
投饲率对鲻幼鱼生长性能和生理机能的影响
2022-09-21 09:12:40许明珠董兰芳韦媛媛
水产科学 2022年5期
童 潼,许明珠,董兰芳,韦媛媛,梁 钰,张 琴
( 1.广西民族大学 海洋与生物技术学院,广西多糖材料与改性实验室,广西高校微生物与植物资源利用重点实验室,广西 南宁 530006;2.广西海洋研究所有限责任公司,广西海水养殖新品种繁育工程技术研究中心,广西 北海 536000 )
鲻(Mugilcephalus)属硬骨鱼纲鲻科鲻属,俗称乌头、白眼等,主要分布于我国渤海、黄海、东海以及南海海域[1-2]。鲻属广温性(3~35 ℃)和广盐性(0~40)鱼类,其肉质鲜美,营养价值高,富含蛋白质、脂肪酸、维生素E、钙、镁等营养物质,并兼具食疗药用价值,深受广大消费者喜爱。同时,鲻还具有生长速度快、生存能力强、疾病少、对饲料要求低、个体大等特点,近年来成为许多国家水产养殖的重要经济鱼类[3]。
投饲率是影响鱼类生长的重要因素之一,在适宜的范围内,随着投饲率的增加,鱼类的生长性能增强,超出或低于适宜的投饲率,鱼类的生长会受到不同程度的影响。因此确定养殖鱼类最适投饲率,并提高养殖鱼类品质和养殖经济效益是当前水产养殖业的研究热点之一。投饲率又称日投饲率,是指日投喂的饲料质量占摄食鱼体质量的百分数。迄今,已有学者研究了投饲率对吉富罗非鱼(Oreochromisniloticus)[4]、日本黄姑鱼(Nibeajaponica)[5]、花鲈(Lateolabraxmaculatus)[6]及异育银鲫(Carassiusauratusgibelio)[7]等鱼类养殖生长的影响。而关于鲻的研究主要集中在人工养殖技术[8-10]、人工繁殖与其组织学[11-12]、细胞学[13]等方面,投饲率对鲻的影响研究尚未见报道。笔者通过比较分析不同投饲率对鲻生长性能、体成分、消化酶活性和肝脏免疫酶活性的指标变化,确定最适宜的投饲率,旨在为今后鲻养殖过程中科学合理的饲料投喂策略提供理论依据,以促进鲻生长发育、提高饲料利用率、提高经济效益和扩大鲻生产量。
1 材料与方法
1.1 试验设计与饲养管理
试验于2019年8月4日开始,于广西海洋研究所海水增养殖试验基地进行,为期8周。根据初期预试验结果(饱食水平为10%),设置饥饿、半饱食、饱食和过饱食4个投饲率,分别对应饲料投喂量为鲻体质量的2.5%、5%、10%和15%。试验所用鲻苗种购于广东省湛江市吴川鱼苗场,选取同一批健康无病害、体质量相近[平均体质量(37.96±0.98) g]的鲻幼苗240尾,随机分成4组,每组3个平行,每个平行20尾。投喂饲料为健马牌鳗鱼粉(蛋白含量43.8%,脂肪含量4.9%,水分含量9.5%)。试验于2 m×2 m的水泥池中进行,水深1 m,水温26~30 ℃,盐度21~25,流水3 h/d,7 d更换池水1次,连续充氧。试验期间,每隔10 d取样称量质量1次,根据鲻体质量情况更换饲料投喂量,观察记录鲻每天的摄食生长情况。
1.2 样品采集及处理
饲养试验结束后进行样品采集与处理。取样前将鲻禁食24 h,以每个平行为单位进行打捞,当场记录每组鲻的总数量与总质量,每组随机选取4尾测量体长、体质量、肝脏质量、内脏团质量,得到的数据用于计算每个平行试验鲻的质量增加率(wWGR,%)、特定生长率(RSG,%/d)、成活率(RS,%)、饲料利用率(wFCR,%)、肥满度(CF)、肝体指数(wHSI,%)和脏体指数(wVSI,%)。
wWGR=(mt-m0)/m0×100%
RSG= (lnmt-lnm0)/t×100%
RS=nt/n0×100%
wFCR= (mt-m0)/mf×100%
CF=100m/L3
wHSI=mh/m×100%
wVSI=mv/m×100%
3.2 播种造林法:又称直播造林,是将林木种子直接播种在造林地进行造林的方法。这种方法省去了育苗工序,而且,施工容易,便于在大面积造林地上进行造林。但是这种方法造林对造林立地条件要求较严格,造林后的幼林抚育管理措施要求也较高。播种造林的适用条件∶适合于种粒大、发芽容易、种源充足的树种,如橡栎类、核桃、油茶、油桐和山杏等大粒种子。其要求造林地土壤水分充足,各种灾害性因素较轻,对于边远且人烟稀少地区的造林更为适宜。
式中,mt为终末平均体质量(g),m0为初始平均体质量(g),t为试验时间(d),n0为初始尾数(尾),nt为终末尾数(尾),mf为总投喂量(g),m为体质量,L为体长,mh为肝脏质量(g),mv为内脏团质量(g)。
1.2.2 体成分的测定
每个平行随机抽取2尾鲻,采用美国分析化学家协会(AOAC 1995)[14]的方法分析试验鲻的体成分和饲料常规成分。即105 ℃恒定质量法测定全鱼水分含量,索氏抽提法测定全鱼粗脂肪含量,凯氏定氮法测定全鱼粗蛋白含量,550 ℃煅烧法测定全鱼灰分含量。
1.2.3 消化酶和肝脏免疫酶活性的测定
每个平行随机抽取5尾活体鲻进行解剖采样,获取肝脏并记录其质量。将肝脏放入玻璃匀浆器内,按照质量体积比1 g∶1 mL的比例加入pH 7的磷酸盐缓冲液进行研磨捣碎,将研磨液转移至 1.5 mL的离心管中,4 ℃ 3000 r/min(离心半径6.5 cm)离心10 min,用移液枪吸取上清液,所得上清液即为待测酶液,以上操作均在低温冰浴的条件下进行,待测酶液均置于-80 ℃下保存。消化酶、过氧化氢酶、超氧化物歧化酶、溶菌酶活性均采用南京建成生物工程研究所生产的测定试剂盒进行测定,按照试剂盒操作步骤进行操作,每组重复测定3次。
1.3 数据统计与分析
采用SPSS 19.0软件对试验结果进行统计分析,数据采用平均值±标准差表示,若试验结果误差达到显著水平(P<0.05),则采用图基法进行多重比较。
2 结 果
2.1 不同投饲率对鲻生长性能的影响
投饲率对鲻质量日增加率、特定生长率、肥满度以及肝体指数有显著影响(P<0.05)(表1)。随着投饲率的上升,鲻的质量增加率和特定生长率呈先升后降趋势(P<0.05),在投饲率为10%时达到最大值,与15%试验组差异不显著,但显著高于2.5%与5%试验组。投饲率对鲻的肥满度有显著影响(P<0.05),在投饲率为10%试验组达到最大,显著高于2.5%试验组。投饲率同样对肝体指数有显著影响(P<0.05),投饲率较高的试验组(10%、15%)肝体指数显著高于投饲率较低的试验组(2.5%、5%),但投饲率高的两组之间差异不显著(P>0.05),投饲率低的两组差异也不显著(P>0.05)。不同投饲率对鲻的存活率和脏体指数影响不显著(P>0.05),各组成活率均为100%,脏体指数为5.33%~6.40%。不同投饲率度鲻生长曲线的影响见图1,10%试验组鲻生长速度最快。
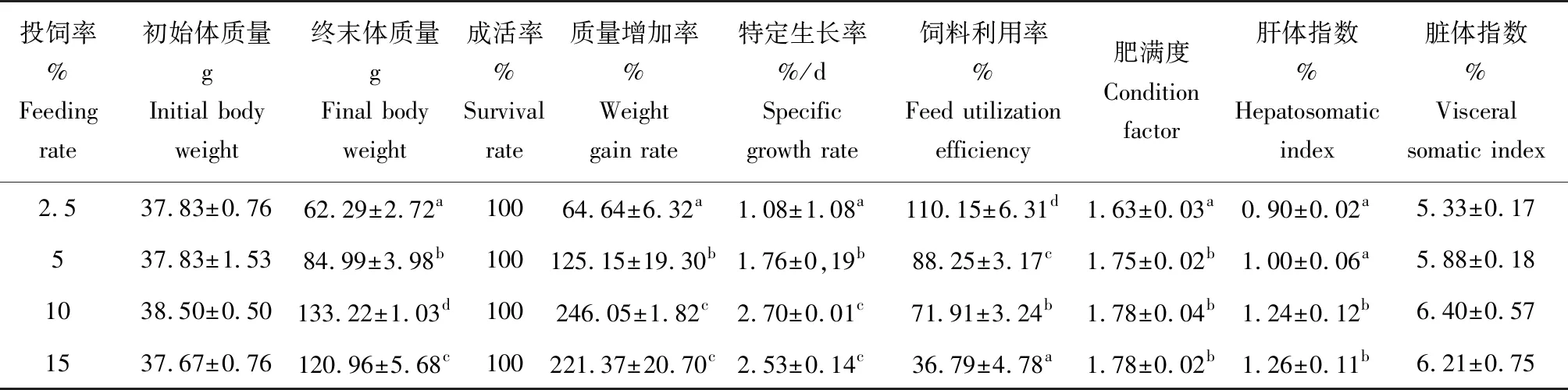
表1 不同投饲率对鲻生长性能的影响(平均值±标准差)Tab.1 The effect of different feeding rate on growth performance of juvenile grey mullet M.cephalus (mean±SD)
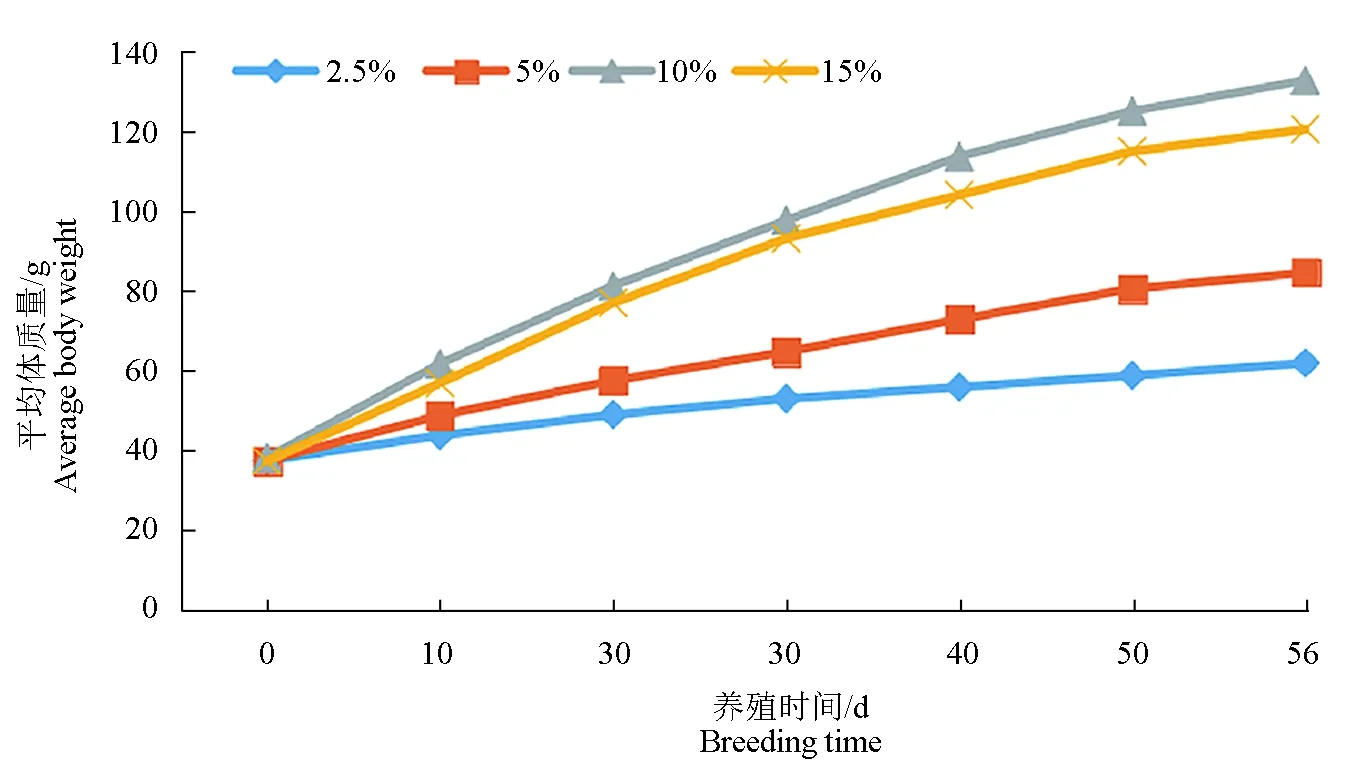
图1 不同投饲率对鲻生长曲线的影响Fig.1 Effects of different feeding rates on growth curve of juvenile grey mullet M. cephalus
2.2 不同投饲率对鲻体成分的影响
投饲率对鲻鱼体水分、粗蛋白以及粗脂肪含量均有显著影响(P<0.05),对鲻鱼体灰分含量影响不显著(P>0.05)(表2)。鲻鱼体的水分含量随着投饲率的增加呈现出逐渐降低趋势,投饲率 2.5% 试验组鱼体水分含量最大,显著高于15%试验组(P>0.05),与5%和10%试验组差异不显著。鲻鱼体粗蛋白含量同样随着投饲率的增加呈现出持续下降的趋势,投饲率2.5%试验组显著高于10%和15%试验组(P<0.05)。鲻鱼体粗脂肪含量则随着投饲率的增加呈现持续上升的趋势,投饲率15%试验组鱼体粗脂肪含量最大,显著高于2.5%和5%试验组(P<0.05)。

表2 不同投饲率对鲻体成分的影响(平均值±标准差) %Tab.2 Effect of different feeding rates on the body composition of juvenile grey mullet M. cephalus (mean±SD)
2.3 不同投饲率对鲻肝脏消化酶活性的影响
不同投饲率对鲻肝脏蛋白酶和脂肪酶活性影响显著(P<0.05),对淀粉酶活性影响不显著(P>0.05)(表3)。鲻肝脏蛋白酶活性随着投饲率的增加先升后降,投饲率10%试验组鲻肝脏蛋白酶活性最强,达到56.39 U/mg,投饲率2.5%试验组蛋白酶活性显著低于其他各组。投饲率的改变同样对鲻肝脏脂肪酶活性有显著影响(P<0.05),投饲率10%试验组脂肪酶活性显著高于2.5%和15%试验组。各试验组肝脏淀粉酶活性差异不显著(P<0.05),活性为0.22~0.33 U/mg。

表3 不同投饲率对鲻肝脏消化酶活性的影响(平均值±标准差)Tab.3 Effect of different feeding rates on liver digestive enzyme activities of juvenile grey mullet M. cephalus (mean±SD)
2.4 投饲率对鲻肝脏免疫酶活性的影响
不同投饲率对鲻肝脏过氧化氢酶活性、超氧化物歧化酶活性、溶菌酶活性均有显著影响(P<0.05)(表4)。随着投饲率的增加过氧化氢酶活性呈现出显著上升的趋势,投饲率15%试验组过氧化氢酶活性最大,显著高于2.5%试验组(P<0.05),但与5%和10%试验组差异不显著(P>0.05);各试验组超氧化物歧化酶活性随着投饲率的上升呈先升后降的趋势,投饲率 10%试验组超氧化物歧化酶活性最大,显著高于2.5%和15%试验组(P<0.05);投饲率10%试验组溶菌酶活性最大,其他组差异不显著。

表4 不同投饲率对鲻肝脏免疫酶活性的影响(平均值±标准差) U/mgTab.4 Effect of different feeding rates on the liver immune enzyme activity of juvenile grey mullet M. cephalus (mean±SD)
3 讨 论
3.1 不同投饲率对鲻生长性能的影响
水产养殖实际生产中确定养殖品种适宜投饲率,既能有效提高鱼体生长性能,也能避免过量投饲引起的资源浪费和养殖水质恶化,或投饲量过低引起的鱼体规格大小不匀、品质不佳等问题[15-16],以取得最佳养殖效益。本试验结果显示,鲻幼鱼的饲料系数随着投饲率(2.5%~15%)的增加呈下降趋势,各组之间差异显著,这与锦鲤(Cyprinuscarpiokoi)幼鱼[16]、达氏鳇(Husodauricus)幼鱼[17]等的研究结果相似。而鲻的质量增加率和特定生长率随投饲率(2.5%~15%)的增加呈先增后趋于平稳的趋势,在投饲率10%时达到最大值,与15%试验组差异不显著,但显著高于2.5%、5%试验组,这与对日本黄姑鱼幼鱼[5]、黄鳝(Monopterusalbus)[15]、大菱鲆(Scophthalmusmaximus)[18]和褐牙鲆(Paralichthysolivaceus)[19]等鱼类的研究结果相似。饲料利用率、质量增加率和特定生长率是衡量鱼类生长的重要指标,低投饲率试验组虽可获得较高的饲料利用率,但也会因为鱼摄食不足而影响其生长。适当提高投饲率后,鲻幼鱼饲料利用率下降,但整体上可用于生长的能量增加,鲻幼鱼的质量增加率和特定生长率得到显著提升。而过量投喂则导致饵料残余,饲料利用率降低的同时还会污染水质,最终影响鲻的食欲和生长发育。
鱼类的肥满度是反映肥瘦程度和生长情况的重要指标,笔者发现,在投饲率小于10%时,鲻的肥满度逐步上升,之后继续升高投饲率,鲻肥满度趋于稳定。分析原因:在适当范围内(2.5%~10%)提高投饲率,鲻摄食的营养物质与能量随着投饲率的增加得到补充,肥满度随之提高[17,19];超过适当范围,由于鲻消化能力有限,肥满度不再增加。鱼类肝体指数、脏体指数是体现长期或短期营养方式变化的指标,肝脏作为鱼体内的营养储存所,在大分子营养物质的代谢中起着重要的作用[20]。本试验结果显示,鲻的肝体指数和脏体指数同样随着投饲率的增加呈先升高后趋于稳定的趋势,这一结果与对花鲈[6]和异育银鲫[7]的研究结果相似。投饲率2.5%试验组肝体指数最低,说明投饲率低的情况下鲻摄食量不足,物质能量大多用于鱼体生长发育,肝脏储存营养物质较少导致肝体指数较低。当投饲率升至10%和15%时,鲻肝体指数达到一定值后趋于稳定,表明投饲率越高,肝脏代谢负荷越重,过多的能量物质在肝脏组织中转变为脂肪沉积在肝脏中,从而使鱼体肝体指数增加[15],达到最大承受量后不再储存。不同投饲率对鲻脏体指数无显著影响,不同组间脏体指数无显著差异,表明鲻生长过程中各脏器之间与体质量比值恒定,脏器总体发育正常。
3.2 不同投饲率对鲻体成分的影响
笔者测定了不同投饲率下鲻的水分、粗蛋白、粗脂肪以及灰分含量。试验结果表明,投饲率对鲻水分、粗蛋白、粗脂肪含量均有显著影响(P<0.05),对灰分含量无显著影响(P>0.05)。随着投饲率的上升,试验组鲻肌肉水分含量呈下降趋势,投饲率15%试验组水分含量显著低于其他各组,这与对日本黄姑鱼幼鱼[5]、大杂交鲟(Husodauricus♀×A.ruthenus♂)幼鱼[21]、达氏鳇幼鱼[17]、锦鲤幼鱼[16]等的研究结果类似。在低投喂水平下,水分含量较其他组高,原因可能是:在饥饿状态下,鱼体首先消耗自身的糖原和脂肪以补充身体正常活动,其次消耗蛋白质,由于身体大量营养物质被消耗,在保持体质量没有显著波动的条件下,肌肉的水分含量相对增多[22]。脂肪含量是衡量食品营养价值的重要指标,在一定范围内肌肉的脂肪含量与肉质的风味呈正相关关系[23]。Mihelakakis等[24]研究发现,随着摄食水平的提高,金头鲷(Sparusaurata)的脂肪含量显著增加;李文龙等[17]对达氏鳇幼鱼研究发现,随着投饲率升高达氏鳇体组成脂肪含量也逐渐增加;陈云飞等[15]报道,随着投喂水平的升高,大、小规格黄鳝的粗蛋白含量整体呈下降趋势,粗脂肪含量呈显著上升趋势。本试验结果显示,随着投饲率的升高,鲻肌肉粗蛋白含量显著降低,粗脂肪含量显著升高,与上述研究结果相似。分析原因可能为:当鱼体处于饥饿状态时,由于摄食的营养物质不能满足日常生活需要,鱼体先消耗自身肌肉储存的脂肪供能,进而蛋白沉积多。随着投饲率升高,组间鱼体粗蛋白含量差异不大,说明摄食饵料中营养物质的增加已经满足了鲻体内蛋白质的需求[25],体内调节生长的相关因子通过不同的作用途径,将获得营养物质的增益部分用于生长。高摄食水平的肌肉粗脂肪含量最高,粗蛋白含量最低,说明此时投喂量过大,鱼体处于过饱食状态,从饲料中摄食到的营养物质充分满足鱼体各项组织需要,只能将过量的物质能量转化为脂肪沉积在肌肉中,做储备能量使用[15]。还可能存在的影响原因是投饲率过高使得鲻长时间处于饱食状态,进一步影响鲻的进食速度,使得饲料在水中浸泡时间长,导致营养成分流失,鱼体吸收消化的蛋白量较少[26]。鲻本身就是高蛋白鱼类,本试验结果证明,可通过适当改变投饲率控制鲻体脂含量,进而改善鱼肉品质。灰分含量是测定水生生物体内无机物含量是否符合正常的指标,本试验结果显示,投饲率对鲻鱼体灰分含量无显著影响(P>0.05),这种现象同样出现在日本黄姑鱼幼鱼[5]、黄鳝[15]和施氏鲟[27]的研究中。不同投饲率对鲻灰分含量影响不大,各组间参数小范围的浮动可能是由鱼粉矿物质在鱼体内沉积导致。
3.3 投饲率对鲻肝脏消化酶活性的影响
消化酶主要指由消化腺和消化系统分泌的具有消化作用的酶,其活性是反映鱼体消化吸收营养物质能力的指标之一,尤其在鱼类生活史早期阶段,其形态、生理等都会发生很大变化,可通过研究鲻消化酶活性特点,深入了解鲻摄食、消化和吸收的机制[28]。本试验中,在不同投饲率下生长发育的鲻体内肝脏消化酶活性受到的影响也不尽相同。结果显示,投饲率的升高对鲻肝脏中淀粉酶活性无显著影响,而蛋白酶和脂肪酶活性则显著升高。蛋白酶活性随着投饲率的升高呈先升后降的趋势,原因可能是不同生长发育阶段、饵料因素等造成的蛋白酶活性不同。沈文英等[29]在对不同生长阶段的银鲫消化酶的研究中指出,银鲫的胰蛋白酶活性在鱼苗期最低,在亲鱼期可达到最高。本试验中,鱼体摄食量不同,摄食量低的鲻会比摄食量高的营养吸收少、发育晚,进而影响蛋白酶活性。褚志鹏等[21]研究发现,随着投饲率的升高,大杂交鲟幼鱼胃蛋白酶活性有升高的趋势,这与本试验结果相似。淀粉酶活性一般与鱼体对碳水化合物的吸收利用能力有关[15],鲻淀粉酶活性随着投饲率的升高有略微上升的趋势,但差异不显著,说明鲻对饲料中淀粉的消化能力不高,略微差异有可能是由肝脏消化酶之间的相互影响导致[23]。对糖类利用能力低的水产鱼类来说,脂肪的供能作用显得特别重要,脂肪在脂肪酶的作用下分解成脂肪酸和甘油醛混合物,被血管和淋巴管运输至各身体组织,给鱼体生长提供必需营养物质[30]。本试验中,投饲率为10%的试验组脂肪酶活性显著高于其他组,原因可能是在低投饲率下,摄食的营养物质不足使得消化腺发育小,分泌的消化酶不足、活性低。同时,投饲率升高,鱼体也会适应性分泌较多的消化酶和提高消化酶活性来充分吸收分解食物中营养物质[9],促进生长发育。本试验结果显示,鲻肝脏脂肪酶活性在投饲率为10%试验组最大,说明此时饲料脂肪含量多,脂肪酶与底物作用面积大,更好地刺激脂肪酶的分泌,促进酶活性提高[28]。当投饲率升至15%时,脂肪酶活性反而下降,说明此时肝脏可能为负荷状态,机体在超过一定的承受范围后受损,脂肪酶活性受到抑制。
3.4 投饲率对鲻肝脏免疫酶活性的影响
免疫酶活性能够在一定程度上反映鱼类免疫、代谢功能的状况[21]。本试验结果表明,不同投饲率对鲻肝脏中过氧化氢酶、超氧化物歧化酶和溶菌酶活性有显著影响(P<0.05)。随着投饲率的增加,过氧化氢酶活性呈持续上升趋势,在投饲率为15%时达到最大值;超氧化物歧化酶和溶菌酶活性呈先升后降的趋势,在投饲率为10%时达到最大值。有研究表明:随着投饲率的增加,鱼类的免疫功能也随之增加,但投饲率过低或过高均有可能影响鱼类的肝脏免疫酶活性;投饲率过低导致鱼类处在较低的代谢水平下,从而使得各免疫酶活性较低;投饲率过高,则会使鱼类的肝脏受损,从而导致代谢紊乱[17,21,30]。这些可能是造成本试验鲻肝脏中各免疫酶在低投饲率和高投饲率的试验组活性均较低的原因。
4 结 论
综合生长速度、饲料利用率、鱼体成分、消化酶脂和免疫酶活性等指标,在水温26~30 ℃的条件下,鲻幼鱼体质量为37.67~133.22 g时,确定养殖最适投饲水平为10%。
猜你喜欢
中老年保健(2022年4期)2022-11-25 14:45:02
肝博士(2022年3期)2022-06-30 02:49:00
当代水产(2021年9期)2021-12-02 01:35:10
当代水产(2021年8期)2021-11-04 08:49:02
水电与抽水蓄能(2021年2期)2021-05-14 01:42:26
农业工程学报(2021年4期)2021-05-09 08:23:58
医学前沿(2021年18期)2021-04-14 02:53:06
家庭医学(下半月)(2020年4期)2020-05-30 12:42:52
农业工程学报(2020年6期)2020-05-19 07:46:38
安徽医科大学学报(2016年12期)2017-01-15 14:21:49
