
重度冷应激下民猪背最长肌应答分子调控网络分析
2022-09-23 04:06马守正李忠秋王文涛张冬杰
中国农业大学学报 2022年9期
马守正 汪 亮 李忠秋 王文涛 刘 娣, 张冬杰*
(1.东北农业大学 动物科学技术学院,哈尔滨 150038; 2.黑龙江省农业科学院 畜牧研究所,哈尔滨 150086)
猪是对环境温度较为敏感的一种大型家养动物,尤其新生仔猪。现代化集约饲养模式的兴起,虽然提供了一个稳定的环境,但也导致猪对温度的敏感性提高。冷应激会对机体的神经、心血管和免疫系统造成损害,因此突降的灾害性天气会导致巨大的经济损失。在人和小鼠上的研究显示,骨骼肌和棕色脂肪组织是机体遭受冷应激后的主要产热部位。骨骼肌在室温环境下,它的产热占比在20%左右,但在寒冷环境下,它的产热占比可升到40%。它同时以战栗性产热和非战栗性产热两种方式为机体提供热量,这对棕色脂肪较少的大型哺乳动物或没有棕色脂肪的鸟类和家猪格外重要。骨骼肌的战栗性产热主要是通过骨骼肌不随意的节律性收缩,增加代谢率和产热量。骨骼肌的非战栗性产热独立于战栗性产热,目前已知线粒体质子泄漏和肌浆网钙循环是其主要产热机理。
1954年,Sellers等首次提出在寒冷环境下,小鼠的骨骼肌会通过电活动(Electrical acticity)产热维持体温。Davis在对狗的骨骼肌研究中发现,低温环境会导致骨骼肌的耗氧量增加,切除肌肉内的神经不会影响肌肉产热,提出肌肉的非战栗性产热可能是体液介导的。1997年,de Mei等研究发现,肌质网膜上的Ca-ATP酶(Sarcoplasmic reticulum Ca-ATPase,SERCA
)可介导Ca由细胞基质中泵入肌质网中储存起来,使肌质网内Ca浓度比胞质中高出许多。当机体受到神经冲动或环境变化刺激后,Ca由浓度高的肌质网腔中释放入胞质中,在这一过程中,部分Ca与ATP的合成相耦合,还有一部分Ca泄露,偏离ATP合成,并导致热量散失,被认为是肌肉产热的主要来源。其中起关键作用的SERCA酶受小分子蛋白受磷蛋白(Phospholamban,PLB)和肌质蛋白(Sarcolipin,SLN)调控,尤其是SLN
。2017年Bal等发现在轻度和重度冷适应期间,肌肉的非战栗性产热与棕色脂肪的代谢产热同时被启动,如果一条途径被阻断,另一条途径会增加产热以维持核心温度,2个途径彼此间存在相互作用。此外,肌质网上的线粒体串扰、线粒体生物合成的增加、解偶联蛋白3(Uncoupling protein 3,UCP3)诱导产热以及糖异生的关键酶-果糖-1,6-二磷酸酶(Fructose 1,6-bisphosphatase 2,FBP2)的变化等都会改变骨骼肌的非战栗性产热。由此可知,低温诱导下的骨骼肌非战栗性产热机理非常复杂。但受实验条件所限,目前对大型家养动物的相关研究报道很少。猪既是一种大型的模式动物,同时也是一种重要的经济动物。研究低温环境对猪骨骼肌产热的影响,一方面可为人类的相关研究提供借鉴,另一方面也可为生猪健康养殖提供理论帮助。本研究所选择的民猪是东北地区唯一一个受到国家级保护的地方猪种,因长期生活在较为寒冷的环境中,使其具有明显的耐寒特性。在-30 ℃的低温环境中,也不会出现任何冻伤。本研究通过对遭受重度冷应激后民猪背最长肌进行转录组测序,从全基因组水平上筛选和分析了受低温诱导的基因及lncRNA的表达情况,旨在为猪的骨骼肌非战栗性产热研究提供理论依据。
1 材料与方法
1.1 试验动物
本研究在黑龙江省农业科学院畜牧研究所民猪养殖场进行。2021年1月3日,将6头6月龄体重相近的雌性民猪随机分成2组,每组3头猪。试验开始前,6头民猪全部在室内有供暖的猪舍内饲养,温度控制在(18±2) ℃,试验开始后,一组置于室外的半敞篷舍内饲养,温度随外界环境温度变化而变化,白天气温平均-17 ℃,夜间气温平均-26 ℃,一组仍留在温暖的舍内饲养,两组均保证自由采食和饮水,共处理3 d。试验结束后,屠宰6头民猪,取100 mg背最长肌置于液氮中保存。
1.2 RNA的提取
采用酚/氯仿法提取RNA,使用NanoDrop 2000&8000微量分光光度计对所提取的RNA进行纯度检测,使用安捷伦2100 Bioanalyzer进行质量浓度与完整性检测。
1.3 文库构建和上机测序
每个样品取1 μg总RNA作为起始量构建lncRNA文库。首先使用Ribo-off rRNA Depletion试剂盒去除rRNA,向反应体系中加入 Fragmentation Buffer使RNA片段化成为短片段,再以片段后的RNA为模板,利用随机引物进行逆转录,实现cDNA第一链合成,并加入2nd Strand Marking Buffer、2nd Strand/End Repair Enzyme Mix合成cDNA第二链,后经末端修复、加碱基A,加测序接头,通过磁珠筛选回收目的片段,加UNG酶消化cDNA二链,并进行PCR扩增,最后通过磁珠纯化回收目的片段,从而完成整个文库制备工作。使用NovaSeq 6000 S4对构建好的文库进行测序。
1.4 原始数据的处理
测序得到的原始下机序列,含有测序接头序列以及低质量序列,为了保证信息分析数据的质量,需要对原始下机数据序列进行过滤,包括去除接头污染的Reads,低质量的Reads,含N比例大于5%的Reads,以及与核糖体RNA匹配的Reads,得到高质量的Clean reads后再进行后续分析。
1.5 lncRNA的鉴定
lncRNA的筛选条件如下:转录本长度≥200 bp,外显子个数≥2;计算每条转录本的reads覆盖度,筛去所有样本中均小于5的转录本;利用gffcompare(http:∥ccb.jhu.edu/software/stringtie/gff.shtml)同猪的注释文件进行比较,筛除猪中已知的mRNA及其他非编码RNA(rRNA,tRNA,snoRNA,snRNA等);根据比较结果中的class_code信息(“u”,“i”,“x”)筛选潜在的lincRNA、intronic lncRNA和anti-sense lncRNA。将筛选所得的lncRNA作为最终的候选lncRNA进行后续分析。综合4种分析软件进行是否具有编码潜能的筛选,主要有CNCI分析、CPC分析、PFAM蛋白结构域分析和CPAT分析。这4种分析方法均判别为non-coding的转录本为最终的lncRNA数据集。
1.6 民猪背最长肌受冷应激诱导的基因表达量分析
使用FPKM定量估计基因表达值,采用DEseq2进行试验组和对照组间差异表达基因分析,满足|logFC|≥1和q
<0.05的差异基因被认为达到显著水平。根据两组间上下调基因情况绘制差异表达基因的火山图。1.7 民猪背最长肌受冷应激诱导的基因功能分析
GO富集分析。针对GO数据库中的二级条目,计算每个条目的基因数目,应用超几何检验,找出与整个基因组背景相比,差异表达基因显著富集的GO条目。对KEGG中每个Pathway应用超几何检验进行富集分析,找出差异表达基因显著性富集的Pathway。
1.8 民猪背最长肌受冷应激诱导的lncRNA与mRNA的联合分析
对于差异表达的lncRNA的靶基因,分别进行Cis和Trans靶标分析,通过靶基因间接预测其功能。Cis作用靶基因预测主要是通过lncRNA的功能与其在基因组座位上临近的蛋白编码基因的相关性进行的。将lncRNA相邻位置(上下游50 kb)的蛋白编码基因筛选出来作为靶基因。Trans靶基因预测主要是通过lncRNA的功能不依赖于和编码基因的位置关系,而是与共表达的基因相关性进行的。根据lncRNA同mRNA的表达量相关性系数(斯皮尔曼相关系数,corr≥0.9)进行筛选。筛选后获得的靶基因开展同mRNA相似的功能分析。根据识别出的差异表达lncRNA基因与mRNA基因及lncRNA的顺式、反式靶标预测的基因的关系,绘制差异lncRNA与靶基因的调控网络分析。
1.9 民猪背最长肌差异表达基因的Real-time PCR验证
利用Roche实时荧光定量PCR仪LightCycler480Ⅱ对处理组存在显著差异的高表达8个基因和2个lncRNA进行SYBR Green实时定量检测,引物信息见表1。反应体系为:cDNA样品0.5 μL,2×SYBR Green PCR Mixture 10 μL,特异性引物上下游各0.5 μL,灭菌水补充至20 μL。反应程序为:95 ℃ 10 min;95 ℃ 15 s,60 ℃ 30 s,72 ℃ 30 s,40个循环。检测结果根据2法计算各模板中目的基因相对于内参基因β
-actin
的表达量。2 结果与分析
2.1 民猪遭受冷应激后背最长肌转录组测序结果
原始下机数据经过滤处理后,每个样品平均获得98 M的Clean reads,与基因组的比对效率为96.12%,其中,38.94%的序列比对到外显子区域,30.50%的序列比对到内含子区域,30.56%的序列比对到基因间区, 筛选出的lncRNA经CNCI分析、CPC分析、PFAM蛋白结构域分析和CPAT分析后,取四者间的交集,共获得10 220个lncRNA。
表1 qRT-PCR检测用引物信息
Table 1 Primer information for qRT-PCR
基因或长链非编码RNAGene or lncRNA引物序列(5'-3')Primer sequence (5'-3')SRXN11F: TCCACTCGGGTTGTATCGC1R: TGTCCACGAGGCTCTGCACSF12F: TTGCCTTTGAGTTTGTAGACC2R: TTGGCATTGGGAGTGTTGGPRC5A3F: GTGGATTGCCCTGCTCTT3R: ATGGTGTTCCGTTGCTTGIL1R14F: CGGGTCCACCTCTAACTC4R: GTAGTCGTCCCTGCCACACD2095F: TCTTCGTCTCATTGGGTTTC5R: TGGGTCTCCTGCTGGTCTARID5B7F: GGGCAACCCAGGTATCAT7R: TCATCCCTCGCAATCAGTMMP198F: CCCCAAGAGGCTGAATAG8R: GGTAGCTGCTGAAGTCCATITGA59F: CTCGCCAGGCTAGTTCCA9R: AAGGATGGTGACATAACCGTAGlncRNA13646010F: AGGAGGGAACAGATGGATTG10R: GTGAGGGCTGGCTTGTATTTlncRNA7596311F: TAGACAGAGGCTCCCTTCAT11R: CTGGGTTGCACAACTTCCβ-actin12F: CGGGACCTGACCGACTACCT12R: GGGCCGTGA TCTCCTTCTG
2.2 民猪遭受冷应激后背最长肌差异表达基因的筛选与分析
按照|logFC|≥1和q
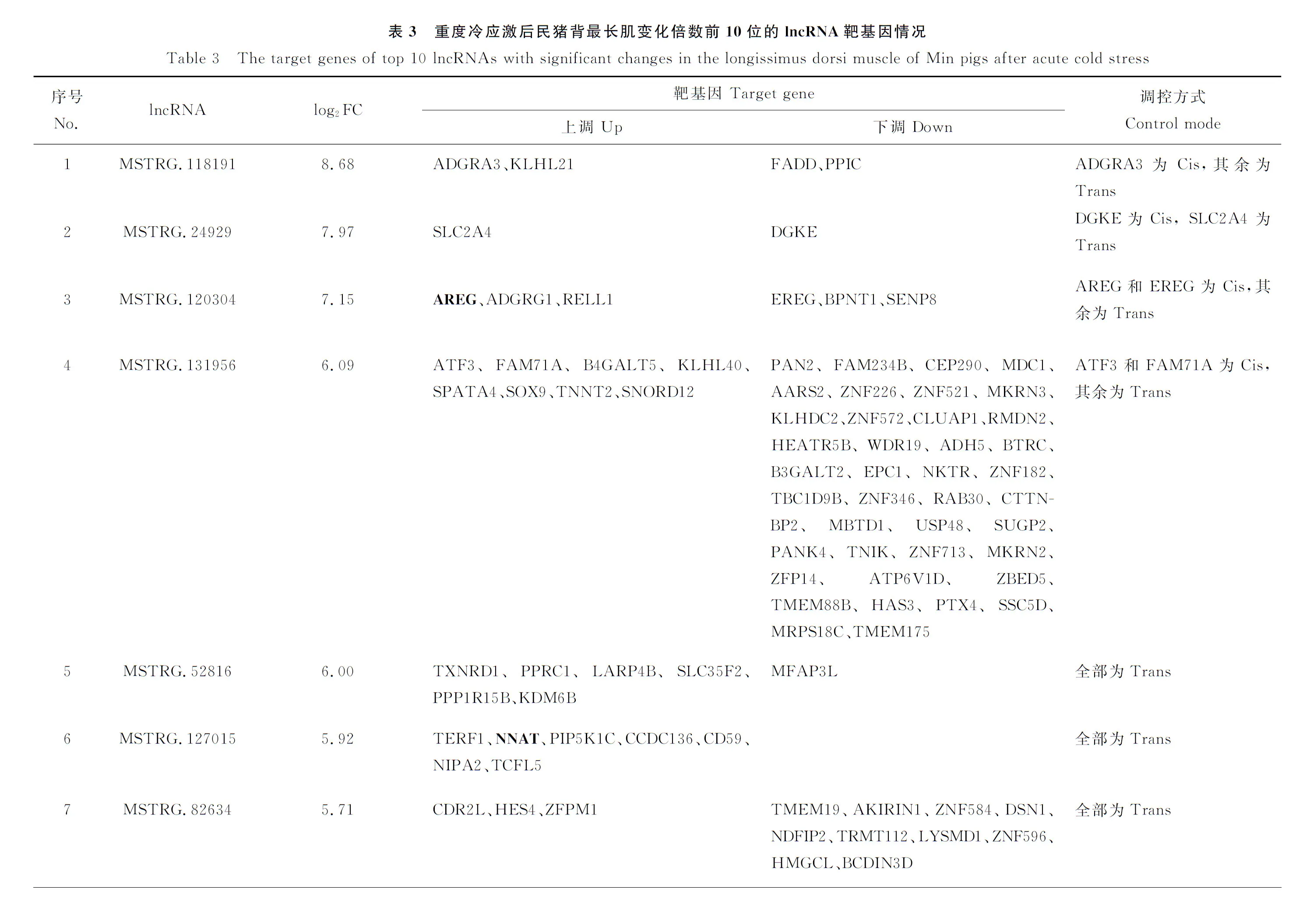
<0.05的条件筛选后,民猪在遭受急性重度冷应激后,背最长肌共有88个基因表达发生了显著变化,其中53个基因表达发生了显著上调,35个基因表达发生了显著下调。91个lncRNA表达发生了显著变化,其中53个lncRNA表达发生了显著上调,38个lncRNA表达发生了显著下调。由于猪的lncRNA数据库还不完善,因此这91个lncRNA均显示为novel lncRNA。表达发生显著变化的前10个基因列表见表2。上调变化倍数最大的基因是肝细胞生长因子激活因子(Hepatocyte growth factor activator,HGFAC
),上调了9.55倍;下调变化倍数最大的基因是多亮氨酸重复区和跨膜域1(Leucine rich rRepeats and transmembrane domains 1,LRTM1
),下调了7.04倍。2.3 民猪遭受冷应激后背最长肌差异表达基因的功能分析
对差异表达的基因进行GO功能注释,发现在分子功能(Molecular function)中不存在显著富集的条目,在生物过程(Biological process)中,有21个条目存在显著富集,其中富集程度最高的是胶原分解代谢过程(Collagen catabolic process),在细胞组分(Cellular component)中,有5个条目存在显著富集,其中4个是位于细胞外区间(图1)。
表2 重度冷应激后民猪背最长肌表达发生显著变化的前10个基因
Table 2 The top 10 genes with significant changes in the longissimus dorsi muscle of Min pigs after acute cold stress
基因Genelog2差异倍数log2 Fold change变化Change基因Genelog2差异倍数log2 Fold change变化ChangeHGFAC9.55UpLRTM1-7.04DownTREH5.59UpKCTD4-3.06DownFOSL14.87UpPCSK1N-2.82DownAREG4.53UpTMEM139-2.80DownMMP254.52UpCSRNP3-2.71DownHAS14.26UpLRRN3-2.63DownSLC11A13.72UpNCBP2-AS2-2.49DownTRIB33.67UpCOL8A2-2.41DownSLCO4A13.47UpENHO-2.25DownPRSS363.44UpNPPC-1.91Down
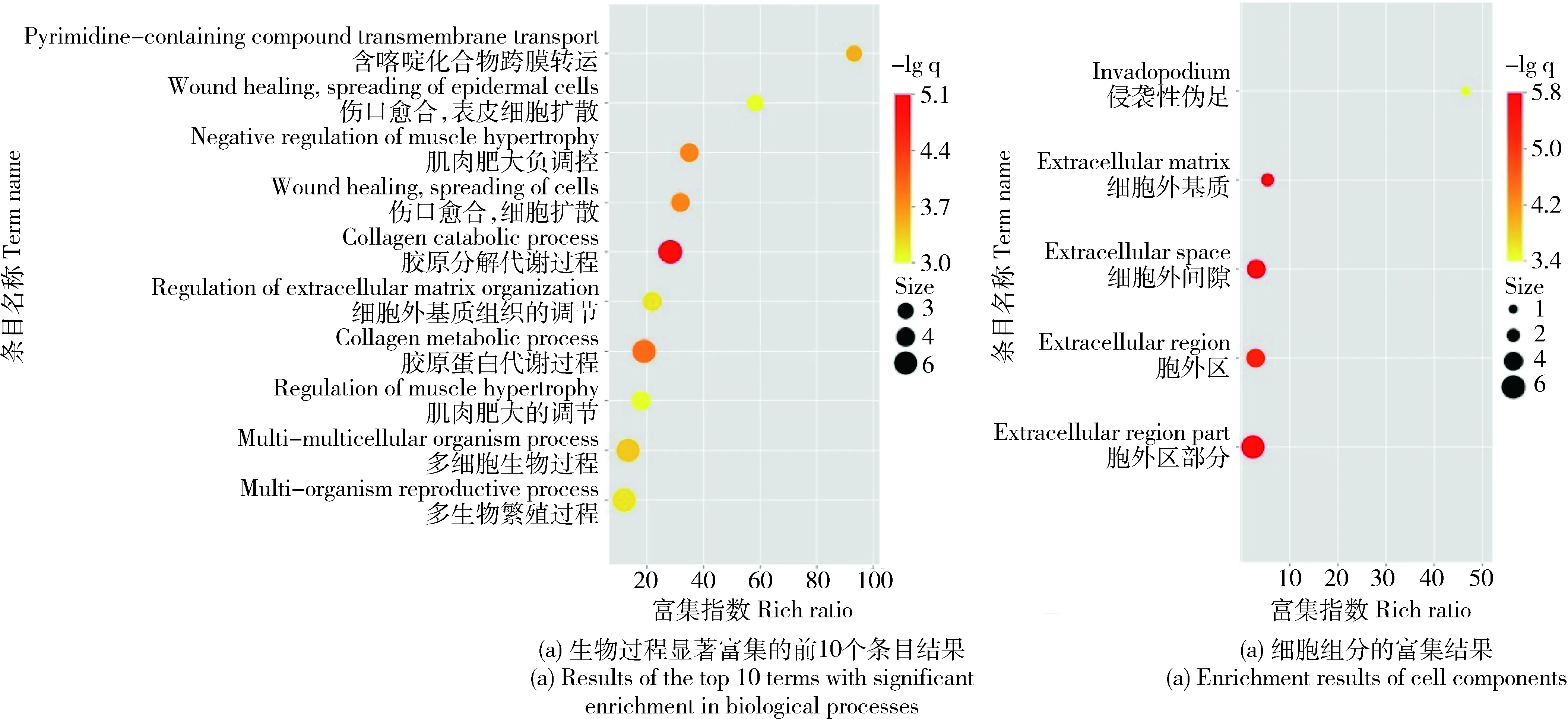
每个点的颜色表示该GO条目的富集程度。每个点的大小表示富集到该GO条目的基因的个数。用于表示富集程度的rich ratio,其计算公式为:(该通路的差异基因/所有的差异基因)/(注释到该通路的基因/所有能被注释到的基因)。 The abscissa represents the degree of enrichment, and the ordinate represents the GO term. The color of each dot indicates the degree of enrichment of that GO term. The size of each point represents the number of genes enriched for that GO term. The rich ratio used to express the degree of enrichment is calculated as (differential genes of this pathway/all differential genes)/(genes annotated to this pathway/all genes that can be annotated).图1 重度冷应激下民猪背最长肌差异表达基因GO条目FDR值富集图Fig.1 Enrichment map of Go terms FDR value of differentially expressed genes in longissimus dorsi of Min pig under acute cold stress
对差异表达的基因进行pathway富集分析,发现仅丝裂原活化蛋白激酶(MAPK)信号通路存在显著富集,该通路中7个基因白细胞介素1受体1型(Interleukin 1 receptor type 1,IL1R1
)、双调蛋白(Amphiregulin,AREG
)、丝裂原活化蛋白激酶8(Mitogen-activated protein Kinase 8,MAP3K8
)、70 ku热休克蛋白1L(Heat Shock Protein Family A (Hsp70) Member 1 like,HSPA1L
)、集落刺激因子1(Colony stimulating factor 1,CSF1
)、RELB原癌基因NF-κβ亚基(RELB proto-oncogene, NF-kB subunit,RELB
)的表达发生了显著上调,2个基因成纤维细胞生长因子受体4(Fibroblast growth factor receptor 4,FGFR4
)和p21活化激酶1(p21- activated Kinase 1,PAK1
)的表达发生了显著下调。2.4 民猪遭受冷应激后背最长肌差异表达lncRNA的靶基因预测
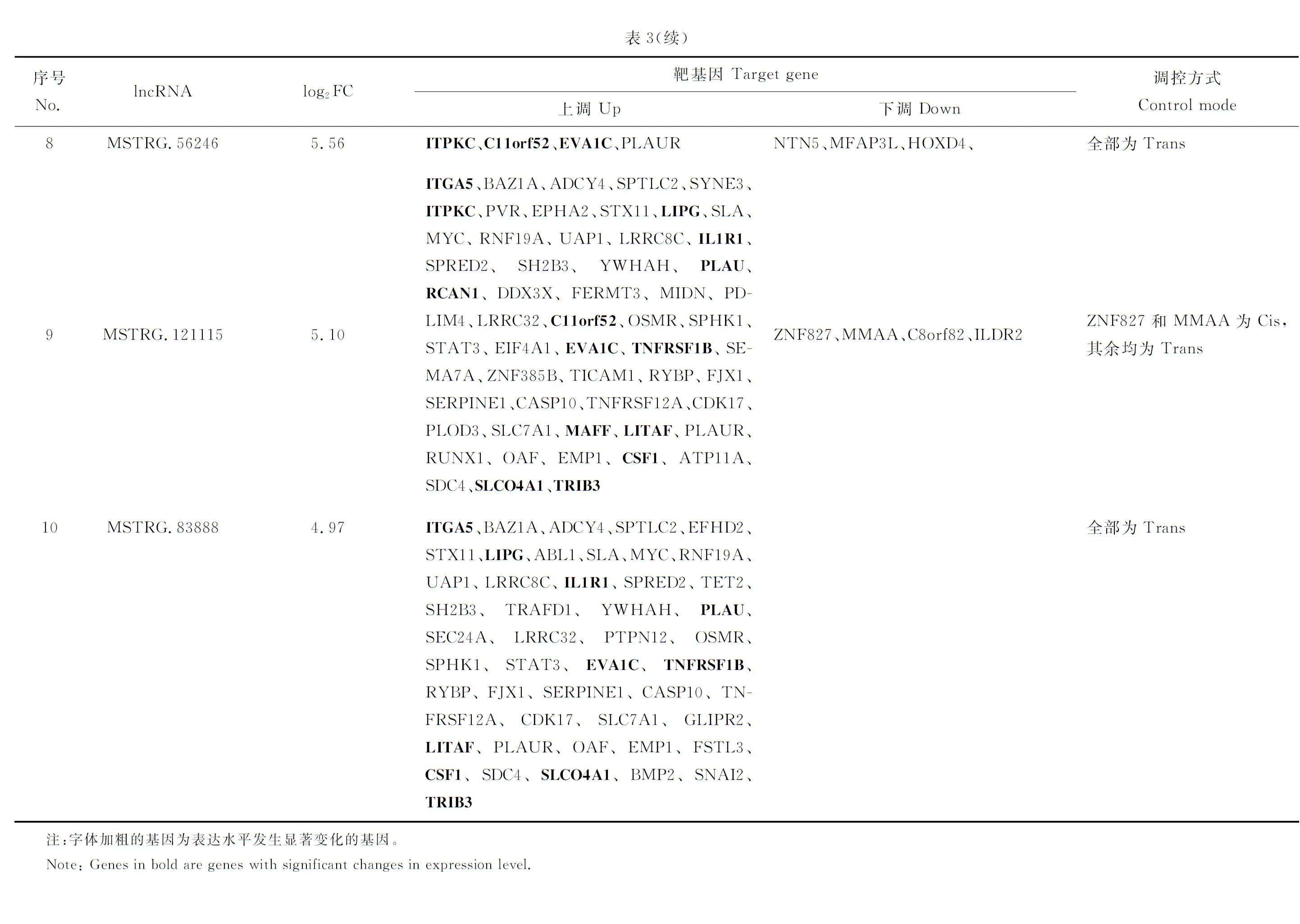
lncRNA不编码蛋白,主要通过顺式(Cis)或反式(Trans)方式作用于蛋白编码基因来实现。本研究中表达发生显著变化的91个lncRNA顺式调控129个靶基因,反式调控750个靶基因。通过构建lncRNA与mRNA之间的相互作用网络(仅列出了发生显著变化的前10个),发现两者间的调控关系错综复杂(图2)。比如lncRNA122198同时调控39个基因,使它们的转录发生了向上或向下的变化,而基因C11orf52
则同时受到9个lncRNA的调控。表3列出了表达发生显著变化的前10个lncRNA及其靶基因表达变化和调控模式。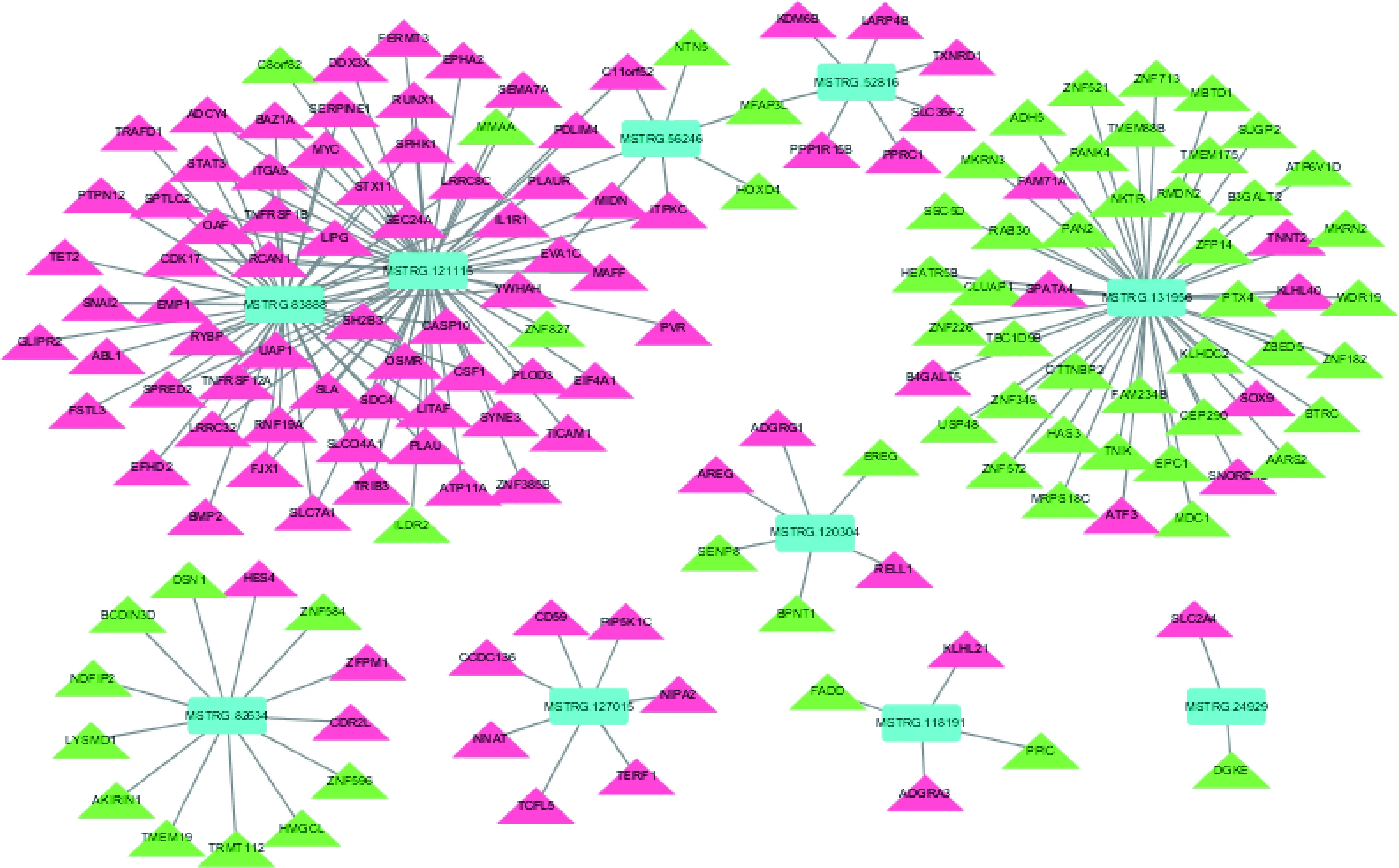
蓝色代表lncRNA,红色代表上调表达的基因,绿色代表下调表达的基因。 Blue represents lncRNA, red represents up-regulated genes, and green represents down-regulated genes.图2 重度冷应激下民猪背最长肌差异表达lncRNAs和mRNAs之间的调控网络图Fig.2 Regulatory network between differentially expressed lncRNAs and mRNAs in the longissimus dorsi of Min pigs under acute cold stress
2.5 民猪遭受冷应激后背最长肌差异表达lncRNA的靶基因功能分析
对差异表达的lncRNA靶基因进行GO功能注释,发现在分子功能中有19个条目存在显著富集,其中磷酸转移酶活性(Phosphotransferase activity)富集程度最高;在生物过程中,有232个条目存在显著富集,其中富集程度最高的是单核细胞活化(Monocyte activation),在细胞组分中,有20个条目存在显著富集,其中富集程度最高的在核质(Nucleoplasm)和核部分(Nuclear part),每个本体中富集程度前10的条目情况见图3。
对差异表达的lncRNA靶基因进行pathway富集分析,发现MAPK信号通路(MAPK signaling pathway)和TNF信号通路(TNF signaling pathway)存在显著富集,MAPK通路中有21个基因的表达发生了显著上调,6个基因的表达发生了显著下调。TNF通路中,有11个基因的表达发生了显著上调,3个基因的表达发生了显著下调。

每个点的颜色表示该GO条目的富集程度。每个点的大小表示富集到该GO条目的基因的个数。用于表示富集程度的rich ratio,其计算公式为:(该通路的差异基因/所有的差异基因)/(注释到该通路的基因/所有能被注释到的基因)。 The abscissa represents the degree of enrichment, and the ordinate represents the GO term. The color of each dot indicates the degree of enrichment of that GO term. The size of each point represents the number of genes enriched for that GO term. The rich ratio used to express the degree of enrichment is calculated as (differential genes of this pathway/all differential genes)/(genes annotated to this pathway/all genes that can be annotated).图3 重度冷应激下民猪背最长肌差异表达lncRNA的靶基因GO条目FDR值富集图Fig.3 Enrichment map of GO terms FDR value of target genes of differentially expressed lncRNAs in the longissimus dorsi of Min pig under acute cold stress
2.6 测序结果的实时定量PCR验证
为了验证RNA测序结果的准确性,选择了8个显著上调表达的mRNA和2个显著上调表达的lncRNA进行了 qRT-PCR验证。结果显示,qRT-PCR定量检测值与测序值相比,变化趋势基本一致(图4),表明测序结果可信。
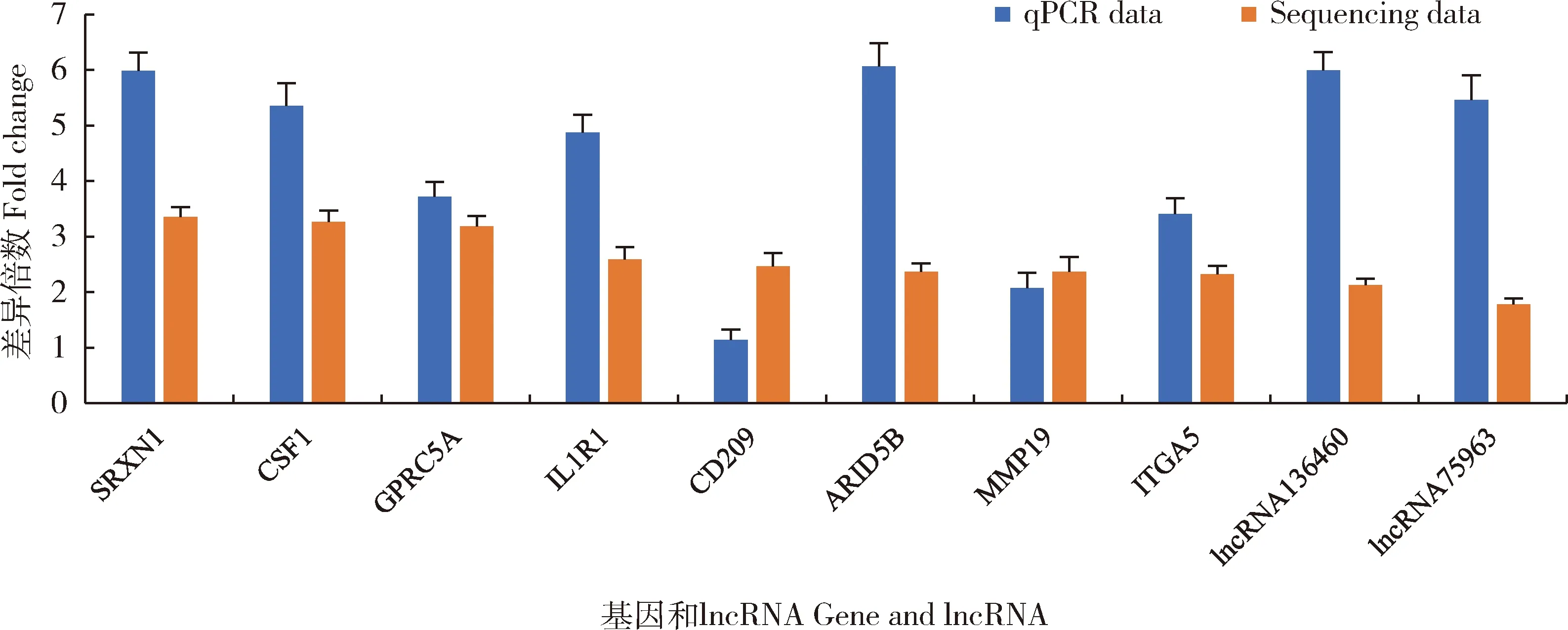
图4 测序结果的qRT-PCR验证Fig.4 qRT-PCR validation of sequencing results
3 讨 论
战栗是肌肉收缩的一个重复过程,是产生热量的一个重要机制,是急性寒冷暴露的第一道防线。各种转基因小鼠模型的研究表明,单靠战栗不足以维持机体核心温度(Core body temperature,Tc),必须激活非战栗机制来维持热量生产。对新生哺乳动物和成年啮齿动物的研究表明,棕色脂肪组织(Brown adipose tissue,BAT)是非战栗性产热(Nonshivering thermogenesis,NST)的重要部位。因此,近二十年来对非战栗性产热的研究大多集中在棕色脂肪上。虽然BAT在许多哺乳动物中发挥重要作用,但它通常仅限于新生儿阶段,在成年大型非冬眠哺乳动物(包括人类)中作用减弱。此外,还有一些恒温动物,如鸟类、有袋类动物和猪,它们可以在没有BAT的情况下依旧保持恒定的Tc,并在寒冷的气候中正常生长。因此,有人提出骨骼肌是非战栗性产热的另一个重要部位。
二代测序技术的出现,使得研究人员可以在全转录组水平利用测序技术同时进行定量与定性分析,提高了分析的效率和准确性。本研究采用该技术对遭受重度冷应激后的民猪背最长肌进行分析,发现88个基因受到显著影响(P
<0.05),其中HGFAC
上调倍数最大,HGFAC是活化单链前体肝细胞生长因子(HGF)成为有生物学功能的HGF的关键酶,在受损组织器官的修复和再生中发挥重要作用,HGFAC-HGF-PPARγ信号通路可参与调控全身的碳水化合物、葡萄糖和脂质代谢。它的上调表达,推测是由于肌肉组织受低温影响发生了损伤。海藻糖酶基因(Trehalase,TREH
)在低温刺激下也发生了5.59倍的上调表达。海藻糖酶是一种催化海藻糖水解的酶,在哺乳动物中鲜有报道,但在昆虫和植物中已经明确其可受低温胁迫影响,该酶在能量代谢、生长和逆境恢复中起着重要作用。其他的,如双调蛋白基因(Amphiregulin,AREG
)、透明质酸合成酶1基因(Hyaluronan synthase 1,HAS1
)、Tribbles同源蛋白3基因(Tribbles pseudokinase 3,TRIB3
)等都发生了显著的上调。AREG是一种细胞因子,参与抵抗由蠕虫引起的感染免疫、免疫调节和伤口修复,是炎症环境下肌成纤维细胞分化的重要驱动因素。HAS1是负责透明质酸合成的三种同工酶之一, 它的表达和活性由白细胞介素和细胞因子等促炎因子诱导,在许多炎症反应中如骨关节炎、感染性肺病中,HAS1的表达都增高,表明它参与了炎症反应。TRIB3属于Tribbles信号调节蛋白家族,是关键的“压力调节开关”,参与内质网应激反应、细胞生长分化等生物过程。此外,与先天性免疫相关的MMP25
和天然抗性相关的SLC11A1
也发生了显著上调。被认为在非战栗性产热中起重要作用的SERCA1a
并未发生显著变化,但其调节因子SLN
的表达水平下降了一半。通过对发生显著上调的基因分析发现,急性重度冷应激下民猪的背最长肌发生了应激和炎症反应。本课题组在前期研究中发现急性重度冷应激会造成民猪皮下脂肪与免疫、呼吸、氧化应激、血液循环和脂类代谢等相关基因的表达水平显著上调,而与神经系统相关基因的表达水平发生显著下调。小鼠骨骼肌中的催产素受体(Oxytocin receptor,Oxtr
)、瞬时感受器电位香草酸受体1 (Transient receptor potential vanilloid 1,TRPV1
)、冷诱导RNA结合蛋白(Cold-induced RNA-binding protein,CIRP
)、锌α2糖蛋白(Zinc-α2-glycoprotein,ZAG
)等基因也会受到冷诱导而上调表达。本研究中筛选出的差异表达基因与前人的研究结果存在较大差异,推测这是由基因表达特性导致的。基因的表达具有显著的时间特异性和空间特异性,受外界环境因素影响。因此种属、日龄、组织、环境温度以及作用时间等因素都会导致筛选出的基因存在差异。通过对差异表达基因的GO功能注释分析和pathway富集分析后发现,这些差异表达基因所涉及的生物过程主要集中在胶原代谢、伤口愈合表皮细胞扩散和肌肉肥大调节过程等。这与人们在大鼠比目鱼肌上发现的冰敷可促进胶原沉积增加、新生雏鸡的急性冷暴露会导致心肌肥大等现象是一致的。差异表达基因在细胞组分(CC)的分析中发现,它们多位于细胞外区间,推测这可能与急性冷暴露下Ca、K等离子的转运变化相关。重度冷刺激下,民猪肌肉组织中仅有1条生物学通路(MAPK信号通路)受到影响,说明低温对民猪肌肉组织的正常生理功能影响较小。MAPK信号通路有三级的信号传递过程,可调节细胞的生长、分化、应激、炎症反应等多种重要的生理/病理效应,是将环境刺激转化为细胞反应的重要信号模块。目前,在植物中已明确其对冷应激的应答反应机制,但在哺乳动物中还未见其对冷应激应答的报道。
lncRNA是长度大于 200 个核苷酸的非编码 RNA,在剂量补偿效应、表观遗传调控、细胞周期调控和细胞分化调控等众多生命活动中发挥重要作用。民猪背最长肌遭受重度冷应激后,91个lncRNA发生了显著变化,因猪的lncRNA数据注释有限,所以这些lncRNA均显示为novel lncRNA,它们主要以反式调控的方式作用于靶基因。在对靶基因的功能注释中发现,许多基因与单核细胞激活作用、钙非依赖性细胞间粘附的调节、蛋白质去甲基化、嘧啶脱氧核糖核苷代谢等生物过程相关。分子功能主要集中在醇基作为受体的磷酸转移酶活性和RNA聚合酶Ⅱ调节区特异性 DNA 结合序列等。靶基因所富集的生物学通路,除了差异表达基因所富集的MAPK通路外,还有肿瘤坏死因子(TNF)信号通路。TNF是哺乳动物免疫反应中的一个关键传递者和调节者,它的主要生化反应是激活并调控NF-KB和AMPK途径,在某些病理状态下, 也能诱发细胞死亡。该通路的激活,也再次证明了冷应激下,骨骼肌内发生了炎症反应。通过以上分析可知,冷应激影响了民猪背最长肌部分基因的表达,同时也影响了调控基因转录的部分lncRNA的表达。
本研究仅从全基因组水平探讨了重度冷应激对民猪骨骼肌基因和lncRNA表达模式的影响,未来应从分子、细胞和个体水平深入研究冷应激对家猪能量代谢及其他方面的影响,通过培育耐寒品种、改善饲料营养等多种方式提高家猪抗寒性。
4 结 论
民猪在遭受3 d的重度冷应激后,背最长肌的88个基因受到显著影响。与炎症反应、低温胁迫、应激反应和营养物质代谢相关的基因发生了显著上调。差异表达基因所涉及的生物过程主要集中在胶原代谢、伤口愈合表皮细胞扩散和肌肉肥大调节过程等,且多位于细胞外区间。显著富集的生物学通路仅MAPK通路1个。91个lncRNA的表达发生了显著变化,这些lncRNA调控879个基因,且以反式调控为主。lncRNA调控的靶基因显著富集到MAPK和TNF通路。
猜你喜欢
中国现代医生(2022年19期)2022-11-04
中国现代医生(2022年19期)2022-08-25
中国典型病例大全(2022年11期)2022-05-13
诗潮(2019年10期)2019-11-19
现代仪器与医疗(2017年1期)2017-03-29
山东体育学院学报(2016年2期)2016-05-25
诗选刊(2016年2期)2016-05-23
荷城文艺(2015年2期)2015-06-10
销售与管理(2006年9期)2006-09-17
青年文摘·上半月(1991年2期)1991-01-01
