
2019年舟山近海浮游动物群落结构变化与水团的关系
2022-09-21 09:02胡亚丽许永久王慧君朱文斌张洪亮蒋日进吴丽航徐纪晓杨丽萍姜淇俊李立华
水产科学 2022年5期
胡亚丽,梁 君,许永久,郑 基,王慧君,朱文斌,郭 爱,张洪亮,蒋日进,吴丽航,徐纪晓,杨丽萍,姜淇俊,李立华
( 1.浙江海洋大学 水产学院,浙江 舟山 316022;2.浙江省海洋水产研究所,浙江 舟山 316021 )
浮游动物是海洋次级生产者中的主要类群,在生态系统中起着非常重要的调控作用[1-2]。舟山近海海域位于长江口以南、杭州湾以东,受长江、钱塘江两大江河冲淡水以及来自外海的台湾暖流的强烈影响,水域环境复杂多变,水团的交汇带来了大量的营养盐,不仅为该海域浮游动物的生长提供了充足的物质基础,而且水团交汇引起的海洋锋使得锋区浮游动物密度比较高,非锋区浮游动物密度比较低,也会引起该海域浮游动物群落结构产生明显的异质性[3-4]。因此,研究浮游动物多样性、群落结构、季节动态变化以及与环境因子的关系对浮游动物群落结构动态变化机制有重要的参考意义,也可为海洋生态系统修复提供科学依据。
目前,关于我国近海浮游动物群落结构与时空分布已有诸多报道[5-12],这些研究大多集中在浮游动物的种类组成、数量空间分布、生态特征及影响因素等方面,而关于舟山近海水团季节变化对浮游动物群落结构影响等方面的研究较少。笔者根据2019年春季(4月)和秋季(11月)在舟山附近海域对生态系统开展综合调查所获得的浮游动物的资料,研究浮游动物的群落结构、优势种丰度、多样性指数的季节变化及其与水团的关系,旨在为生态系统的修复提供基础资料[13]。
1 材料与方法
1.1 采样设计与方法
2019年4月11—23日(春季)和2019年11月5—21日(秋季),搭载“浙普渔43019”开展两个航次的浮游生物生态调查,调查区域为N 29°~32°,E 121°~124°。该海域为传统渔业作业海域,底层海域作业和捕捞渔具较多,未进行垂直拖网,采取水平拖网的方法进行调查。利用浅水Ⅰ型浮游生物拖网(口径50 cm、筛绢CQ14、网长145 cm、孔径 0.505 mm)在调查区域进行水平拖曳,春季的调查站位为41个,秋季的调查站位为46个(图1)。每站表层水平拖网10 min,这与以往调查浮游动物的方法[14-15]一致。所获样品均经5%甲醛溶液固定保存后,带回实验室内通过显微镜分类、鉴定和计数。每个站位采集的浮游动物均按照统一的单位计算出浮游动物在单位水体中的丰度(个/m3)。水温、盐度使用温盐深仪同步测定,利用水桶采集表层水样,利用尼斯金采水器采集5 m处的水样,两层水样经过whatsman GF/F玻璃纤维滤膜过滤,滤膜用锡箔纸包裹,然后放置于-80 ℃液氮内冷冻保存。叶绿素样品在低温避光的环境下用90%丙酮萃取48 h,采用Trilogy荧光计测定上清液的荧光值,从而得到叶绿素a的质量浓度[5]。浮游动物样品的采集、保存、运输和分析均按照GB/T 12763.6—2007《海洋调查规范 海洋生物调查》中的规定执行。

图1 2019年春季(a)和秋季(b)调查站位空间分布Fig.1 Sampling stations in spring (a) and autumn (b) of 2019代表杭州湾水团(1区),代表舟山本岛上升流水团(2区),代表舟山近海水团(3区).The water mass in the Hangzhou Bay(AreaⅠ) is marked by ,upwelling water mass of the Zhoushan Islands (Area Ⅱ) is marked by ,and the offshore water mass of the Zhoushan(Area Ⅲ) is marked by .
1.2 数据处理
1.2.1 浮游动物类群划分
鉴定的浮游动物类群主要划分为:桡足类、水母类、磷虾类、糠虾类、毛颚类、十足类、浮游幼体,将出现频率均低于5%的枝角类、浮游软体动物、介形类、海樽类、端足类、多毛类等统归为其他类进行研究。浮游动物的鉴定方法参考文献[16],因为浮游动物数据的差异性比较大,为减少极值对数据的影响,将所有站位每种浮游动物数量进行标准化处理,公式[17]如下:
Yi=log (Xi+1)
(1)
式中,Xi为原始丰度,Yi为转换后数值。
1.2.2 群落结构参数计算公式
(1)浮游动物的丰度(N)和优势度(Y)
丰度利用采样时的滤水体积来计算,以每立方米水体中的数量(个/m3)来表示。
丰度(N)计算公式为:
N=n/V
(2)
式中,V为采集浮游动物样品中过滤海水体积(m3),n为样品鉴定计数获得的浮游动物数量(个)。
公式2中滤水体积(V)计算方法为:
V=A×S
=π(0.25)2×926
≈181.7275
其中,A为网口面积,S为拖行距离。拖行距离926 m通过拖行速度乘以拖行时间来计算,拖行速度v=3 kn=5556 m/h,拖行时间t=1/6 h,即S=v×t=926 m。
优势度(Y)的计算公式为:
Y=(Ni/N)fi
(3)
式中,N为各采样站位所有种类的丰度,Ni为第i种的丰度,fi为该种在各站位出现的频率。
以Y≥0.02为优势种的划分标准[18-19]。
(2)多样性指数计算
香农—维纳多样性指数(H′):
(4)
式中,S为浮游动物的总物种数量,Pi为此物种个体数占总个体数的比例。
Margalef物种丰富度指数(D):
D=(S-1)/lnn
(5)
式中,S为浮游动物的总物种数量,n为浮游动物总个体数量。
Pielou物种均匀度指数(J):
选取我院收治的38例患者都是疑似结核性胸膜炎的患者,他们在医生的指导之下都自愿接受了B超和CT的检查,其中有18位女士和20位男士,研究人员在本次对结核性胸膜炎患者的B超影像学的表现极其诊断价值中将这些患者当作研究的对象进行观察。研究对象的选择主要有以下几个标准:首先已经确诊患上了结核性胸膜炎,其次在进行研究以及资料的公开之前都将整个研究过程告知的患者并且经过了结核性胸膜炎患者的同意,这些患者年龄20~80岁,患者在知道所有事项的情况下资源接受超声和CT等有助于研究的检查,结核性胸膜炎患者的临床资料都是十分完善的,最后本次研究经过了相关国家机构以及医院的批准在公开的情况下进行。
J=H′/log2S
(6)
式中,S为浮游动物的总物种数量。
环境因子空间分布图、浮游动物类群丰度分布图和多样性指数的空间分布图等均使用Sufer 13软件绘制。
1.3 统计分析
利用SPSS 19.0软件中的系统聚类分析方法,根据环境数据中的海水表层温度和海水表层盐度,对空间站位聚类;利用PRIMER 5.0和EXCEL 2010进行统计分析,得到浮游动物的丰度(N)、种类数(S)、物种丰富度指数(D)、物种均匀度指数(J)和香农—维纳多样性指数(H′);对春、秋季的多样性数据进行独立样本t检验;采用Sufer 13软件绘制物种空间分布图。利用PRIMER 5.0软件中的相似性百分比分析方法,计算不同水团的浮游动物种类组成差异的贡献种和贡献率。采用相似性分析(ANOSIM)检验不同水团群落结构间差异的显著性[20]。优势种与环境因子的关系通过CANOCO 5软件中的典范对应分析[20]来研究。其显著性根据蒙特卡罗检验确定,当变量对应轴之间的相关系数|R|≥0.4时,即认为该变量具有重要意义[21]。
2 结 果
2.1 环境因子分析
2019年春季海水表层温度为13.4~18.3 ℃,平均为15.61 ℃;海水表层盐度为8.6~32.3,平均为24.4;叶绿素a质量浓度为0.08~3.64 mg/m3,平均为0.75 mg/m3。2019年秋季海水表层温度为18.7~22.4 ℃,平均为20.7 ℃;海水表层盐度为12.3~33.7,平均为26.5;叶绿素a质量浓度为0~0.75 mg/m3,平均为0.15 mg/m3。2019年春季和秋季的温度、盐度和叶绿素a质量浓度均呈现出近海高、近岸低的趋势;从季节差异来看,2019年春季的海水表层温度和海水表层盐度均明显低于秋季,而叶绿素a质量浓度高于秋季(图2)。
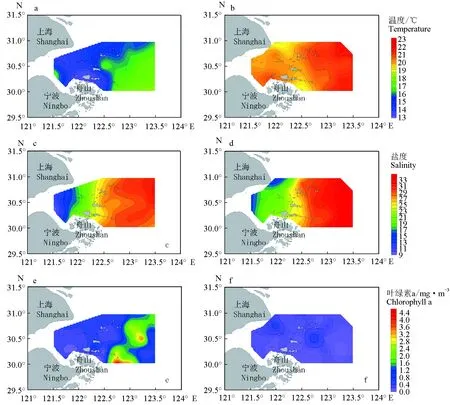
图2 2019年春季和秋季舟山近海海域海水表层温度、海水表层盐度、叶绿素a质量浓度分布与季节变化Fig.2 Distribution and seasonal changes in surface temperature,surface salinity,and chlorophyll a concentration in the Zhoushan offshore waters in spring and autumn of 2019a.春季海水表层温度;b.秋季海水表层温度;c.春季海水表层盐度;d.秋季海水表层盐度;e.春季叶绿素a;f.秋季叶绿素a.a.sea surface temperature in spring;b.sea surface temperature in autumn;c.sea surface salinity in spring;d.sea surface salinity in autumn;e.sea surface chlorophyll a concentration in spring;f.sea surface chlorophyll a concentration in autumn.
对海水表层温度和海水表层盐度进行系统聚类,根据分类结果(图3),将调查区域分为:杭州湾内水团(1区)、舟山本岛上升流水团(2区)和舟山近海水团(3区)3个水团。杭州湾内水团的性质春季为低温低盐水,秋季为高温低盐水;舟山本岛上升流水团的性质春季为低温低盐水,秋季为高温低盐水;舟山近海水团的性质春季和秋季皆为高温高盐水[20]。
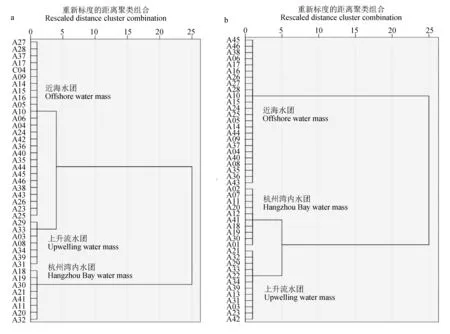
图3 2019年春季(a)与秋季(b)水团聚类Fig.3 Water mass group clustering of sampling stations in spring (a) and autumn (b) of 2019
2.2 舟山近海浮游动物群落结构
2019年春季共鉴定出浮游动物31种,其中:10种属于桡足类,桡足类浮游动物物种数量占浮游动物总物种数量的32.3%,为第一优势类群;4种属于水母类,水母类浮游动物物种数量占浮游动物总物种数量的12.9%,为第二优势类群(表1)。2019年秋季鉴定的浮游动物物种数量明显增多,共63种,其中:23种属于桡足类,桡足类浮游动物物种数量占浮游动物总物种数量的36.5%,为第一优势种类群;17种属于水母类,水母类浮游动物物种数量占浮游动物总物种数量的27.0%,为第二优势种类群。

表1 舟山近海海域浮游动物种类组成Tab.1 Species composition of zooplankton in Zhoushan offshore waters
2.3 舟山近海优势种空间分布

表2 2019年春季和秋季浮游动物的优势种和优势度Tab.2 Dominant species and their dominance in spring and autum of 2019
2019年春季优势种中华哲水蚤丰度在上升流水团的近岸海域出现高值(图4a),丰度为0~47.61个/m3,平均为5.12 个/m3;捷氏歪水蚤多分布在杭州湾内水团(图4b),丰度为0~72.90 个/m3,平均为3.82个/m3;五角水母多分布在上升流水团的近岸海域(图4c),丰度为0~14.09 个/m3,平均为1.05 个/m3。秋季与春季相比,由于温度、盐度和叶绿素a等环境因子的变化,调查海域内的优势种也发生改变。秋季优势种哲水蚤幼体多分布在上升流水团和舟山近海海域(图4d),丰度为0~15.74 个/m3,平均为1.37 个/m3;中华假磷虾丰度在舟山近海海域出现较高值(图4e),丰度为0~11.35个/m3,平均为1.02 个/m3;猛水蚤目多分布在舟山近海海域,且分布比较广泛(图4f),丰度为0~8.94 个/m3,平均为0.89 个/m3。2019年春季和秋季浮游动物总丰度时空分布特征为30.5 °N附近海域值较高,春季浮游动物的丰度与秋季浮游动物的丰度有显著差异,且春季丰度显著大于秋季(P<0.05)。

图4 2019年春季、秋季浮游动物优势种丰度分布(个/m3)Fig.4 Distribution of dominant zooplankton species abundance in spring and autumn of 2019a.中华哲水蚤;b.捷氏歪水蚤;c.五角水母;d.哲水蚤幼体;e.中华假磷虾;f.猛水蚤目;g.2019年春季物种总丰度;h.2019年秋季物种总丰度.a.C. sinicus;b.T. derjugini;c. M. atlantica;d.C. sinicus larva;e.P. sinica;f.Harpacticoida;g.total species abundance in spring 2019;h.total species abundance in autumn 2019.
2.4 春、秋季浮游动物的多样性变化
2019年春季各站位浮游动物种类数为3~14,平均值7.32;物种丰富度指数为0.27~1.90,平均值0.94;物种均匀度指数为0.24~0.82,平均值0.44;香农—维纳多样性指数为0.04~2.46,平均值1.26。2019年秋季各站位浮游动物种类数为5~33,平均值17.99;物种丰富度指数为1.20~4.12,平均值2.70;物种均匀度指数为0.20~0.98,平均值0.61;香农—维纳多样性指数为0.85~3.71,平均值2.46。春季和秋季的浮游动物的种类数、物种丰富度指数、物种均匀度指数和香农—维纳多样性指数均有显著差异,且秋季的多样性显著大于春季(P<0.01,图5)。

图5 2019年春季、秋季浮游动物多样性空间分布Fig.5 Spatial distribution of zooplankton diversity in spring and autumn 2019a.春季种类数;b.秋季种类数;c.春季物种丰富度指数;d.秋季物种丰富度指数;e.春季物种均匀度指数;f.秋季物种均匀度指数;g.春季香农—维纳多样性指数;h.秋季香农—维纳多样性指数.a.number of species in spring;b.number of species in autumn;c.species richness index in spring;d.species richness index in autumn;e.species evenness index in spring;f.species evenness index in autumn;g.Shannon-Wiener′s diversity index in spring;h.Shannon-Wiener′s diversity index in autumn.
2.5 舟山近海浮游动物群落结构、优势种、多样性季节变化及与水团的关系
3种水团对2019年浮游动物群落结构的空间分布格局有显著的影响(ANOSIM检验,P<0.05)。2019年春季,1区和2区、2区和3区以及1区和3区平均相异性分别为65.6%、61.4%和82.5%;引起1区和2区、2区和3区以及1区和3区差异的主要贡献种为捷氏歪水蚤、夜光藻、捷氏歪水蚤(贡献率分别为20.1%、21.1%、22.08%)。2019年秋季,1区和2区、2区和3区以及1区和3区平均相异性分别为62.3%、69.6%和77.02%;引起1区和2区、2区和3区以及1区和3区差异的主要贡献种为中型莹虾、拿卡箭虫、拿卡箭虫(贡献率分别为6.2%、6.5%、7.7%)。季节变化对3种水团的浮游动物群落结构均有显著影响(ANOSIM检验,P<0.05),1区、2区、3区,春季和秋季的平均相异性分别为71.04%、75.71%、91.70%。引起2019年春、秋季相对应的3种水团差异的主要贡献种为捷氏歪水蚤、中华哲水蚤和夜光藻(贡献率分别为10.87%、11.10%和10.94%)。
2019年春季和秋季各优势种丰度与环境变量的典范对应分析(CCA)结果显示(图6),前4轴与环境变量之间的相关性显著(F=15.6,P=0.002),CCA1轴的特征值为0.390,CCA2轴的特征值为0.248,物种—环境之间表现出较高的相关性,CCA1和CCA2解释了物种累计方差百分比的58.85%和物种—环境累计方差总百分比的96.35%,海水表层温度、海水表层盐度和海水底层温度聚集在CCA1的负轴,与CCA1负相关。CCA1轴主要反映了季节的差异,CCA2轴主要反映了近岸和近海的差异。中华假磷虾聚集在CCA1的负轴,与海水底层温度呈现明显的正相关性,说明海水底层温度是影响其分布水域的主要因素。中华哲水蚤聚集在CCA1轴的正轴,与海水表层叶绿素呈现出明显的正相关性,与海水表层温度和海水底层温度呈现出明显的负相关性。除此之外,中华假磷虾聚集在近海海域,说明暖温环境是影响其分布的主要因素。哲水蚤幼体则聚集在近岸海域,这些分隔模式可能反映了浮游动物对栖息地类型的偏好或者其生态习性的适应性。

图6 浮游动物优势种与环境因子典范对应分析Fig.6 Correspondence analysis between dominant zooplankton species and environmental factors黄色代表春季站位,黑色代表秋季站位;SST代表海水表层温度,SSS代表海水表层盐度,SSC代表海水表层叶绿素a,SBT代表海水底层温度,SBS代表海水底层盐度;31.46%和51.50%表示CCA1和CCA2共解释了浮游动物物种变异的51.50%;下同.Yellow color stands for data in spring and black color stands for data in autumn;SST stands for sea surface temperature,SSS stands for sea surface salinity,SSC stands for sea surface chlorophyll a,SBT stands for sea bottom temperature,and SBS stands for sea bottom salinity;31.46% and 51.50% indicate that CCA1 and CCA2 explain 51.50% of the zooplankton species variation;et sequentia.
由多样性与环境因子的典范对应分析结果(图7)可见,前4轴与环境变量之间的相关性显著(F=7.0,P=0.002),CCA1和CCA2解释了物种累计方差百分比的51.91%和物种—环境累计方差总百分比的99.67%,海水表层温度、海水底层温度、海水表层盐度、海水底层盐度和海水表层叶绿素a聚集在CCA2的正轴,与CCA2正相关。CCA1轴主要反映了季节的差异,CCA2轴主要反映了近岸和近海的差异。由图7可见,丰富度和海水表层温度、海水底层温度呈现出明显的正相关性,种类数和海水表层叶绿素呈现出明显的正相关性,多样性、均匀度和海水表层盐度、海水底层盐度呈现出明显的负相关性。因此,温度是影响春、秋季浮游动物多样性差异的主要环境因子,这与以往的研究结果相一致[22]。
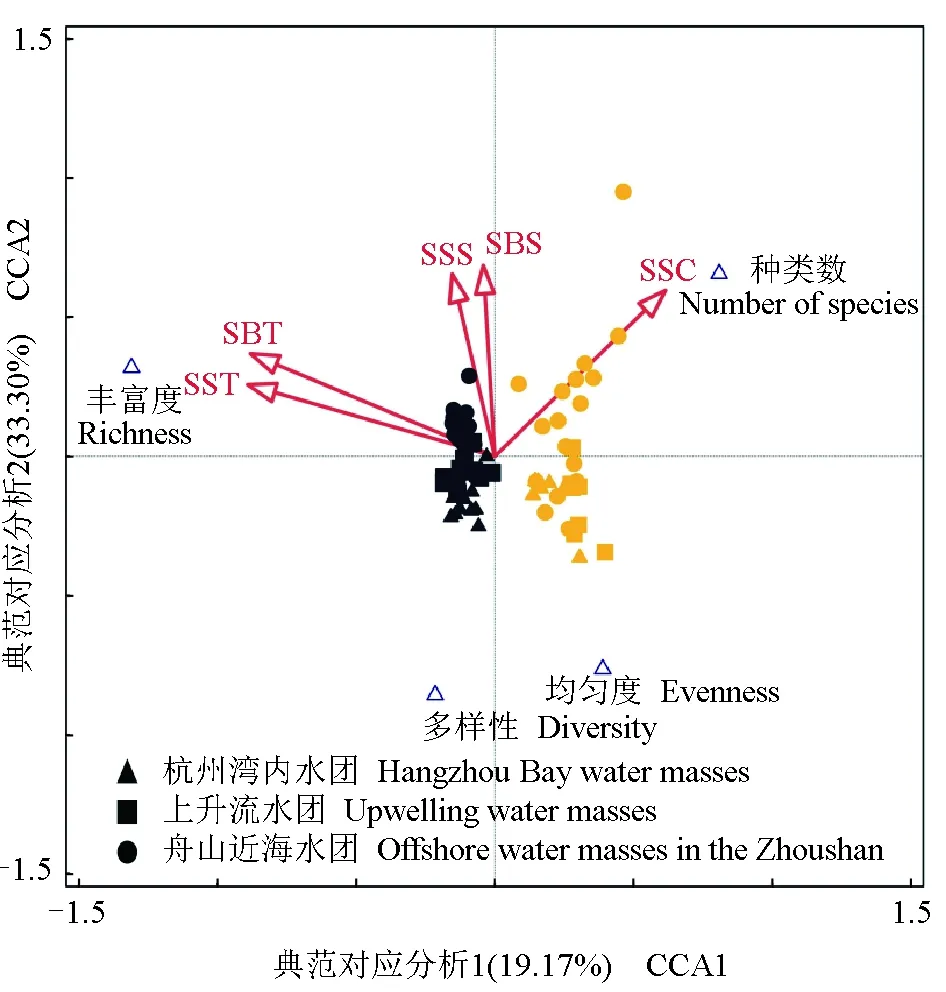
图7 浮游动物多样性与环境因子典范对应分析Fig.7 Correspondence analysis between zooplankton diversity and environmental factors19.17%和33.30%表示CCA1和CCA2共解释了浮游动物物种变异的33.30%.19.17% and 33.30% indicate that CCA1 and CCA2 explain 33.3% of the zooplankton species variation.
3 讨 论
3.1 春、秋季浮游动物优势种的空间分布、丰度的季节变化与水团的关系
笔者发现,舟山近海海域浮游动物优势种的季节更替较为明显。2019年春季(4月)浮游动物的优势种为中华哲水蚤、捷氏歪水蚤和五角水母,中华哲水蚤作为春季浮游动物的第1优势种,遍布整个调查区域,在舟山近岸上升流水团的丰度最高,这主要因为春季台湾暖流对调查区域的影响还较弱[23],近岸海域整体温度不高,3种水团的平均海水表层温度为14.1~16.5 ℃,该温度范围较适宜中华哲水蚤的生长和繁殖[24],这与于雯雯等[19,25]在浙江沿岸海域及长江口附近的春季调查结果一致。五角水母多分布在杭州湾水团,由于其为近岸低盐暖温指示种[26],而调查区域内春季杭州湾的水团的平均海水表层温度为14.8 ℃,平均海水表层盐度为13.4,较适宜五角水母的生存。其他生态类群的分布也体现出环境对浮游动物空间分布的影响。春季各水团浮游动物丰度由低到高依次为舟山近海水团(平均8.32个/m3)、上升流水团(平均19.46 个/m3)和杭州湾水团(平均21.11 个/m3),浮游动
物丰度从近岸到近海呈现出不断减少的趋势,这与以往的研究[7-9]结果存在差异。笔者猜测,造成这一差异的主要原因可能是2019年春季舟山近海发生了较强的浮游植物藻华,大量的藻类在舟山近海叶绿素浓度较高的区域聚集[20],侵占了浮游动物原本的生存空间[27-28],使得较多的浮游动物在杭州湾水团和舟山本岛上升流水团聚集[20],因此,杭州湾水团和上升流水团的浮游动物丰度比舟山近海水团的浮游动物丰度高,也说明藻类的暴发会影响或改变浮游动物的密度和群落结构[29],最终形成2019年春季浮游动物的空间分布格局。另外,舟山近海夜光藻和杭州湾水团中华哲水蚤的大量生存,也一定程度上增加了春季浮游动物的总丰度,因此,叶绿素a和海水表层温度是影响春季浮游生物总丰度的主要环境因子。
秋季(11月)的优势种和春季的优势种有很大不同,主要为哲水蚤幼体、中华假磷虾和猛水蚤目,3种水团内的浮游动物的种类和数量都有很大的差别,浮游动物丰度由低到高依次为杭州湾水团(5.55 个/m3)、上升流水团(5.70 个/m3)和舟山近海水团(7.92个/m3)。浮游动物的空间分布呈现出近岸低、近海高的趋势,这与以往的研究结果一致[12]。从杭州湾水团到上升流水团再到舟山近海水团,盐度呈现出不断增高的梯度变化,浮游动物的丰度也随盐度逐渐上升,近岸海域盐度相对较低,哲水蚤幼体(广温广盐种)和猛水蚤目多聚集在近岸海域,这与以往的研究结果相一致[30-32],近海海域的盐度相对较高,则中华假磷虾(近海物种)等数量较高。因此,盐度是影响秋季浮游动物空间分布的主要环境因子[2,20]。另外,秋季台湾暖流、黑潮等外海暖流势力仍维持较强盛地位,受其影响的海域,浮游动物群落种暖水种和外海种(水母类)仍占有一定的比例[33]。
3.2 春、秋季浮游动物多样性的季节变化与水团的关系
春季海水表层温度低,海水表层盐度低,叶绿素a含量高,浮游动物总丰度大,多样性指数较低;秋季海水表层温度高,海水表层盐度高,叶绿素a含量低,浮游动物总丰度小,多样性指数较高。春季浮游动物的丰度高于秋季浮游动物的丰度,春季浮游动物的多样性小于秋季浮游动物的多样性,这与邵倩文等[4,22]的研究结果相一致。一个区域内群落结构的稳定性与该区域内浮游动物优势种的种类数和数量有很大关系[34],通常优势种的种类数越多且优势度的值越小,说明群落结构越复杂、稳定[35]。本次调查结果显示,春季的浮游动物优势种数目较少(4种)但优势度较高,秋季优势种数目较多(8种)但优势度较低,表明在本次调查区域内,秋季浮游动物群落结构较为稳定,这也在一定程度上说明同一水体中不同季节环境因子的状况会有所不同,且环境因子也一定程度上决定了浮游动物的群落结构特征。
本研究结果显示,春季和秋季浮游动物种类数、物种丰富度指数、物种均匀性指数和香农—维纳多样性指数有显著的差异。秋季的海水表层温度(平均20.7 ℃)相比春季(平均15.6 ℃)较高,研究海域水温的升高有助于浮游动物的生长和繁殖[4],因此秋季的温度较适宜浮游动物的生长,浮游动物种类较多,这也一定程度上增加了浮游动物的多样性,与殷晓龙等[36]的研究结果一致。就该研究区域而言,在N 30.5°附近海域为浮游动物种类数、物种丰富度指数、物种均匀度指数和香农—维纳多样性指数的高值区,表明该区域的浮游生物群落机构复杂稳定,组成丰富。主要原因可能是:N 30.5°处于长江口与舟山渔场,属于亚热带向温带过渡区域,首先生物有充足的饵料,长江径流为该地区带来的丰富物质,为生物的生长提供大量的物质基础;其次生物种类多,台湾暖流为该海域输送了大量的生物种类,集合了丰富的生态类群。因此N 30.5°上的几个站位浮游动物平均种类数较高,这与杨杰青等[24]的研究结果一致。由此可见,温度可能是影响春秋季节浮游生物多样性的主要环境因子。另外,叶绿素a、盐度、饵料等也会影响浮游动物的多样性。
4 结 论
根据2019年春季(4月)和秋季(11月)的浮游动物丰度、优势种、多样性和环境因子等数据分析发现,舟山近海春季和秋季的浮游动物群落结构和环境因子有明显的差异,并且不同水团对浮游动物群落结构优势种分布格局影响显著。盐度是影响浮游动物空间分布差异的主要原因,温度是影响浮游动物季节差异的主要原因。另外,高含量的叶绿素a会给浮游动物提供丰富的饵料,增加浮游动物丰度。
致 谢
对“浙普渔43019”号全体船员在出海采样过程中的帮助,及A17海洋资源与环境班级董军、官道伊等同学在部分实验中的付出,谨致谢忱。
猜你喜欢
河北渔业(2022年10期)2022-10-15
当代水产(2022年8期)2022-09-20
中国海洋大学学报(自然科学版)(2022年9期)2022-09-05
中国音乐学(2022年2期)2022-08-10
昆明医科大学学报(2022年2期)2022-03-29
文萃报·周二版(2022年10期)2022-03-19
学校教育研究(2020年7期)2020-04-09
山花(2019年8期)2019-08-16
华东师范大学学报(自然科学版)(2019年2期)2019-06-11
河北渔业(2014年3期)2014-07-08
