
中华蜜蜂ZFP37基因的生物信息学分析及高温下表达特性的研究
2022-09-22 01:49李新宇马卫华杜亚丽姜玉锁
环境昆虫学报 2022年4期
李新宇,马卫华,杜亚丽,徐 凯,姜玉锁
(1.山东轻工职业学院,山东淄博 255300;2.山西农业大学园艺学院,山西太原 030031;3.山西农业大学动物医学学院,山西太谷 030801;4.吉林省养蜂研究所,吉林吉林 132001;5.山西农业大学动物科学学院,山西太谷 030801)
温度是影响昆虫生理状况和生存状态的重要非生物因素。蜜蜂属于变温动物,其个体保持和调节体温的能力不强,外界环境温度的变化会对蜜蜂生理代谢产生影响,如导致蜜蜂的应激反应、改变生理代谢、影响成虫的存活、幼虫的发育等(Kiuchietal.,2008;Bauerfeind and Fischer,2013;Colinetetal.,2018)。当外界环境温度超过蜜蜂所能承受的范围时,蜜蜂的各项生命活动和体内的生理代谢都会受到严重的干扰。
高温可以改变细胞的内环境,对细胞骨架造成损伤,还会影响蛋白质的合成过程(杜尧等,2007)。蜜蜂在长期的进化过程中进化出了一系列措施来应对温度胁迫,如调整自身代谢,合成保护蛋白等,以减缓或者修复高温胁迫对机体造成的伤害。其中最显著的机体响应就是抗逆蛋白的诱导产生,目前来说,在众多抗逆基因中,热激蛋白基因家族发挥着极其重要的作用,它是生物细胞在面临多种应激条件下所产生的一类维持细胞内蛋白质平衡的蛋白(Feder and Hofmann,1999)。热激蛋白具有能够维持细胞骨架稳定、抑制蛋白质变性、恢复变性蛋白原有的空间结构和生物活性的生物功能。目前的研究发现了很多参与昆虫抗逆的基因,其中以热激蛋白基因为主(Huangetal.,2007),其次还包括锌指蛋白、丝氨酸/苏氨酸蛋白激酶等(Krupaetal.,2004;Duanetal.,2008)。
锌指蛋白(Zinc finger proteins,ZFPs)通常是指蛋白结构中含有一段自身折叠形成的手指状结构,且常与Zn2+结合的一类蛋白质。最早的锌指蛋白是在非洲爪蟾卵母细胞中发现的(Frankel and Pabo,1988),之后,许多其他锌指蛋白又被陆续鉴定出来。锌指蛋白在真核生物中广泛分布,通常由半胱氨酸和组氨酸组成,通过与Zn2+结合来维持结构的稳定性;其结构非常多样,其中绝大多数锌指蛋白为类C2H2锌指、高音谱号锌指、钢卷尺状锌指(赵楠等,2009)。锌指蛋白具有许多不同的功能,包括转录的调节、细胞凋亡的调控、蛋白质折叠和组装以及脂质结合等。随着对锌指结构研究的深入,许多具有新型拓扑结构的锌指蛋白逐渐被人们认识到,这些发现可帮助进一步了解锌指蛋白的生物学功能。例如,通过对经典的C2H2结构的锌指蛋白的深入研究使人们对其与DNA结合的机制有了新的认识(Johnetal.,2001)。锌指蛋白作为一类转录因子,不仅可以调控基因的表达和细胞的分化,还在动植物抗逆方面发挥重要作用。锌指蛋白能在转录水平上调控,是由于它能与生物大分子结合,或者通过锌指蛋白间的结合,调控基因转录。Zn2+是影响锌指蛋白功能的关键因素,可以通过锌指结构,使激活子蛋白与增强子序列特异性结合而调节基因的表达。锌指蛋白参与生物体应对多种生物或非生物因素胁迫的抗逆机制(Girietal.,2011),在机体应对高温胁迫时,多个锌指蛋白基因在其转录后调控机制中发挥着重要的作用(Drolletal.,2013)。
ZFP37是C2H2型锌指蛋白家族中的一员,被认为在转录调控中发挥重要作用。有研究显示,ZFP37主要在成年小鼠睾丸和大脑中表达,这表明其可能在控制小鼠精子生成中发挥潜在的作用(Payenetal.,1998)。同样的,ZFP37也被发现参与调控猪子宫内膜上皮细胞的凋亡(Zhangetal.,2019)。现阶段,对于ZFP37的生物功能的研究多集中在小鼠和哺乳动物的生殖代谢上,在昆虫中的作用鲜见报道。
中华蜜蜂Apisceranacerana是我国本土蜂种,其生活习性适应了我国多变的自然环境,对于维持我国生态平衡起着重要作用(Zhangetal.,2014),相比其他常见蜂种,中华蜜蜂具有抗逆性和抗螨能力强等优势(曾志将等,2002;Yang,2005)。本试验根据之前的研究结果,选取中华蜜蜂在高温胁迫下响应较强的一个基因ZFP37,对其蛋白结构进行预测分析,研究胁迫温度和胁迫时间对其表达的影响,以探究中华蜜蜂在遭受热胁迫时ZFP37可能发挥的生理功能,为蜜蜂抗热机制的研究提供理论基础。
1 材料与方法
1.1 供试昆虫
试验所需中华蜜蜂由山西农业大学动物科技学院实验蜂场提供,选取健康无病蜂群3群。每群各选取1~2张老熟的封盖子脾,置于34℃±0.5℃和RH 75%~80%条件下孵育。待幼蜂大量出房后,收集出房幼蜂并标记(N=500/群),将标记的幼蜂放回原蜂群饲养,采集20日龄的标记蜜蜂作为试验样品。
1.2 温度胁迫
从每群随机选取300头标记的20日龄工蜂,将蜜蜂随机分为若干组,置于15 mL离心管中,每管放置1头蜜蜂,离心管上扎有小孔,确保蜜蜂处于设置的温湿度条件下,之后将试验蜜蜂放入恒温恒湿箱中,湿度设置为RH 30%,温度分别为25℃、30℃、35℃、40℃和45℃,胁迫2 h。高温不同胁迫时间的处理为:在45℃,RH 30%条件下分别胁迫处理0.5 h、1 h、1.5 h和2 h,其中0 h 处理组的蜜蜂直接采自蜂箱中的标记蜜蜂。处理完成后,随机选择5头蜜蜂作为1个生物学重复用于提取RNA,每个处理组包括3个生物学重复,立即将蜜蜂投入液氮中,于-80℃下保存备用。
1.3 主要试剂与仪器
Trizol试剂盒(Ambion),PrimeScriptTM RT-PCR reagent Kit试剂盒(TaKaRa),SYBR Premix Ex Taq TM 试剂盒(TaKaRa),HWS0358恒温恒湿箱(江南仪器厂),Veriti©96 well梯度PCR仪(ABI),7500荧光定量PCR仪(ABI),ND-1000核酸蛋白测定仪(Nanodrop),紫外凝胶成像仪(BIO-RAD),DYCP-31DN琼脂糖水平电泳仪,5810R高速冷冻离心机(Eppendorf)。
1.4 RNA的提取和cDNA第一链的合成
使用Trizol试剂盒提取不同样品总RNA,通过1.5%琼脂糖凝胶电泳分析提取的RNA质量,最后将提取的RNA反转录为cDNA。
1.5 引物设计
通过东方蜜蜂全基因组测序数据(NC_014295.1),查找中华蜜蜂ZFP37基因序列,根据所得序列的CDS区使用Primer 3软件设计RT-qPCR引物。
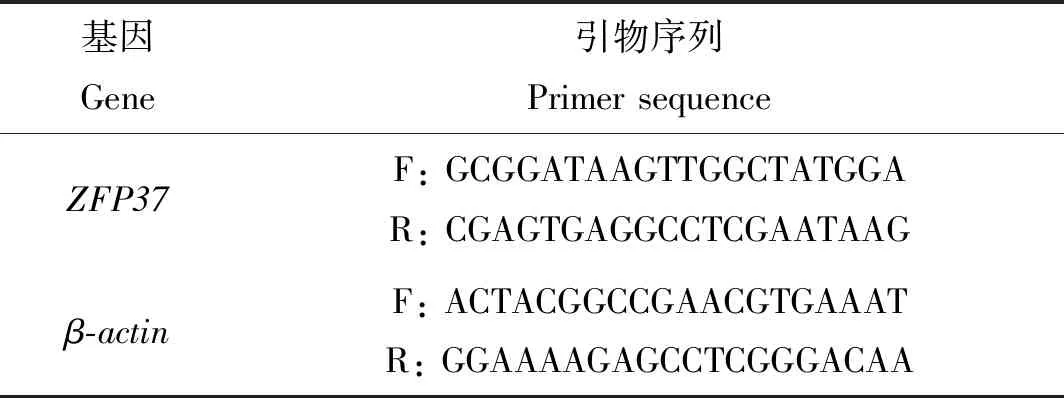
表1 引物序列Table 1 Primer sequence
1.6 RT-qPCR反应
将cDNA模板稀释备用,使用SYBR Premix Ex TaqTMⅡ试剂盒进行RT-qPCR反应。反应体系总体积为10 μL,其中包括5 μL SYBR Premix Ex TaqTMII,1 μL cDNA,0.2 μL ROX Reference Dye II (50×),上、下游引物各0.4 μL,无菌水 3 μL。qRT-PCR热循环程序如下:95℃预变性30 s,95℃变性5 s,60℃退火及延伸34 s,45个循环。用β-actin作为内标基因,每个组织样本重复测定3次。
1.7 序列分析
通过多种生物信息学软件对中华蜜蜂ZFP37的序列进行预测分析,所用各软件如表2。

表2 生物信息学分析所用软件Table 2 Software for bioinformatics analysis
1.8 数据统计与分析
对于qRT-PCR试验结果,根据标准曲线以及荧光曲线的Ct值,采用2-ΔΔCT法进行数据分析。运用SPSS 21软件中的单因素ANOVA方法进行数据分析(P<0.05),正态性检验,方差齐性检验,对于未通过这些检验的数据,使用Kruskal-Wallis H检验(P<0.05)。
2 结果与分析
2.1 中华蜜蜂ZFP37编码蛋白分析
分析中华蜜蜂ZFP37基因序列后发现,其开放阅读框(ORF)全长1 413 bp。该基因编码123个氨基酸,分子量为13.7 kDa,其中包含19种常见氨基酸,不含Trp;Lys含量最高(8.1%),Tyr含量最低(1.6%),等电点为8.72,脂溶指数为53.09,平均亲水力为-0.737,不稳定系数为39.41。
2.2 中华蜜蜂ZFP37的信号肽、跨膜结构、糖基化位点、磷酸化位点预测
中华蜜蜂ZFP37信号肽预测结果显示,其全部氨基酸的S值均在阈值以下,表明其无信号肽区域(图1)。跨膜结构预测结果显示中华蜜蜂ZFP37各氨基酸的预测值均大于阈值1,说明蛋白全部在膜外,无跨膜结构,不属于膜蛋白(图2)。

图1 中蜂ZFP37信号肽结构的预测Fig.1 Predicted signal peptide of AcerZFP37
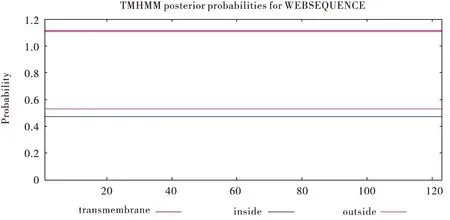
图2 中蜂ZFP37跨膜结构的预测Fig.2 Predicted transmembrane regions of AcerZFP37
蛋白糖基化位点预测结果表明,该蛋白Thr60的O糖基潜力值高于阈值0.5达到0.756,说明中华蜜蜂ZFP37存在O糖基化位点;N-糖基化位点预测显示所有位点潜力值均小于阈值,未预测到带相应的N糖基化位点(图3)。蛋白磷酸化位点预测结果表明:中华蜜蜂ZFP37存在磷酸化位点,且出现在丝氨酸Ser、苏氨酸Thr以及酪氨酸Tyr这 3种氨基酸残基上。包含12个磷酸化位点,分别是Ser20、Thr21、Ser29、Thr39、Thr60、Ser67、Ser68、Tyr69、Ser70、Tyr78、Ser83、Ser107(图4)。

图3 中蜂ZFP37糖基化位点的预测Fig.3 Predicted glycosylation of AcerZFP37
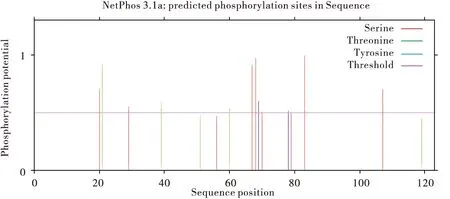
图4 中蜂ZFP37磷酸化位点的预测Fig.4 Predicted phosphorylation sites of AcerZFP37
2.3 中华蜜蜂ZFP37蛋白二级结构和三级结构预测
中华蜜蜂ZFP37蛋白二级结构的预测结果显示:中华蜜蜂ZFP37的二级结构主要包括3种结构:α螺旋、股和环,所占比例分别为8.13%、10.57%和81.3%。而三级结构中主要包括α螺旋和β折叠(图5)。

图5 中蜂ZFP37二级结构和三级结构的预测Fig.5 Predicted secondary and tertiary structure of AcerZFP37注:A,中蜂ZFP37的二级结构;B,中蜂ZFP37的三级结构。Note:A,Secondary structure of AcerZFP37;B,Tertiary structure of the AcerZFP37.
2.4 中华蜜蜂ZFP37的进化树分析
根据NCBI Blast检索到若干膜翅目昆虫ZFP37氨基酸序列,并利用MEGA 7.0软件构建包含中蜂在内的系统进化树,系统进化树结果如图6,结果显示:构建系统进化树所用的昆虫聚类主要分为3个分支,第一分支由中华蜜蜂、大蜜蜂Apisdorsata、迴条蜂Habropodalaboriosal、驼切叶蚁Cyphomyrmexcostatus和切叶蚁Acromyrmexechinatior组成,第二分支为小蜜蜂Apisflorea和美洲东部熊蜂Bombusimpatiens,而第三分支则为佛罗里达弓背蚁Camponotusfloridanus、欧洲熊蜂Bombusterrestris和西方蜜蜂Apismellifera。其中,中华蜜蜂ZFP37序列与大蜜蜂的相似性最高,而与其他膜翅目昆虫的相似性存在差异,表明其具有较高的变异性。
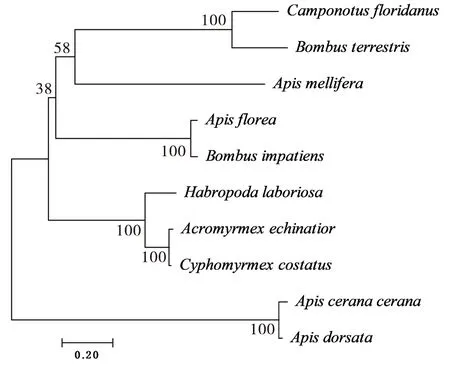
图6 不同昆虫ZFP37氨基酸序列所构建的系统发育树Fig.6 Phylogenetic tree of ZFP37 from different insect species based on amino acid sequences注:中华蜜蜂,Apis cerana cerana,PBC34093.1;西方蜜蜂,Apis mellifera,XP 006562781.1;小蜜蜂,Apis florea,XP 003694613.1;大蜜蜂,Apis dorsata,XP 031367390.1;欧洲熊蜂,Bombus terrestris,XP 012166515.1;美洲东部熊蜂,Bombus impatiens,XP 003492637.1;佛罗里达弓背蚁,Camponotus floridanus,XP 025268432.1;迴条蜂,Habropoda laboriosal,KOC69890.1;驼切叶蚁,Cyphomyrmex costatus,KYN01212.1;切叶蚁,Acromyrmex echinatior,EGI62930.1。
2.5 不同温度胁迫对中华蜜蜂ZFP37表达的影响
中华蜜蜂(20日龄成蜂)暴露于不同温度下(25℃、30℃、35℃、40℃和45℃)2 h,ZFP37在不同温度下的mRNA相对表达量会受高温胁迫的影响,不同温度处理间的表达存在显著差异(P<0.05),ZFP37的表达模式,从25℃至45℃间总体上表现出前低后高的趋势,高温处理能使ZFP37表达上调,尤其是45℃处理组。ZFP37在45℃时表达量最高,且显著高于其他处理组(P<0.05)。
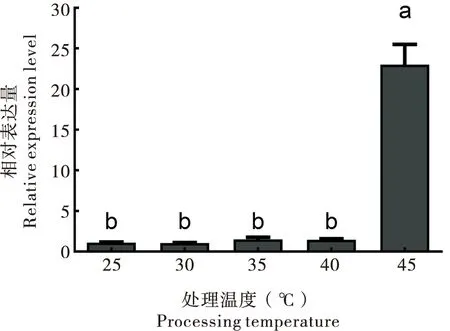
图7 中华蜜蜂ZFP37在不同温度下的mRNA相对表达量Fig.7 Relative expression levels of ZFP37 mRNA in response to different temperature treatments注:图中数据为平均值±标准误;柱上不同的字母表示不同温度处理间差异显著(P<0.05)。下图同。Note:Data were mean±SE.Different letters above bars indicated significant difference at the 0.05 level between different temperature treatments.Same below.
2.6 不同胁迫时间对中华蜜蜂ZFP37表达的影响
中华蜜蜂(20日龄成蜂)暴露于45℃不同处理时间下(0 h、0.5 h、1 h、1.5 h和2 h),ZFP37在不同处理时间下的mRNA相对表达量会受高温胁迫时间的影响,不同胁迫时间处理组的表达存在显著差异(P<0.05),ZFP37的表达模式从0 h至2 h间总体上表现出前低后高的趋势,说明处理时间的延长能使ZFP37表达上调。ZFP37在1.5 h表达水平最高,显著高于0 h和1 h处理组(P<0.05),0 h表达量最低,且显著低于其他处理组(P<0.05)。
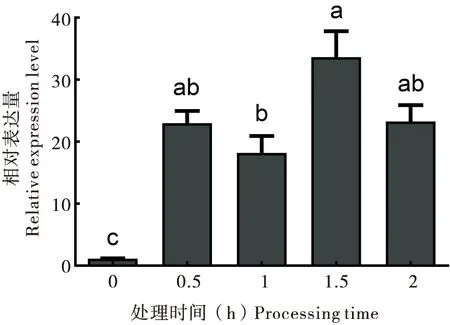
图8 中华蜜蜂 ZFP37在不同胁迫时间下的mRNA相对表达量Fig.8 Relative expression levels of ZFP37 mRNA in response to different process time
3 结论与讨论
锌指蛋白作为一类具有锌指结构的转录因子在真核生物中分布十分广泛,在动植物、酵母等真菌中都有发现(Caoetal.,2016;Yuanetal.,2018)。锌指蛋白参与生命调控的方式主要是与DNA、RNA或蛋白质进行结合,之后在转录水平或翻译水平上对相关生命活动进行调控。研究发现锌指蛋白能够参与到生物的基因调控、细胞分化、胚胎发育、应激反应等生命过程(Nakashimaetal.,2002;Jangetal.,2016;Imbeaultetal.,2017)。动植物在环境压力胁迫下,其体内多个调控蛋白的基因会表达增加,进而调控下游效应基因,改变机体的生理代谢并引发相应的应激反应,帮助生物体应对环境胁迫(Hranitzetal.,2010;kerletal.,2010;Liuetal.,2012;Guanetal.,2016)。本研究发现中华蜜蜂ZFP37无跨膜结构域,不属于膜蛋白,这一结果也与其转录因子的身份相对应。通过序列比对发现中华蜜蜂ZFP37的序列与蜜蜂科昆虫的相似性最高,而与其他膜翅目昆虫相似性存在差异,这可能与昆虫的生活环境和习性有关。
蜜蜂在面对各种环境胁迫时会作出相应的应激反应,其中合成抗逆蛋白是最主要的抗逆方式,常见的抗逆蛋白主要有热激蛋白和锌指蛋白等。而研究发现,昆虫遭受温度胁迫后,一些抗逆基因的高表达能提高昆虫耐受性,在众多抗逆基因中,热激蛋白基因家族发挥着极其重要的作用,在多种应激条件下机体产生的热激蛋白可以显著提高生物的耐受性(Huangetal.,2007)。锌指蛋白的转录调控作用在昆虫响应环境胁迫中发挥重要作用(Andrewetal.,2006),因此,锌指蛋白是帮助生物体应对各类胁迫的重要抗逆蛋白,在生物体抗逆过程中的调控作用一直受到研究者的关注。
本研究结果表明ZFP37在不同温度下的表达模式存在显著差异,在45℃处理下表达水平最高,说明高温能显著提高ZFP37的表达水平,中华蜜蜂ZFP37的表达受高温胁迫的诱导,ZFP37能对高温胁迫做出快速的响应。同样的,高温下不同胁迫时间的处理结果显示,中华蜜蜂ZFP37的表达受胁迫时间的诱导,长时间的高温胁迫可以导致ZFP37表达增加。因此在高温胁迫下,多个锌指蛋白基因被激活,其中既有表达上调的,也有表达下调的,推断ZFP37的活化可能是蜜蜂调节耐受性的一项重要机制。参考对意大利蜜蜂ApismelliferaligusticaZFP37的研究结果,发现两者ZFP37的氨基酸组成及蛋白各项理化性质非常相似,中华蜜蜂和意大利蜜蜂的ZFP37均无信号肽区域,都属于无跨膜结构的膜外蛋白,都含有相应的O糖基化位点和磷酸化位点,且两者的二、三级结构也较为相似,这一结果也与两者亲缘关系较近这一情况相对应。本研究认为ZFP37在中华蜜蜂和意大利蜜蜂体内的作用是相似的,都可能参与了蜜蜂抗逆过程的代谢调节。蜜蜂调节耐受性的一项重要机制锌指蛋白对蜜蜂的耐热性具有重要的调节作用,但其具体的调节机制仍不清楚,还需要进一步的深入研究。
猜你喜欢
今日农业(2022年16期)2022-11-09
分子催化(2022年1期)2022-11-02
今日农业(2022年15期)2022-09-20
环球时报(2022-06-20)2022-06-20
中国核电(2021年3期)2021-08-13
昆明医科大学学报(2021年3期)2021-07-22
烟草科技(2021年6期)2021-06-24
电脑知识与技术(2018年19期)2018-11-01
小天使·一年级语数英综合(2017年9期)2017-10-20
阅读与作文(小学高年级版)(2016年5期)2016-05-10
