
干旱胁迫对屋顶绿化植物小叶黄杨叶片抗氧化特性的影响*
2022-09-21 11:08张崇洋刘世亮周孟椋吴苗苗姜桂英
林业与环境科学 2022年3期
刘 慧 张崇洋 刘世亮 周孟椋 吴苗苗 姜桂英 刘 芳 赵 颖
(河南农业大学 资源与环境学院,河南 郑州 450002)
伴随着城市化进程的不断加快,钢筋水泥等建筑日益增多,绿地面积日渐减少,导致许多城市的高温、干燥等环境愈加严重。屋顶绿化以其独特的优势,如集中节约利用城市空间、增加绿地面积、提高生态环境质量等受到更多的关注和快速发展[1-2]。但这独特的立地条件使植物种植与地面隔离,缺少地下水补给,且光照强烈、夏天高温、高风速、水分挥发较快,对植物抗旱性要求非常高[3]。
小叶黄杨Buxus microphylla属黄杨科黄杨属植物,常绿灌木或小乔木,枝叶茂密、常青,有研究表明小叶黄杨具备较强的耐旱性[4]。近年来,有关干旱胁迫对小叶黄杨幼苗生理特性的影响主要在小叶黄杨的耗水规律和光合作用变化特征[5]、水分利用效率[6]等方面进行了研究。对干旱胁迫下小叶黄杨保护酶活性、膜脂过氧化及渗透调节等方面研究较少。干旱胁迫往往会导致植物体内活性氧(ROS)累积,进而对植物造成氧化胁迫。为保护机体免受损害,植物进化出了抗氧化系统以有效清除ROS,因此,植物叶片的抗氧化酶是植物抗逆性生理评价的重要指标。在自然选择和遗传变异的双重作用下,植物对干旱胁迫的抗性逐渐形成,这种抗逆性与植物的生理状态和内部结构密切相关[7]。因此,研究不同干旱胁迫下小叶黄杨抗氧化酶活性和渗透调节物质含量的变化,对了解小叶黄杨适应干旱机制具有重要意义。本文通过对小叶黄杨在不同干旱胁迫条件下叶片抗氧化特性的变化特征进行了研究,以期为小叶黄杨屋顶环境种植提供理论依据。
1 材料与方法
1.1 研究区概况
在河南省郑州市金水区硅谷广场楼顶(E113°40′~113°47′,N34°50′~34°57′)进 行 试 验,该区属于温带季风气候,年平均气温14.8 ℃,降雨量586.1 mm,其中郑州市6—8月降雨量达到322 mm,8月平均气温25.9 ℃,屋顶温度最高可达50~55 ℃。
1.2 试验材料
供试小叶黄杨为两年苗龄,由郑州市陈寨花卉市场购得。
供试土壤采自郑州市郊农田土壤(0~20 cm),属典型褐土,基本理化性质如下:pH 7.79,EC值0.11 ms·cm-1,有机质含量7.77 g·kg-1,碱解氮、有效磷、速效钾含量44.97 mg·kg-1,7.44 mg·kg-1,62.16 mg·kg-1,田间持水量32.65%。
1.3 试验方法
于2019年6—9月进行试验,选用高30 cm直径40 cm的塑料盆,每盆装土12.50 kg,移栽前施用2.50 kg的商品有机肥(有机质≥45%,N+P2O5+K2O≥6%)。
试验共设4个土壤水分处理:正常水分处理,田间持水量的80%~90%(CK);轻度干旱胁迫,田间持水量的60%~70%(TB);中度干旱胁迫,田间持水量的50%~60%(TC);重度干旱胁迫,田间持水量的35%~45%(TD)。每个处理设3次重复(即每个处理种植3盆),每盆种植1株小叶黄杨,进行正常栽培管理,采用称量法控制土壤含水量。
盆栽置于塑料大棚下,每天18:00时称重补充水分。在移栽后0、20、40、60、80 d,采集由顶端往下第6~8片完全伸展叶,采集后迅速用液氮冷冻,置于-80 ℃冰箱保存,用于相关指标测定。
1.4 项目测定
土壤pH、EC采用多参数分析仪(沪制02220142)测定;有机质含量采用重铬酸钾容量法—外加热法测定;碱解氮含量采用碱解扩散法测定;速效磷含量采用0.5 mol·L-1NaHCO3浸提—钼锑抗比色法测定;速效钾含量采用火焰光度法测定[8]。
叶片过氧化氢酶(CAT)活性采用紫外分光光度法测定[9];丙二醛(MDA)含量采用硫代巴比妥酸法测定[10];超氧化物歧化酶(SOD)和过氧化物酶(POD)活性采用南京建成公司生产的SOD试剂盒和POD试剂盒测定[11];可溶性蛋白质含量采用考马斯亮蓝G-250法测定[9]。
1.5 数据分析
试验数据为3次重复的平均值,使用WPS 2016软件整理计算,不同干旱胁迫处理间各种指标之间的差异由单因素方差分析中Duncan检验完成,显著水平为0.05。采用Origin 8.5软件绘图。
2 结果与分析
2.1 不同干旱胁迫处理对小叶黄杨叶片抗氧化酶活性的影响
2.1.1 超氧化物歧化酶(SOD)活性 由图1可知,随胁迫时间的延长,SOD活性变化趋势呈升高-降低-升高,20 d时,不同干旱胁迫处理间SOD差异不显著,但均显著低于CK处理(P<0.05),分别比CK处理降低了39.02%、31.44%、28.60%;40 d时,TB与TC处理的SOD活性达到最低,且显著低于CK与TD处理(P<0.05),TD与CK处理差异不显著;60 d时,TB和TD处理SOD活性显著低于CK处理(P<0.05),TC与CK处理差异不显著;80 d时,TC处理SOD活性最低为10.04 U·g-1,且仅TC处理显著低于CK处理(P<0.05)。

图1 不同干旱胁迫处理下小叶黄杨叶片SOD活性的变化Fig.1 Change of SOD activity in B. microphylla leaves under different drought stress treatments
2.1.2 过氧化氢酶(CAT)活性 由图2可知,随干旱胁迫程度和胁迫时间的增加,小叶黄杨叶片CAT活性呈先升高后降低的变化趋势。20 d时,TB、TC、TD处理的CAT活性均显著低于CK处理(P<0.05);40 d时,TC处理CAT活性显著高于CK处理(P<0.05),TB和TD处理CAT活性也均高于CK处理,但差异不显著;60 d和80 d时,各处理的CAT活性均显著低于CK处理(P<0.05)。其中,60 d时,CK处理CAT活性最高为19.27 U·g-1,分别比其他处理高了17.92%、24.37%、32.57%。
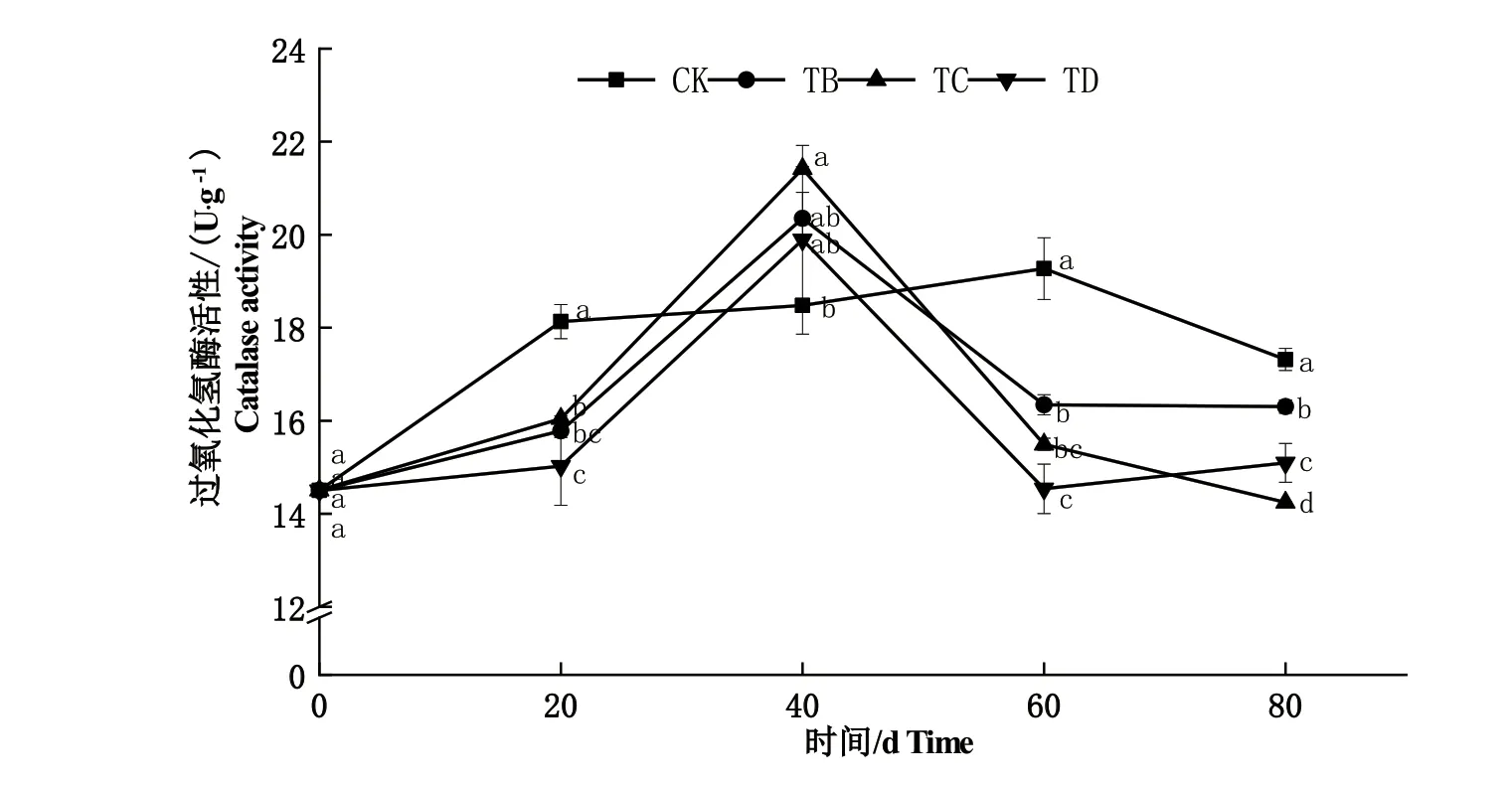
图2 不同干旱胁迫处理下小叶黄杨叶片CAT活性的变化Fig.2 Change of CAT activity in B. microphylla leaves under different drought stress treatments
2.1.3 过氧化物酶(POD)活性 由图3可知,随干旱胁迫时间的延长,各处理的POD活性呈先升高后降低再升高的变化趋势,在40 d达到峰值,分别比0 d增加了366.49%、461.06%、450.46%、335.35%;20 d时,不同干旱胁迫处理的POD活性均显著低于CK处理(P<0.05),而TC处理的POD活性显著高于TB与TD处理(P<0.05);40 d时,TB和TC处理的POD活性显著高于CK处理(P<0.05),而TD处理的POD活性低于CK处理,但差异不显著;60 d时TB、TC、TD处理的POD活性均显著低于CK处理(P<0.05);80 d时,TB、TC、TD处理的POD活性分别比CK处理低了10.86%、14.75%、17.68%,且TC处理分别与TB、TD处理差异不显著,TB和TD处理间差异显著(P<0.05)。

图3 不同干旱胁迫处理下小叶黄杨叶片POD活性的变化Fig.3 Change of POD activity in B. microphylla leaves under different drought stress treatments
2.2 不同干旱胁迫处理对小叶黄杨叶片丙二醛(MDA)含量的影响
由图4可知,随干旱胁迫时间的延长,各处理的MDA含量大体上呈先降低后升高的变化趋势,在干旱胁迫末期(80 d)均达最大值。与胁迫初期(0 d)相比,TB、TC、TD处理在胁迫末期(80 d)的MDA含量分别提高126.33%、1.46%、5.58%;20 d时,TB、TD处理的MDA含量显著高于CK处理(P<0.05),而TC处理的MDA含量略低于CK处理;40 d时,只有TB处理的MDA含量显著高于CK处理(P<0.05);60 d时,TB、TC和TD处理的MDA含量显著高于CK处理(P<0.05);80 d时,TB、TC和TD处理的MDA含量均显著低于CK处理(P<0.05)。此 外,80 d与60 d时 相 比,TB、TC和TD处理的MDA含量分别提高323.51%、109.95%、103.61%。

图4 不同干旱胁迫处理下小叶黄杨叶片MDA含量的变化Fig.4 Change of MDA content in B. microphylla leaves under different drought stress treatments
2.3 不同干旱胁迫处理对小叶黄杨叶片可溶性蛋白质含量的影响
由图5可知,随着干旱胁迫时间的增加和程度的加深,小叶黄杨叶片可溶性蛋白质含量整体趋势先上升,60 d时达到峰值后下降。20 d时,TB、TC、TD处理的可溶性蛋白质含量均显著高于CK处理(P<0.05),其中TD处理的可溶性蛋白质含量最高为2.59 mg·g-1;40 d时,TB和TC处理显著低于CK处理(P<0.05),而TD处理的可溶性蛋白质含量略高于CK处理,但差异不显著;60 d时,TB和TD处理的可溶性蛋白质含量均高于CK处理,其中TD处理显著高于CK处理(P<0.05)且含量最高为3.33 mg·g-1,80 d时,除TC处理显著高于CK处理(P<0.05)外,其他处理与CK处理相比均差异不显著。
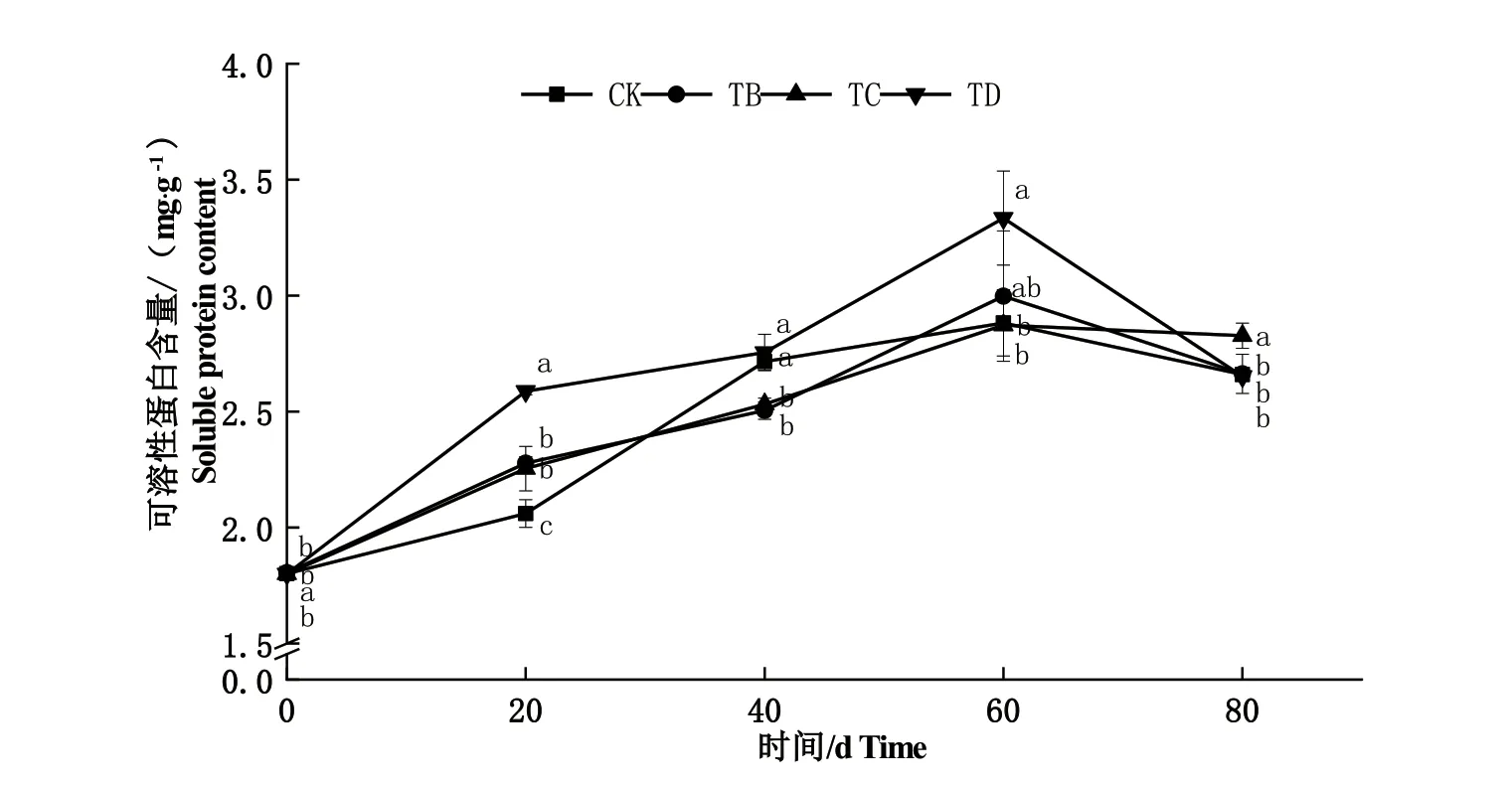
图5 不同干旱胁迫处理下小叶黄杨叶片可溶性蛋白质含量的变化Fig.5 Change of soluble protein content in B. microphylla leaves under different drought stress treatments
2.4 不同干旱胁迫处理下各指标相关性
由表1可知,POD活性与SOD活性、CAT活性、MDA含量、可溶性蛋白质含量均呈正相关关系,其中POD活性与CAT活性和可溶性蛋白质含量(P<0.01)呈极显著相关关系,SOD活性与MDA含量(P<0.05)呈显著相关关系;CAT活性和可溶性蛋白质含量均与SOD活性、MDA含量呈负相关关系。
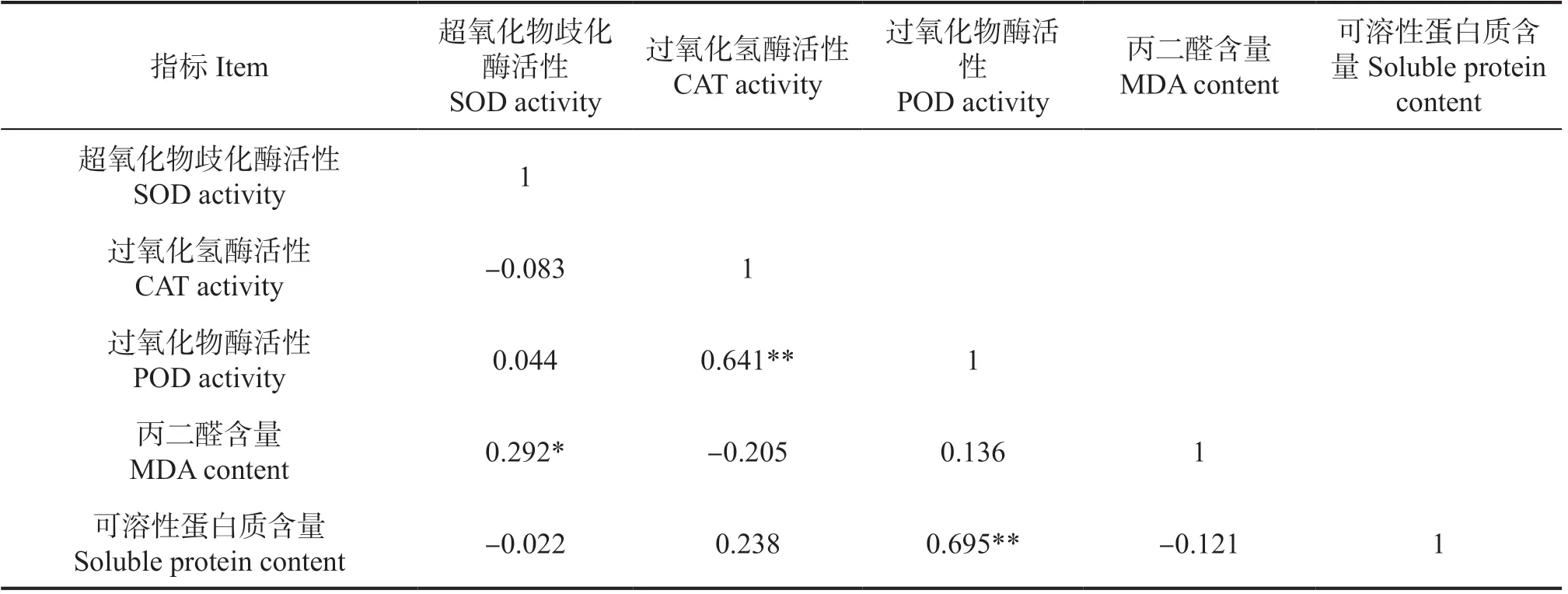
表1 小叶黄杨叶片各指标相关性分析Table 1 Correlation analysis of each indicator in B. microphylla leaves
3 讨论与结论
3.1 干旱胁迫下过量的活性氧(ROS)会造成细胞膜脂过氧化,损坏植物细胞和膜,抗氧化酶可以促进植物抗逆性和防御系统[12-13]。研究表明,SOD在干旱胁迫初期0~20 d时,含量增加迅速,20 d时达到高峰。表明0~20 d时,小叶黄杨主要靠SOD清除·O2-,将其歧化为H2O2,进而缓解干旱胁迫带来的伤害[14]。相关研究表明,随着干旱胁迫的增强,孟连苗叶片的SOD活性呈现先升高后下降再升高变化的“S”曲线[15],本试验所得结论与上述观点一致。此外,本研究发现,正常水分处理的SOD活性在20~60 d时,呈下降趋势。这可能是因为6—8月屋顶温度较高,水分蒸发较快,一定程度上抑制了正常水分处理的SOD活性,使其整体下降。
本研究发现,在一定强度干旱胁迫下,小叶黄杨叶片CAT活性增加,严重的干旱胁迫可能会抑制CAT活性,40 d时CAT活性达到峰值。CAT可以将H2O2直接转化成H2O,降低活性氧的含量。此外,清除H2O2可以清除细胞中的活性氧,缓解植物氧化损伤[16-17]。本研究中,CAT活性在20~40 d时,呈上升趋势,40 d后下降。可能是因为6月20日—7月10日温度逐渐升高,干旱胁迫对小叶黄杨的影响日益明显,而7月10日—8月温度较高抑制了小叶黄杨的正常生长和代谢,进而CAT活性随之降低。熊仕发等[18]研究表明,在干旱胁迫过程中,小叶黄杨叶片CAT活性变化呈现先上升后下降趋势,与本研究结果一致。
本研究发现,在0~60 d随着干旱胁迫的加剧,小叶黄杨叶片中POD活性先上升后下降,各干旱胁迫处理下的POD活性与CK处理相比均差异显著,这可能是由于干旱胁迫下的膜脂过氧化作用,使得H2O2积累而导致SOD活性减弱,将超氧阴离子转化为H2O2的能力降低,H2O2诱导POD活性增强,POD将H2O2分解,防止H2O2与剩余超氧阴离子反应生成活性更强的OH-,加剧氧化胁迫,从而达到维护机体免受活性氧的损伤[19]。这与前人研究结果一致[20-21],但60 d后POD活性又上升,这可能是因为气温下降缓解了干旱胁迫程度。
MDA含量在后期尤其是60 d后显著增加,质膜被破坏,大量离子外溢,其中轻度胁迫下质膜的损害严重,很多研究[22-25]得出了与之相似的结果。干旱在前期不会对膜造成太大伤害,后期干旱程度加剧MDA含量增加。在轻度胁迫下MDA含量增加幅度已很大,尤其60 d后MDA含量出现大幅度增加,表明在干旱胁迫下,小叶黄杨体内氧自由基的积累导致MDA大量增加,从而使植株免受严重损害,赵婵璞等[26]对匍枝委陵菜Potentilla flagellaris的研究与本试验研究结果一致。3.2 可溶性蛋白质是植物体内重要的渗透调节物质,能明显提高细胞的结合水含量和持水能力[27]。本研究发现,随着胁迫时间的增加,各处理可溶性蛋白质含量呈现先升高后降低的趋势,可溶性蛋白质含量的最大值出现在60 d,这与高鹏飞等[28]的研究结果一致。小叶黄杨的叶片因为膜受损伤蛋白质被释放出来,蛋白酶活性升高将蛋白质水解,而水解所得的氨基酸又用于合成如抗氧化酶等响应干旱胁迫的蛋白质,导致小叶黄杨叶片前60 d的可溶性蛋白质含量增加[29],这也表明小叶黄杨有一定抗旱性,与张卫红等[30]对垂穗披碱草的研究结果一致。随着胁迫强度的增加,在一定程度上抑制了蛋白质的合成或加快其降解速度,导致可溶性蛋白质含量在60 d后下降,这与王晓宇等[31]研究结果相一致。也有相关研究表明,干旱可抑制蛋白质合成,诱导蛋白质降解,从而降低某些植物的总蛋白质含量[32]。
此外,各处理在60 d之前可溶性蛋白质含量与干旱持续时间正相关,可溶性蛋白质含量的增加也许能使植物细胞保持在低渗透电位水平,保护细胞结构,延缓衰老,抵抗干旱胁迫造成的损害。随着干旱胁迫的加剧,可溶性蛋白质含量的下降幅度也随之增大,刺激了小叶黄杨保护酶系统做出保护性反应,但随着胁迫时间的延长,各胁迫处理的保护酶活性在达到最大值后均呈下降趋势,这可能是因为过量ROS或H2O2积累,超过保护酶系统阈值,打破动态平衡,致使酶光钝化,抑制新酶的合成。
3.3 在正常条件下,植物活性氧的产生和清除处于动态平衡状态,但在干旱胁迫下,这种平衡被打破,植物为维持正常的生长代谢,必须依靠抗氧化酶系统清除活性氧[33]。本研究发现,在干旱胁迫的不同时期,POD和CAT活性的降低与MDA的积累紧密相关,两者可能互为因果关系。一方面,POD和CAT活性降低使有害自由基积累超过损伤阈值,可能直接或间接地引发膜脂过氧化,增加MDA含量,损伤膜系统;另一方面,随着MDA的积累,POD和CAT活性受到抑制后降低,从而失去了保护酶系统的功能,进而加重膜系统的损伤。说明植物抗旱能力与保护酶活性及其防御功能有关[34],这可能是植物在干旱胁迫下的主要生理反应和损伤机制。此外,可溶性蛋白质与POD活性和MDA含量关系密切。在干旱胁迫前期(0~40 d),可溶性蛋白质与POD活性变化一致,说明二者协同作用以适应干旱逆境造成的伤害。在整个干旱胁迫过程中,可溶性蛋白质与MDA含量的变化趋势相反,可能是可溶性蛋白含量的增加减轻了MDA的积累,进而减轻干旱对植物的损伤。
综上所述,干旱胁迫后,小叶黄杨体内的保护酶系统做出响应,使机体在一定程度上可以维持正常的生长和代谢等活动。在轻度和中度干旱胁迫后,保护酶活性的提高可以有效协调和维持氧化物含量的相对平衡,减少有害氧化物积累所造成的损害;重度胁迫后,小叶黄杨保护酶系统和细胞遭受不可逆的损害,不能发挥正常的保护机体的功能。总之,小叶黄杨作为一种绿色植物,具有较强的抗旱性,在很大程度上能够适应逆境,能够在屋顶这种特殊的环境下正常生长。
猜你喜欢
现代农业科技(2022年5期)2022-12-14
科学导报(2022年11期)2022-03-03
食品安全导刊(2021年20期)2021-08-30
昆明医科大学学报(2021年8期)2021-08-13
当代水产(2021年6期)2021-08-13
智慧健康(2020年9期)2020-12-03
建材发展导向(2019年10期)2019-08-24
时尚北京(2016年12期)2017-01-16
农家顾问(2016年7期)2016-05-14
医学研究杂志(2015年9期)2015-07-01
