初期需将种胚内所储存的蛋白质等大分子物质分解为氨基酸等可溶性小分子物质,以提供种子萌发时所需的能量物质[51]。这一过程需要各种酶类参与,酶的种类及含量受外界温度影响,进而直接影响种子的代谢状态和能量水平,从而调控种子的萌发[52]。因此,温度过低或过高均会对酶促反应如蛋白酶将贮藏蛋白质降解成氨基酸的过程产生不利影响,进而抑制种子萌发[13-14]。截至目前,关于温度对醉马草种子萌发影响的研究结果已有相关报道,且多采用室内人工气候箱模拟全球增温的方法进行研究。例如,一项对乌鲁木齐市南山的醉马草种子进行高温处理研究发现,随温度和处理时间持续,醉马草种子发芽率、发芽势、发芽指数和活力指数均表现出先升高后降低的趋势,且当75℃处理30 d时,醉马草种子完全不能萌发[53]。上述研究结果表明醉马草种子萌发对温度的响应存在最适温度值或范围[54]。尽管相关研究学者已对醉马草种子萌发的最适温度进行研究,但研究结果并不一致。例如,鱼小军等[23]和Ahmad等[55]的研究结果均发现,醉马草种子最适萌发温度为20℃。然而,一些研究结果表明,醉马草种子在25℃时萌发效果最好[37,56],造成上述研究结果不一致的原因可能是不同研究所使用的试验材料生境不同。此外,以上研究仅关注醉马草种子萌发对恒温的响应,而关于变温如何影响醉马草种子的研究较少。岳永寰等[57]的研究发现,醉马草种子在恒温15℃~35℃和变温20/10℃~40/30℃条件下均能较好地进行萌发。这些研究结果充分说明醉马草种子具有较为宽泛的萌发温幅,温度过低或过高均不利于其种子萌发[13-14]。
醉马草种子萌发对于低温和高温胁迫的表现不尽相同。相关研究表明,低温(5℃)条件下醉马草种子仍然能够萌发,而高温(75℃)条件下醉马草种子已经失活,不能萌发,这表明醉马草种子具有更强的耐低温能力[43,53]。主要因为低温环境致使醉马草种子进入休眠状态进而避开低温对其萌发产生不利影响;相较而言,温度过高导致醉马草种子内部与萌发相关的酶类失活,致使萌发过程中酶促反应不能正常进行,进而使其失活[14-16]。
醉马草种子萌发对温度的响应亦受母本环境的调控。例如,一项对高山草原、干旱草原和荒漠草原的醉马草种子的萌发试验表明,低温条件下高山草原的醉马草种子发芽率显著高于干旱草原和荒漠草原,而高温条件下干旱草原和荒漠草原的醉马草种子发芽率显著高于高山草原[58]。这可能因为母本长期所生长的环境不同,造成同种植物具有不同的温度敏感性,从而致使种子萌发对温度响应的分异性[54,59]。
综上所述,醉马草种子具有较为宽泛的萌发温幅,其在萌发过程中对温度表现出一定的耐受性和适应性,这也是醉马草分布面积和覆盖率扩张的原因之一。全球气候变暖在一定温度范围内会促进醉马草种子萌发,一旦增温幅度超过耐受范围,则会对醉马草种子萌发产生不可逆转的损害。
3 醉马草种子萌发对降水变化的响应
近年来,关于醉马草种子萌发对降水量变化响应的研究已有众多报道,多采用PEG 6 000模拟干旱胁迫和降水变化,但研究结论并不一致,包括正效应、中性效应和负效应。例如,一项使用PEG-6 000(聚乙二醇)模拟干旱胁迫对醉马草种子萌发进程影响的研究发现,低浓度PEG-6 000(5%~10%)可促进醉马草种子发芽,而高浓度PEG-6 000(15%)则抑制其发芽势和发芽指数等[60]。另有研究表明,轻微水分胁迫对醉马草种子萌发无显著影响[55]。此外,刘雪松[61]研究了采自甘肃榆中和内蒙古阿拉善的醉马草种子对水分胁迫的响应并发现,水分胁迫会抑制醉马草种子萌发,且随着干旱程度加剧,种子萌发受到的抑制作用越强。对于上述研究结果的差异性可归结于以下几方面原因:一方面,种子来源不同使得其对于水分胁迫的耐受程度有所差异,致使种子萌发对于水分变化的响应不一致[15];另一方面,不同研究设置的水分梯度不同也会影响试验结果[16,55,62]。
目前关于长期干旱胁迫去除后醉马草种子是否具有萌发活力的研究尚未见报道,而短期干旱胁迫进行复水处理后的醉马草种子萌发状况已有报道。相关研究表明,短暂干旱胁迫不会导致醉马草种子完全丧失活性,复水处理后,反而会促进种子萌发[37,55,60]。这说明醉马草种子具有很强的抗逆性,干旱环境导致醉马草种子被迫进入休眠状态,当处于合适的生长条件时停止休眠并开始萌发[16,55]。干旱时种子进入休眠状态是醉马草种子逐渐适应干旱环境的萌发策略,能确保其在生活周期的初始竞争中占据一定优势,从而使醉马草在天然草地植物群落中占据绝对的生长优势。
母本环境同样会影响醉马草种子萌发对降水变化的响应。位海玲等[60]的研究结果表明,干旱胁迫条件下采自高海拔区域(2 287 m)的醉马草种子发芽率明显小于采自低海拔区域(1 650~1 832 m)的种子发芽率。因此,母本生长于中低海拔区域的醉马草抗旱性高于高海拔区域。
综上所述,目前关于醉马草种子萌发对降水量变化响应的研究报道较多,但研究结论并不一致,这可能因为不同研究的水分梯度设置不同。因此,迫切需要建立统一的研究方法以厘清醉马草种子萌发对降水变化的潜在响应机理和过程。
4 禾草内生真菌调控种子萌发对温度和降水变化的响应
禾草内生真菌指在宿主禾草体内完成大部分或整个生活史且与宿主禾草形成互惠共生体的一类真菌[30,63-64]。截至目前,已证实在全球300多种禾草中存在内生真菌,并且从中分离鉴定了45个香柱菌属(Epichloё)内生真菌[65-69]。诸多研究发现我国北方天然草原的醉马草种子中内生真菌带菌率可达100%[26-28],且发现2种内生真菌存在于醉马草体内,分别是甘肃内生真菌和醉马草内生真菌(表2)[24,70]。众多研究结果表明,内生真菌能够增强醉马草对干旱、高低温、盐碱和重金属等逆境的耐受程度[23,31-35]。此外,醉马草-内生真菌共生体主要通过种子进行垂直传播,因此内生真菌的存在势必对醉马草种子萌发产生一定的影响[38,69]。

表2 2种与醉马草共生的内生真菌Table 2 Two endophyte symbiotic with Achnatherum inebrians
目前相关研究表明,高温、干旱等逆境条件下内生真菌能够促进宿主植物种子萌发,主要集中在高羊茅(Festucaarundinacea)、多年生黑麦草(Loliumperenne)、多花黑麦草(Loliummultiflorum)和野大麦(Hordeumbrevisubulatum)等草本植物[71-74],而关于内生真菌如何调控醉马草种子萌发对温度和降水变化响应研究相对缺乏。例如,不同低温胁迫对醉马草种子萌发影响的研究发现内生真菌可以提高醉马草种子的发芽率和发芽速度,促进胚根和胚芽的生长[43]。与之相似,柳莉等[71]的研究结果亦表明内生真菌能够显著增加低温环境下醉马草种子萌发能力。内生真菌促进醉马草种子对低温的耐受能力可能是以下4方面原因:首先,相比于未携带内生真菌的醉马草种子而言,体内存在内生真菌的醉马草种子更加的成熟饱满,种子内部储存的能量更为充足[73];其次,低温条件下内生真菌导致参与多个代谢途径的一系列基因发生了差异表达,进而提高了醉马草对低温环境的耐受程度[43];再次,内生真菌菌丝可以突破种皮并为醉马草种子萌发提供碳水化合物等能量物质[74];最后,内生真菌的次生代谢物中存在醉马草种子萌发所需的信号物质,例如菌根菌自身所分泌的Strigolactones是种子萌发的重要信号物质之一[43]。
研究表明,内生真菌可以促进干旱条件下醉马草种子萌发,且干旱胁迫越严重这种促进作用愈加明显[69]。此外,内生真菌对醉马草种子抗旱性的促进作用亦受其母本环境影响。例如,有研究结果发现,内生真菌对湿润地区醉马草种子抗旱性的促进作用强于干旱地区的醉马草种子[69]。这可能因为长期处于干旱地区,醉马草-内生真菌共生体已经形成了通过渗透调节和抗氧化保护系统等一系列生理生化反应诱导植物提高其抗旱能力的应对机制,而湿润地区水分并不是限制醉马草生长和扩繁的主要因素,使得内生真菌对于醉马草种子抗旱性的促进作用未得到激发[23]。
5 研究展望
截至目前,关于增温和降水变化对醉马草种子萌发影响研究已取得一定进展[10-11,22-25],但缺乏增温和降水变化对种子萌发影响的作用机理和分子机制研究。同时,禾草内生真菌在调控醉马草种子萌发对增温和降水变化响应过程中扮演怎样的角色仍具有较大的不确定性。基于此,建议今后从以下几方面进一步开展相关研究:
(1)降水变化不仅表现为降水量的变化,还体现在降水频度等格局变化方面[8]。在水分影响醉马草种子萌发的研究中,大多使用PEG-6000进行室内模拟干旱胁迫,关于醉马草种子萌发表现对降水频度等格局变化响应的研究尚未见报道。
(2)种子萌发受氧气、埋藏深度和水热等多种非生物因子及其交互作用共同调控,然而现有研究多数聚焦于单一环境因子,缺乏多种环境因子及其交互作用对醉马草种子萌发的影响研究。
(3)醉马草即可作为一种草原毒害草进行有效防控,亦可作为一种潜在牧草资源和抗逆先锋种进行深度开发利用,因此,阐明其在全球气候变化大背景下种群更新机制尤为重要。后续研究应系统阐明全球气候变化对醉马草种子萌发、幼苗生长和成苗特性的影响。
(4)应用微生物学、分子生物学和多组学等理论方法,进一步开展内生真菌对醉马草种子抗旱和耐高低温的作用机理研究,以期从分子水平深入解析内生真菌对逆境下醉马草种子萌发的作用机理。
猜你喜欢
黑龙江气象(2021年2期)2021-11-05 07:07:00
科学(2020年2期)2020-08-24 07:56:56
湖北农机化(2020年4期)2020-07-24 09:07:16
世界农药(2019年4期)2019-12-30 06:25:10
今日农业(2019年11期)2019-08-15 00:56:32
乡村地理(2018年2期)2018-09-19 06:44:02
小学科学(学生版)(2018年8期)2018-08-21 12:05:50
家教世界(2018年16期)2018-06-20 02:22:00
成都信息工程大学学报(2016年6期)2016-06-01 12:10:06
微生物与感染(2015年5期)2015-12-08 07:03:20
