
蝴蝶兰属间杂交育种研究进展
2022-09-23 04:32陈和明吕复兵肖文芳
中国农业大学学报 2022年9期
陈和明 吕复兵 李 佐 肖文芳
(广东省农业科学院 环境园艺研究所/广东省园林花卉种质创新综合利用重点实验室,广州 510640)
蝴蝶兰(Phalaenopsis
spp.)是兰科(Orchidaceae)蝴蝶兰属植物,是我国最重要的年宵花卉之一,也是世界上产销量最大的兰花品种,因其花形奇特、花色丰富、色泽艳丽、花序整齐和花期长等特点,成为全世界广泛流行的热带兰花,深受国内外消费者喜爱,被称为“兰花皇后”。1887年,自英国皇家园艺学会(Royal Horticultural Society,RHS)正式注册登录第1个蝴蝶兰属人工杂交种以来,蝴蝶兰经过长达130多年的杂交育种取得了巨大的育种成就。截至2020-12-31,已有37 043个蝴蝶兰杂交组合在英国皇家园艺学会上正式登录,包括尖囊兰属(Kingidium
)、五唇兰属(Doritis
,又称朵丽兰属)的杂交种及其与蝴蝶兰属的属间杂交种一起归并为蝴蝶兰属。因此,蝴蝶兰的杂交育种极大地推动了蝴蝶兰产业的发展。但长期以来蝴蝶兰杂交育种主要集中于属内种间或品种间杂交,与近缘属间的杂交利用较少,导致蝴蝶兰栽培品种存在遗传基础较窄、多数蝴蝶兰没有香味、蓝色花缺乏及抗性较差等系列问题。而近缘属中钻喙兰属(Rhynchostylis
)和指甲兰属(Aerides
)等的花朵多数有香味、万代兰属(Vanda
)中蓝色花表现极佳和火焰兰属(Renanthera
)花色艳丽等性状均有利于蝴蝶兰的改良,且多数近缘属抗性较强。因此,本研究拟对蝴蝶兰近缘属种质资源、属间杂交育种情况和属间杂交存在的问题等方面进行分析,结合最新的研究成果,提出现有问题的解决方案,以期为我国蝴蝶兰属间杂交育种提供技术支撑。1 蝴蝶兰近缘属种质资源
在英国皇家园艺学会正式注册登录且与蝴蝶兰属间杂交成功的近缘属有19个,报道出与蝴蝶兰属间杂交成功的萼脊兰属(Sedirea
)和风兰属(Neofinetia
)共2个(表1)。因此,当前蝴蝶兰属间杂交成功的近缘属主要有21个。表1 蝴蝶兰近缘属分布及其种数
Table 1 Distribution and species number of related genera of
序号Code蝴蝶兰近缘属Related generaof Phalaenopsis分布Distribution优良特性Excellentcharacteristics全球原产种数No. ofnativespecies inthe world我国原产种数No.of nativespeciesin China1万代兰属Vanda我国和亚洲其他热带地区花色艳丽且植株抗性较强4092火焰兰属Renanthera东南亚至热带喜马拉雅花色艳丽且植株抗性较强1533钻喙兰属Rhynchostylis热带亚洲多数有芳香、多花且植株抗性较强624指甲兰属Aerides我国南方至东南亚多数有芳香、多花且植株抗性较强2045槽舌兰属Holcoglossum主要分布于我国花数较多且植株抗性较强886狭唇兰属Sarcochilus未记载未知未知未知7蜘蛛兰属Arachnis东南亚至新几内亚岛等花数较多且植株抗性较强131
表1(续)
序号Code蝴蝶兰近缘属Related generaof Phalaenopsis分布Distribution优良特性Excellentcharacteristics全球原产种数No. ofnativespecies inthe world我国原产种数No.of nativespeciesin China8拟万代兰属Vandopsis中国至东南亚等花色艳丽且植株抗性较强529凤蝶兰属Papilionanthe中国南部至东南亚花色艳丽且植株抗性较强11210钗子股属Luisia热带亚洲至大洋洲花朵较小且植株抗性较强501011筒叶蝶兰属Paraphalaenopsis特产于婆罗洲花数较多且植株抗性较强4012盆距兰属Gastrochilus亚洲热带和亚热带地区花朵较小且植株抗性较强472813毛舌兰属Trichoglottis东南亚、新几内亚岛和澳大利亚等 花朵较小且植株抗性较强60214安格兰属Angraecum马达加斯加岛、热带非洲和南非花朵较多且植株抗性较强未知015闭距兰属Cleisocentron中国及其它地方花朵较小且植株抗性较强未知1616钻柱兰属Pelatantheria热带喜马拉雅经印度到东南亚花朵较小且植株抗性较强5317阿梅兰属Amesiella主要分布于菲律宾花朵较多且植株抗性较强未知018蛇舌兰属Diploprora南亚的热带地区花朵较小且植株抗性较强2119漏斗兰属Eurychone未记载未知未知未知20萼脊兰属Sedirea中国、日本和朝鲜半岛南部小花、多数有芳香且植株抗性较强2221风兰属Neofinetia东亚小花、多数有芳香且植株抗性较强22
注:数据主要来源于《中国植物志》。
Note: The data come mainly from .
2 蝴蝶兰与近缘属杂交育种概况
2.1 属间杂交组合登录情况
蝴蝶兰属间杂交组合最早于1931-01-01在英国皇家园艺学会上正式注册登录,其杂交组合为Phal
.amabilis
×Vanda
suavis
,杂交属名为Vandaenopsis
,登录种名为Ferrierensis
,截至2020-12-31,基于表1(序号1~19)且在RHS上登录成功的蝴蝶兰属间杂交组合共有546个(表2)。根据英国皇家园艺学会兰花品种登录规定,相同亲本的杂交组合(无论是正交,还是反交)只能作为1个集体杂交种(Grex)或1个杂交群体进行登录,但从1个杂交群体中可以选育出多个优良单株,通过组培克隆繁殖获得优良株系,进而选育出多个优良品种。因此,蝴蝶兰属间杂交品种或杂交后代优良材料的实际数量要多于RHS上登录的杂交组合数。从F杂交组合总数排名来看(表2),排名前3的万代兰属、火焰兰属和钻喙兰属其杂交组合数分别为243、163和41个,合计杂交组合数为447个,占19个近缘属杂交组合总数的81.9%,表明万代兰、火焰兰和钻喙兰这3个属在蝴蝶兰属间杂交育种中占据了重要位置,为蝴蝶兰属间杂交育种史作出了重大贡献;排名第4、5、6和7的指甲兰属、槽舌兰属(Holcoglossum
)、狭唇兰属(Sarcochilus
)和蜘蛛兰属(Arachnis
)其杂交组合数分别为16、15、14和11个,杂交组合数合计为56个,占19个近缘属杂交组合总数的10.3%。表2 蝴蝶兰与RHS上登录的19个近缘属杂交育种情况
Table 2 Hybrid breeding of . with 19 related generaregistered on RHS
序号Code蝴蝶兰近缘属Relatedgenera ofPhalaenopsis远缘杂交属名Genus nameF1杂交组合数及所占比例 Number and proportion of F1 hybrids近缘属作母本/个Relatedgeneraas femaleparent近缘属作父本/个Relatedgeneraas maleparent总数/个Total总数排名Rank作母本时所占比例/%Proportionas female作父本时所占比例/%Proportionas male蝴蝶兰杂交种为亲本所占比例/%Proportionas parent ofPhalaenopsishybrids1万代兰属VandaVandaenopsis143100243 158.841.255.12火焰兰属RenantheraRenanthopsis134 29163 282.217.876.73钻喙兰属RhynchostylisRhynchonop-sis 1 40 41 32.4097.665.94指甲兰属AeridesAeridopsis 10 6 16 462.537.550.05槽舌兰属HolcoglossumHolconopsis 11 4 15 573.326.753.36狭唇兰属SarcochilusSarconopsis 12 2 14 685.714.328.67蜘蛛兰属ArachnisArachnopsis 10 1 11 790.99.1027.3
表2(续)
序号Code蝴蝶兰近缘属Relatedgenera ofPhalaenopsis远缘杂交属名Genus nameF1杂交组合数及所占比例 Number and proportion of F1 hybrids近缘属作母本/个Relatedgeneraas femaleparent近缘属作父本/个Relatedgeneraas maleparent总数/个Total总数排名Rank作母本时所占比例/%Proportionas female作父本时所占比例/%Proportionas male蝴蝶兰杂交种为亲本所占比例/%Proportionas parent ofPhalaenopsishybrids8拟万代兰属VandopsisPhalandopsis 4 4 8 850.050.075.09凤蝶兰属Papilionan-thePapilaenop-sis 4 4 8 850.050.012.510钗子股属LuisiaLuinopsis 2 5 7 928.671.428.611筒叶蝶兰属Paraphalae-nopsisPhalpha-laenopsis 1 4 51020.080.020.012盆距兰属GastrochilusGastronopsis 1 3 41125.075.050.013毛舌兰属TrichoglottisTrichonopsis 2 0 212100.0050.014安格兰属AngraecumAngraeconop-sis 2 0 212100.00100.015闭距兰属CleisocentronCleisonopsis 2 0 212100.000.016钻柱兰属PelatantheriaPelathanopsis 2 0 212100.000.017阿梅兰属AmesiellaAmenopsis 0 1 1130100.00.018蛇舌兰属DiploproraDiplonopsis 1 0 113100.00100.019漏斗兰属EurychoneEurynopsis 1 0 113100.00100.0合计Total34320354662.837.2
注:数据来源于英国皇家园艺学会(RHS)网站,数据统计至2020-12-31。
Note: Data are adapted from the Royal Horticultural Society website on December 31, 2020.
对19个蝴蝶兰近缘属作为亲本在英国皇家园艺学会上登录的杂交组合数每10年进行统计,结果见图1。19个近缘属的杂交组合数在1931—1960年很低,共登录了17个杂交组合,主要是万代兰属、火焰兰属、槽舌兰属、蜘蛛兰属和凤蝶兰属,平均每10年登录杂交组合数为5.7个;但1961年以后,杂交组合数迅速增加,平均每10年登录杂交组合数为88.2个,是1931—1960年每10年均值的15.5倍。主要是因为上世纪60年代至今,兰花无菌播种技术的发展与应用促进了兰花杂交育种的迅速发展,杂交育种已成为新品种培育的主要手段。就单个近缘属而言,自1961年以来,万代兰属和火焰兰属在每个时期杂交组合数均保持在前2名;钻喙兰属在1931—2010年的80年间登录杂交组合数为20个,但2011—2020年的10年间登录杂交组合数为21个,主要是因为大部分的钻喙兰属有香味,现代人对香味的追求更执着;槽舌兰属在2000年之前只登录了1个,但2001—2020年增长较快,共登录14个杂交组合。
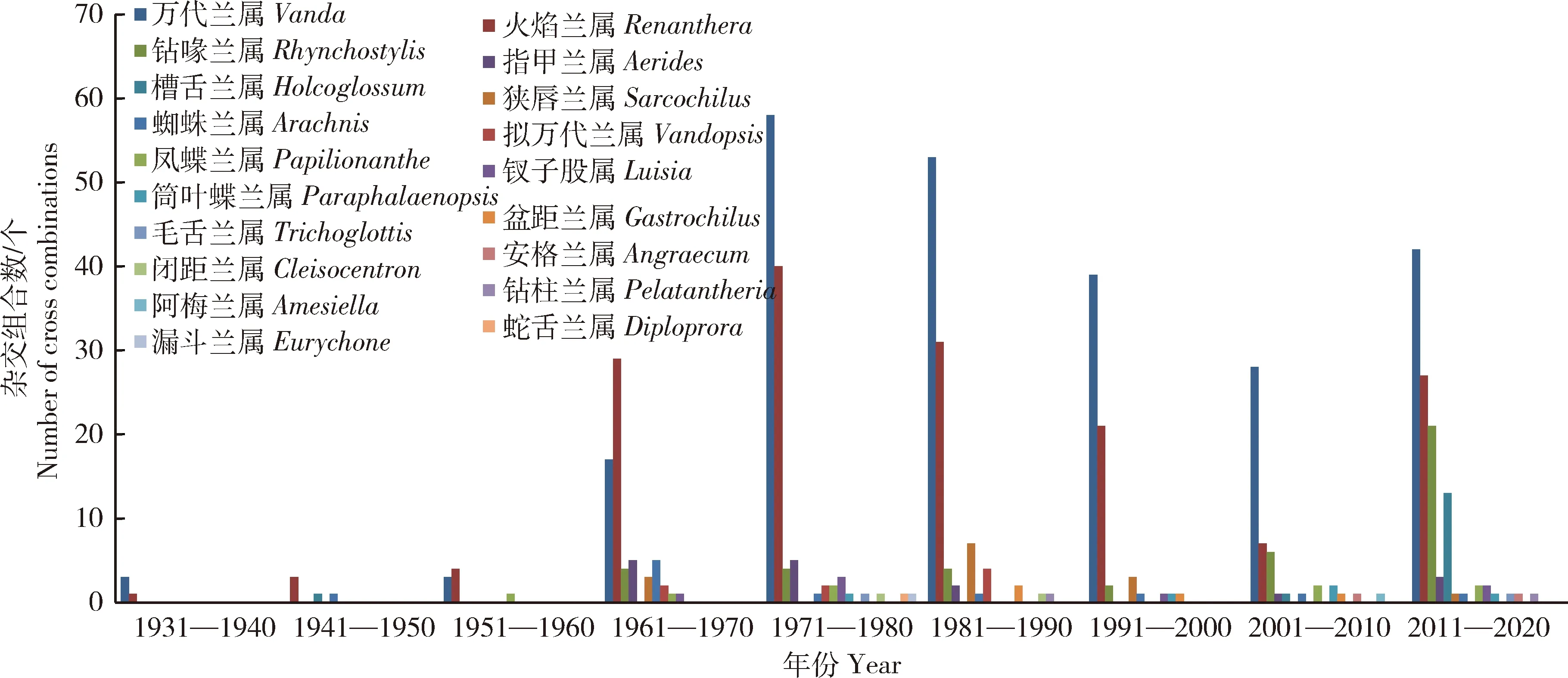
图1 蝴蝶兰19个近缘属为亲本在英国皇家园艺学会上每10年登录的杂交组合数Fig.1 Number of cross combinations registered in RHS with 19 closely related genus of Phalaenopsis in every 10 years
2.2 蝴蝶兰属间杂交差异及效率分析
蝴蝶兰与RHS上登录的19个近缘属杂交成功,但每个近缘属作为父本或母本时与蝴蝶兰属间杂交成功的组合数存在差异(表2)。万代兰属、火焰兰属、指甲兰属、槽舌兰属、狭唇兰属、蜘蛛兰属、毛舌兰属、安格兰属、闭距兰属、钻柱兰属、蛇舌兰属和漏斗兰属这12个近缘属作母本时,其杂交组合数均大于其作父本的杂交组合数;而钻喙兰属、钗子股属、筒叶蝶兰属、盆距兰属和阿梅兰属这5个近缘属作父本时,其杂交组合数均大于其作母本的杂交组合数;但拟万代兰属和凤蝶兰属等2个近缘属作母本或父本,杂交成功率差别不大。同时,以蝴蝶兰杂交种为亲本与排名前3的万代兰属、火焰兰属和钻喙兰属杂交时登录的组合数分别为134、125和27个,占各自登录数的55.1%、76.7%和65.9%(表2)。
在属间杂交效率方面:萼脊兰属和风兰属与蝴蝶兰之间的属间杂交共进行了160个杂交组合,只有12个杂交组合形成的果荚经无菌播种后获得528株杂种,杂交组合成功率为7.50%;蝴蝶兰与火焰兰的远缘杂交过程中共进行了48个杂交组合,其中6个杂交组合的果荚经无菌播种后成功获得441株杂交后代,杂交组合成功率为12.50%;利用钻喙兰(Rhy
.retusa
)与蝴蝶兰进行正反远缘杂交,杂交组合共计36个,其中1个杂交组合成功共获得远缘杂种141株,杂交组合成功率为2.78%;吴容仪等以蝴蝶兰和万代兰进行属间杂交,将万代兰原生种中的橘色和蓝色导入蝴蝶兰中,获得6个橘色杂交组合和1个蓝色杂交组合,并利用蝴蝶兰与海南钻喙兰(Rhy
.gigantea
)进行属间杂交,成功获得有香味的杂交组合5个。以上研究表明蝴蝶兰与其近缘属杂交,虽然杂交组合成功率不高,但可以获得成功,而与墨兰(Cymbidium
sinense
)和碧玉兰(Cym
.lowianum
)等非蝴蝶兰近缘属杂交,则难以成功。3 蝴蝶兰与近缘属杂交存在的问题
从蝴蝶兰与19个近缘属杂交成功组合数的差异可以看出,大部分正反杂交可以成功,但主要以蝴蝶兰杂交种为主要亲本;同时从研究报道可知,蝴蝶兰与近缘属的杂交效率不高,属间杂交成功率在2.78%~12.50%。由于现代蝴蝶兰杂交种遗传背景较复杂,在蝴蝶兰属间、种间和种内杂交中经常产生有不可育种子的情况,表现为杂交不亲和性障碍。
在大多数被子植物中,胚珠和胚囊在授粉时已经完全发育成熟,受精通常发生在授粉后几个小时。而兰花的胚珠发育依赖于授粉,由于授粉和受精之间的时间间隔较长,花粉管在进入成熟胚囊之前必须在子房中存活一段相当长的时间,如天麻(Gastrodiaelata
)需要4 d,苏维斯万代兰(Vanda
suavis
)需要10个月,而蝴蝶兰需要60~65 d,受精发生在授粉后65~70 d。蝴蝶兰在授粉后花粉粒不萌发,直到3 d后开始萌发,这期间四分体的空泡化和分离可能是花粉粒萌发的前提,接下来花粉管继续生长,并迅速充满整个子房腔,同时子房发育并伸长,直到授粉60~65 d,花粉管才朝着几乎在同一时间发育成熟的胚珠方向生长。这可能是由于成熟胚珠发出的信号引导着顶端花粉管朝着胚珠方向生长,从而完成受精,而成熟胚珠助细胞分泌出的富含半胱氨酸的蛋白质是引导花粉管的关键因子。因此,蝴蝶兰属间杂交过程中,花粉管无法到达胚珠完成受精则表现为受精前障碍,即授粉5~10 d内花朵凋谢或授粉1~2个月内能够座果但后期果荚变黄并脱落,如Kim等发现在萼脊兰、风兰与蝴蝶兰属间杂交中有54%的花朵在授粉后脱落,没有形成果荚;若花粉管到达胚珠并完成受精,但后期胚发育停止或发育不正常则表现为受精后障碍,即有些果荚发育时间能够达到3~4个月或更长时间,但果荚内呈棉絮状且无种子。4 克服蝴蝶兰属间杂交障碍的措施
4.1 保持花粉和柱头活性
喻兰等认为蝴蝶兰花粉块随着开放时间的延长,体积减小、颜色加深、质地变硬且活力减弱,柱头活性在蝴蝶兰开花10~30 d内较强,此时进行人工杂交能获得较高的成功率。Balilashaki等得出蝴蝶兰杂交授粉最适季节是冬季(12月—翌年2月),其次是春季(3—5月),而夏季(6—8月)授粉最不利于结实。目前,蝴蝶兰虽然可以通过花期调控实现全年开花,但从喻兰等和Balilashaki等的研究结果来看,若以蝴蝶兰为母本进行属间杂交,选择冬季和春季且蝴蝶兰开花10~30 d内进行授粉有利于成功。低温或超低温保存可延长花粉寿命,这是花期不遇的植物间进行杂交育种的有效途径。蝴蝶兰花粉在室温4周后失去活力,在4 ℃的环境下能存活40周,在-20 ℃或更低温度下保存96周的花粉授粉后仍能产生种子。因此,无论是花期相遇的蝴蝶兰属间杂交,还是通过花粉贮藏解决花期不遇的蝴蝶兰属间杂交,其花粉活力好和柱头活性强是克服属间杂交不亲和的前提条件。
4.2 扩大蝴蝶兰亲本杂交范围及重复授粉
大多数的蝴蝶兰品种都是通过几代或十几代的杂交育种获得的,其遗传背景较复杂。因此,通过扩大亲本杂交范围及增加花朵进行重复授粉有利于提高蝴蝶兰属间杂交成功率。在萼脊兰、风兰与蝴蝶兰的属间杂交过程中,共进行了160个杂交组合,包括29个Sedirea
japonica
×Phal
.杂交组合、65个Phal
.×Sed.
japonica
杂交组合、20个Sed.
japonica
×Doritis
杂交组合、37个Doritis
×Sed.
japonica
杂交组合、4个Phal
.×Neo.
falcata
杂交组合和5个Neo.
falcata
×Phal
.杂交组合,每个杂交组合重复授粉2~3朵,结果有12个杂交组合成功座果。在蝴蝶兰与火焰兰和钻喙兰的远缘杂交过程中,共进行了84个杂交组合,每个杂交组合重复授粉5~8朵,有7个组合的种子萌发,获得远缘杂种582株。4.3 正反杂交有利于属间杂交成功
Kim等在萼脊兰与蝴蝶兰的属间杂交过程中,进行了151个正反杂交组合,以蝴蝶兰为母本的正交组合Phal
.(Doritis
)×Sed.
japonica
共102个,以萼脊兰为母本的反交组合Sed.
japonica
×Phal
.(Doritis
)共49个,其中正交和反交各有5个杂交组合获得杂种后代;风兰与蝴蝶兰的属间杂交过程,以蝴蝶兰为母本的正交组合Phal
.×Neo.
falcata
共4个,以风兰为母本的反交组合Neo.
falcata
×Phal
.有5个,其中反交有2个杂交组合获得杂种后代,而正交未获得杂种后代;火焰兰与蝴蝶兰的属间杂交共进行了48个正反杂交组合,其中正交Phal
.×Renanthera
和反交Renanthera
×Phal
.各24个,以蝴蝶兰为母本的正交有2个获得杂种后代,以火焰兰为母本的反交有4个杂交组合获得杂种后代;钻喙兰与蝴蝶兰的属间杂交进行了正交Phal
.×Rhy
.retus
和反交Rhy
.retusa
×Phal
.各18个,以蝴蝶兰为母本的正交有1个获得杂种后代,而以钻喙兰为母本的反交未获得杂交后代。从本研究中19个近缘属与蝴蝶兰正反杂交也可以看出,正反杂交有利于属间杂交成功。4.4 亲本倍性的选择
陈和明等在蝴蝶兰与火焰兰远缘杂交育种中发现,小花型和中花型的倍性一般较小,大花型的倍性一般较大。小花型中华火焰兰、云南火焰兰与小花型蝴蝶兰‘小飞象’、‘天天红’、版纳蝴蝶兰和安曼蝴蝶兰以及中花型蝴蝶兰‘童真’‘九美’‘日本满天红’‘斯卡利特’属间正反杂交,共有20个组合座果,果荚内没有种子且无菌播种后均未得到杂种后代,但与大花型蝴蝶兰‘聚宝’‘翔凤’‘内山姑娘’‘新红龙’属间正反杂交,有10个组合座果,其中6个经无菌播种后获得杂种后代。因此,选择大花型四倍体蝴蝶兰与火焰兰属间杂交较易成功,但对于别的近缘属如万代兰、钻喙兰和槽舌兰等与蝴蝶兰的属间杂交,其亲本倍性的选择还需进一步研究。
4.5 胚拯救
兰花的种子极为细小,呈粉末状,由种胚和种皮构成,不含贮存组织胚乳,缺乏营养物质,在自然条件下很难萌发。因此,采用组织培养无菌播种的方法进行胚拯救是提高兰花远缘杂交成功率的有效手段,但远缘杂交过程中,胚有没有形成、胚发育正常与否、胚发育是否充分和胚龄等均对胚胎拯救有直接的影响。在萼脊兰、风兰与蝴蝶兰属间杂交的研究中发现,26%的杂交组合中,种子持续生长3~4个月,才能形成正常的种胚,经无菌播种后种子可以萌发;蝴蝶兰与火焰兰和钻喙兰属间杂交产生的种子,其发育时间达到100~125 d才能萌发。
5 展 望
虽然蝴蝶兰与近缘属杂交当前在英国皇家园艺学会上登录了546个杂交组合,取得了一些杂交育种上的研究进展,但有关属间杂种鉴定等方面的研究报道并不多见。Kim等认为蝴蝶兰属间杂交表现出更多蝴蝶兰的形态学性状遗传在后代中,即整体植株趋向于蝴蝶兰,但在细胞及分子鉴定上还未有过报道。而真正的远缘杂种往往表现出亲本某些性状的同时,也表现出自身的特异性状,且兰花属间杂交后代的花色、花梗直立和香味等被认为是定性和定量遗传。比如蝴蝶兰与火焰兰属间杂种F代的主要数量性状及花色介于亲本之间,F代叶片革质较硬趋向于亲本火焰兰,叶端钝形、下弯及叶形倒卵圆形与亲本蝴蝶兰接近,但叶端比蝴蝶兰明显下弯;蝴蝶兰与钻喙兰属间F代大部分株系表现出蝴蝶兰的花色性状,而以白色为主的钻喙兰花色在F代中也充分遗传,花梗直立在F代中与蝴蝶兰一致;蝴蝶兰和万代兰属间杂交的F代,6个橘色杂种群体和1个蓝色杂种群体充分遗传了双亲优点;蝴蝶兰与海南钻喙兰属间杂交F代,5个杂种群体有香味,充分遗传了海南钻喙兰有香味的特点。
目前,属间杂交育种已经在新品种选育中发挥重要作用,包括可以提高产量、改良品质、增强适应性和抵抗力等。在开展蝴蝶兰的属间杂交育种过程中,首先应确定属间杂交育种目标,然后综合运用多种方法如扩大杂交亲本范围、筛选出亲合力高的杂交组合及亲本、晴朗天气条件下进行重复授粉和切柱授粉等措施,有效提高属间杂交亲和性,并建立受精后的胚拯救技术体系,利用分子生物学技术揭示属间杂交不亲和的机制,以有效的手段进行蝴蝶兰属间远缘杂交,开辟蝴蝶兰育种新途径。
猜你喜欢
快乐作文(1.2年级)(2021年11期)2021-12-19
三农资讯半月报(2020年13期)2020-07-31
青少年日记·小学生版(2019年1期)2019-09-02
人文杂志(2017年9期)2018-03-10
农民致富之友(2017年10期)2017-06-15
现代农业科技(2016年22期)2017-03-24
江苏农业科学(2016年4期)2016-06-14
小天使·一年级语数英综合(2015年8期)2015-07-06
现代农业科技(2009年17期)2009-03-08
