
玉米籽粒主要品质性状的分子遗传研究进展
2022-09-29 11:16王丽珊张建国曹士亮杨耿斌李文跃王成波段雅娟欧英卓刘长华
中国农学通报 2022年24期
王丽珊,张建国,于 滔,曹士亮,杨耿斌,李文跃,王成波,段雅娟,欧英卓,赵 晴,刘长华
(1黑龙江大学现代农业与生态环境学院,哈尔滨 150080;2黑龙江省农业科学院玉米研究所,哈尔滨 150086)
0 引言
玉米是全球三大粮食作物之一,主要被应用于食品、医疗、能源和工业等方面。2020年中国玉米总产量约为2.6亿t,占全球总产量的23%[1]。随着社会经济的快速发展,农产品需求剧增,玉米育种在追求高产的同时,对籽粒的绿色、优质和安全提出了更高的要求[2],开展玉米籽粒品质性状基础理论研究能够推动特用玉米新品种的研发,对保障中国粮食安全和适应消费需求具有重要意义[3]。
玉米籽粒品质性状主要包括:蛋白质、淀粉、油分及赖氨酸等[4]。目前,育种家多是通过分析仪器进行早代系的品质选择[5]。但是,玉米品质性状绝大多数是由微效多基因控制的数量性状[6],采用半同胞轮回等常规育种方法提升玉米品质,易降低遗传多样性,存在育种效率低,周期长等问题[7-8]。随着生物技术的快速发展,将分子标记与转基因技术相结合,应用于玉米籽粒品质性状进行遗传改良已经成为可能[9]。本文通过对国内外玉米籽粒品质性状的QTL定位、分子标记辅助改良和候选基因克隆及转基因技术应用的相关研究进展进行了综述,以期为玉米籽粒品质性状的分子设计育种提供参考。
1 玉米籽粒主要品质性状的QTL定位
在利用分子标记进行基因定位的基础上构建高密度遗传图谱,可以实现对玉米籽粒品质性状的分子遗传研究[10]。国内外研究人员应用不同的试验材料和研究方法(表1),对玉米品质性状遗传开展了大量研究,鉴定和定位了一批与玉米籽粒品质性状相关的QTL。
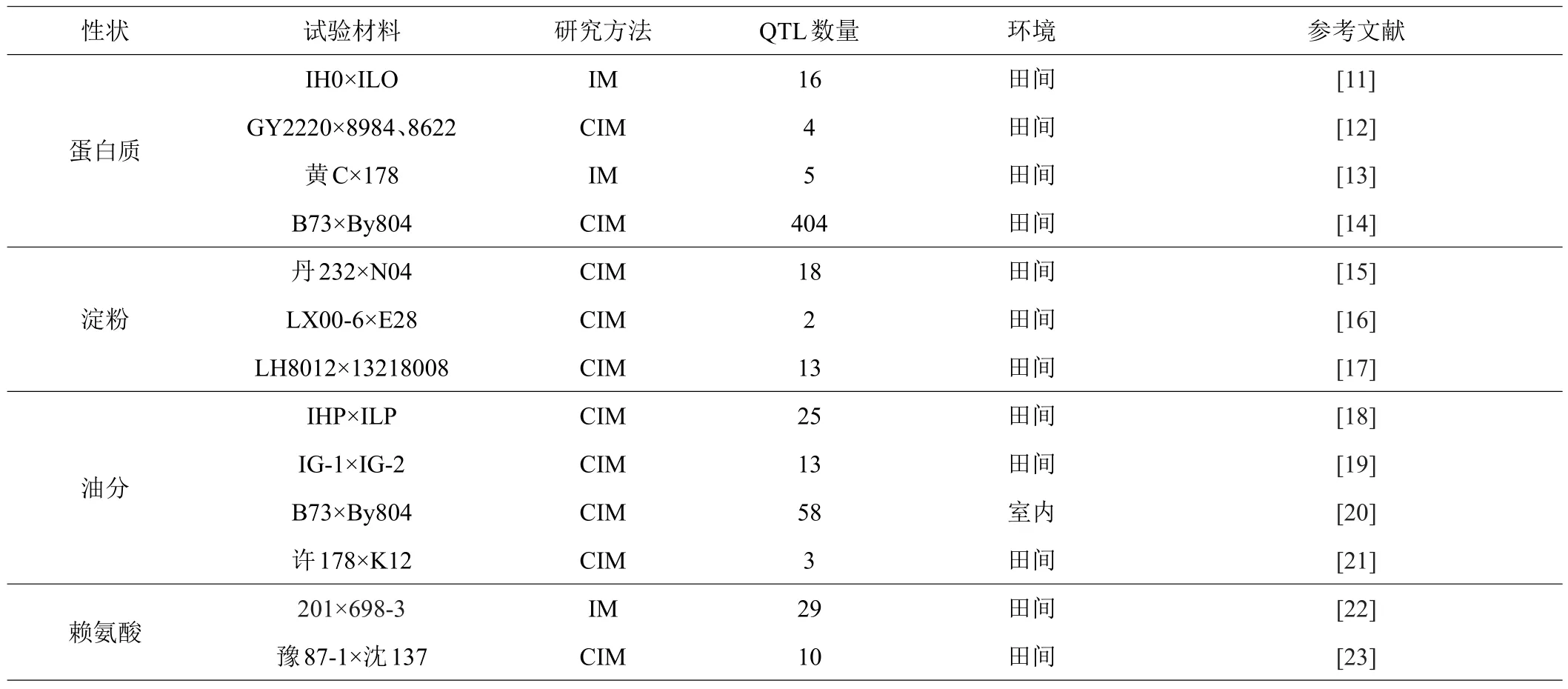
表1 玉米籽粒品质性状的QTL研究
1.1 玉米籽粒蛋白质含量的QTL定位
BERKE等[11]对IHO×ILO群体进行RFLP标记,共发现16个蛋白质含量相关QTL。LI等[12]以高油玉米和普通玉米为材料进行研究时发现4个只与蛋白质含量相关QTL,综合4种品质性状的检测结果发现,蛋白质和淀粉含量存在6个共同位点。孙海艳等[13]通过区间作图法对3种不同环境条件下的玉米材料进行全基因组QTL扫描发现5个与蛋白质含量相关的QTL,分布在第1、2、6、8和10染色体上。仅有1个QTL在3个环境中都能检测到,其表型变异方差介于10.0%~17.8%之间。说明环境是影响籽粒品质QTL表现的重要因素,这些在多个环境中稳定表达的QTL为深入开展分子标记辅助育种研究提供了理论依据。邓敏[14]对玉米自交系的关联群体和3个重组自交系群体进行研究,共鉴定到了404个氨基酸性状相关QTL,3个自交群体解释的表型变异平均为9.03%、9.39%和10.15%。说明群体间相同氨基酸性状变异是存在差异的,可为后续玉米氨基酸的遗传改良提供遗传资源。
1.2 玉米籽粒淀粉含量的QTL定位
玉米籽粒淀粉含量受基因型和环境的共同影响。张中伟[15]以普通玉米自交系与优良爆裂玉米自交系为材料,在3种环境条件下共检测到18个与淀粉含量相关的QTL位点,第5条染色体上具有最多的相关位点,共有5个QTL,其中qCT-5-2在3种环境下的单个贡献率为9.1%~13.4%,认定其为主效QTL。裴玉贺[16]在对LX00-6×E28的F2:3群体研究时,利用复合区间作图法检测到2个与淀粉含量有关的QTL,分布在第1条和第8条染色体上,二者解释表型变异率分别为7.8%和8.3%,第8条染色体上的QTL作用方式为部分显性。2个QTL加性效应值均是正值,分别为0.55和0.40,说明其作用方向是增加籽粒淀粉含量。赵丹[17]以自交系LH8012和13218008组配得到的F2:3家系群体为材料进行两点田间试验,共检测到13个与籽粒淀粉含量相关的QTL,分布于第1、4、5、8和9号染色体,位于区间umc1348~umc1990的QTL具有最高表型贡献率,为15.3%,可以将其看作主效QTL。
1.3 玉米籽粒油分含量的QTL定位
GOLDMAN等[18]利用RFLP位点进行群体标记时,共发现25个与油分含量显著相关的位点,加性效应和显性效应的位点个数分别为11个和4个。MANGOLIN等[19]运用复合区间作图法对F2:3群体进行QTL分析时发现,共有13个与油分含量显著相关的位点,QTL对表型变异的解释范围为2.35%~8.40%,除第8和10条染色体外,其余染色体上均能检测到与油分含量相关QTL。YANG等[20]以普通玉米B73和高油系玉米By804为材料,在26个基因组区域共鉴定出58个籽粒含油量及其组成性状QTL,其中包括8个主效QTL。玉米籽粒品质除受材料和环境影响外,还可能与肥料有关,为探究这一问题,兰天茹等[21]以许178×K12重组自交系群体为材料,进行施氮肥与不施氮肥两种处理。其中,施氮处理下只在第9条染色体上检测到1个油分含量相关QTL,贡献率为8.23%;不施氮肥处理时共在第1条和第6染色体上检测到2个相关位点,贡献率分别为9.32%和9.08%。两种氮素水平下没有共同存在的QTL位点,说明在此试验处理下,玉米籽粒油分含量受环境因素影响较小。
1.4 玉米籽粒赖氨酸含量的QTL定位
赖氨酸是玉米中重要的氨基酸,其含量高低与玉米籽粒品质紧密相关。赵丹[17]在对F2:3家系群体进行玉米籽粒赖氨酸QTL定位时,共检测到5个相关QTL,位于第5、8、10染色体上,与以往赖氨酸含量相关研究比较,未检测到相同或相近的染色体bin位点。石海春等[22]在对玉米自交系201×1698-3组合的F2:3群体进行研究时,在检测到的35个与品质相关QTL位点中,与赖氨酸含量相关的QTL为2个,分布于第2、4号染色体上,两个QTL均表现超显性效应,表型贡献率分别为11.8%和5.9%。张珍珍[23]在对Pop1和Pop2两个RIL群体进行研究时共检测到与玉米籽粒赖氨酸含量相关的10个QTL,这些QTL分别位于Pop1的第1、2、5、7、8染色体上以及Pop2的第5、9染色体上,可以作为后续分子标记辅助选择的位点来进行后代前景选择,从而提高玉米育种效率。
2 玉米籽粒主要品质性状的分子标记辅助改良
利用传统常规的玉米育种方法培育新品种时,选择依据通常为表型性状,而分子标记辅助选择可直接选定目的性状的基因型,加快玉米育种进程[24-25]。宁丽华等[26]开发了与高直链淀粉含量有关的玉米ae基因的功能标记。该功能标记被广泛应用于杂合基因型和个体纯合基因型个体的鉴定,能够精确筛选出高直链淀粉含量玉米品种,从而提高育种效率。ZHENG等[27]在玉米fatB基因的第六外显子中插入一个鸟嘌呤核苷酸后形成了一个截短形式的fatB基因,该基因能够降低籽粒中饱和脂肪酸的含量。玉米fatB基因功能标记可应用于低饱和脂肪酸含量品种的培育,以满足不同的市场需求。
2.1 分子标记在品质改良中的应用
分子标记以遗传物质变异为基础,表现为DNA水平遗传多态性。常见的分子标记有RFLP、SSR、SNP等。利用分子标记获得的信息可对亲本进行质量性状和数量性状的辅助选择[28]。
20世纪60年代MERTZ等[29]研究结果表明玉米opaque-2基因可大幅度增加籽粒赖氨酸的含量。国际玉米和小麦改良中心应用opaque-2基因培育出了优质高蛋白玉米品种QPM[30]。而后中国研究学者宋敏等[31]运用SSR分子标记方法,以phi057为引物,对QPM高代系opaque-2基因频率进行检测时发现opaque-2位点上的基因发生了突变,说明常规育种方法无法保证基因的纯合性,可利用SSR分子标记提前对早代系进行检测以准确筛选高蛋白含量玉米后代。张文龙[32]选用两个高赖氨酸玉米自交系和一个糯玉米自交系组建了两个回交群体,利用o2和wx基因内进行SSR分子标记以及o16基因内的连锁标记,测定籽粒赖氨酸含量,最终获得了不同隐性突变基因聚合的材料。鲁守平等[33]利用连锁SSR标记,采用回交育种方法,把提高籽粒油分的主效QTL转至低油分材料By804自交系中,最终使得3个自交系的改良系平均含油量增加0.58%。王长进等[34]在对甜玉米进行SNP分子标记时,共检测到20个与油分含量相关的SNP位点,位于第5条染色体上的qFAT-5-1表型贡献率最高为17.06%,与油分含量显著关联。
2.2 高通量测序在品质改良中的应用
高通量测序技术作为转录组测序的代表,在生物基因组学中应用广泛。根据高通量测序技术所得到的数据,可以在全基因组范围内找到玉米籽粒物质所累积的有关功能基因,这对于作物育种以及改良具有极其重要的意义[35]。何骋[36]对温带和热带玉米两个类群的籽粒进行转录组测序,利用XP-EHH信号选择方法在两个类群中分别鉴定到了731个和421个选择区域,而后对受选择的基因进行功能注释,最终发现油分合成途径的相关基因GRMZM2G083195和GRMZM2G061885以及与脂肪酸延长相关基因GRMZM2G152105和GRMZM2G003501均在温带玉米中受到了强烈的选择,说明人工选择可在特用玉米研发中起到关键作用。党仁芳[37]在试验前期预测zmmads11突变体能够参与玉米籽粒淀粉合成,并通过多代回交和自交得到纯合的突变体,对野生型和纯合突变体进行转录组测序发现:zmmads11纯合突变体中有2个参与淀粉和糖代谢途径的基因上调,分别为β-葡萄糖苷酶和海藻糖磷酸酶,从而推断zmmads11突变体中淀粉含量与代谢途径中相关酶基因表达量成正比。由于转录调控首先是调节相关酶基因的表达水平,进而调控淀粉的生物合成,此研究针对玉米淀粉生物合成过程中的分子调控机制,为未来培育高淀粉玉米提供理论依据。
2.3 关联分析在品质改良中的应用
关联分析在分子标记的基础上,选择遗传变异相对广泛的自然群体,结合统计学分析,最终鉴定出影响目标性状的关联位点和基因,为后续玉米品质基因克隆和转基因研究提供候选基因,加快育种进程[38]。董青松[39]以吉林省80份玉米自交系为试验材料,对1490007个SNP进行全基因组关联分析(表2)。研究结果表明与蛋白质含量显著关联的SNP位点数最多为11个,其中位于染色体框3.04的SNP最高解释了85.76%的表型贡献率,在连锁不平衡区域内共挖掘出13个候选基因。李长生[40]利用碘染法对464份玉米自交系的直链淀粉含量和重测序基因型数据进行全基因组关联分析结果表明:位于第9号染色体SNP位点与直链淀粉含量显著相关。该SNP信息为C或T,含有SNP T信息的玉米直链淀粉含量普遍偏低,所以该SNP位点可以作为分子标记对糯玉米进行选育。

表2 吉林省80份玉米核心自交系全基因组关联分析结果
3 基因克隆及转基因进展
3.1 玉米籽粒品质性状相关的基因克隆
中国关于玉米籽粒品质基因克隆的研究位于世界前沿。岳静[41]从棉花基因组和谷子cDNA中克隆得到了高赖氨酸蛋白基因GHLRP和SiREM6,为中国玉米品质的改良提供了基因资源。国外研究方面,YANG等[42]克隆了玉米籽粒淀粉合成相关候选基因ZmNAC36和ZmbZIP91,发现这2个基因对玉米籽粒淀粉合成途径具有调控作用。QI等[43]对Dof(DNA binding with one finger)家族基因中的ZmDof3进行功能验证,发现该基因会造成籽粒中所含的淀粉含量下降以及部分糊粉层有一定的缺陷。这一验证表明在胚乳发育过程中ZmDof3作用十分显著,为后续玉米品质基因的克隆提供理论依据和技术支持。
3.2 转基因技术应用于玉米品质性状
自1995年第一株转基因玉米品种获得商业许可后,国内外开展了大量转基因玉米研究[44]。通过转基因技术将外源基因导入玉米中,可实现基因的优质转化,以满足人们对玉米品质的特定需求[45]。研究者通过转基因技术改良了玉米籽粒中直链淀粉、赖氨酸和微量元素含量(表3)。
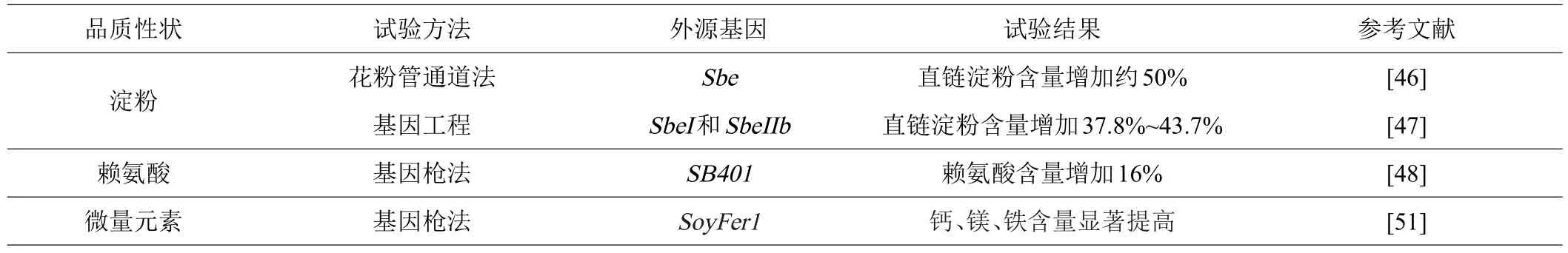
表3 转基因技术应用于玉米品质研究进展
柴晓杰等[46]利用花粉管通道法将玉米淀粉分支酶基因导入植株后,籽粒中直链淀粉的含量提高约50%。JIANG等[47]向玉米中导入SbeI和SbeIIb后,玉米直链淀粉含量增加约40%。
蛋白质中第一限制性氨基酸是赖氨酸,它是评价玉米营养价值的重要指标。孙学辉等[48]将富含赖氨酸的外源蛋白基因SB401导入玉米自交系后,赖氨酸含量提高16%。WU等[49]向玉米中导入拟南芥赖氨酸转运RNA,使得赖氨酸含量提高到26%。
ALURU等[50]将外源基因crtB和crtI分别与γ-zein启动子结合后转入玉米植株中,均能够高效表达。ZHU等[51]利用基因枪法将5个类胡萝卜素基因导入玉米植株后能够成功表达。有研究表明,将大豆铁蛋白SoyFer1基因导入玉米植株后,籽粒中的微量元素含量,例如钙、镁、铁等都大幅度提高。
4 展望
目前国内玉米籽粒品质性状的研究取得了一定的进展,但很多研究工作还处于起步阶段。因此,今后的研究可以考虑从以下几个方面开展:
(1)目前国内外还未充分利用玉米品质基因资源,后续应加强筛选优良等位基因,以推动特用玉米的育种工作进展。
(2)运用区间作图法和复合区间作图法进行QTL初级定位时,存在较大的置信区间,定位结果受作图群体和图谱饱和度影响较大,应结合多种方法对QTL定位进行全面系统研究,提高QTL定位的准确性,为分子标记辅助选择奠定基础。
(3)目前应用最广泛的分子标记方法是SNP和SSR。SNP标记稳定性好且可实现快速高通量检测,但投入花费较高;SSR标记操作简便且多态性高,但无法实现高通量。所以应开发新的功能标记,达到高通量且低成本的检测目的。
(4)目前国内外研究学者在玉米育种工作中主要侧重于培育高产量新品种,而忽略了品质性状的研究,今后应改进育种方法和分析鉴定技术,提高后代遗传稳定性的同时缩短育种年限,走玉米优质且高产的可持续发展路线。
猜你喜欢
中国农业科学(2022年16期)2022-09-19
中国农业科学(2022年15期)2022-08-09
中国农学通报(2022年13期)2022-05-31
江苏农业科学(2022年6期)2022-04-15
作文·小学低年级(2021年10期)2021-01-25
电脑报(2020年40期)2020-11-06
电脑知识与技术(2018年19期)2018-11-01
科学种养(2017年6期)2017-06-13
农民致富之友(2016年5期)2016-10-21
农村百事通(2016年8期)2016-05-14
