
拟南芥LecRK Ⅳ.3胞内激酶域的原核表达、纯化及鉴定
2022-09-20 02:18:02赵记龙许大金任旭萌张胜伟
河北师范大学学报(自然科学版) 2022年5期
赵记龙,许大金, 任旭萌, 张胜伟
(河北师范大学 生命科学学院 分子细胞生物学教育部重点实验室,河北 石家庄 050024)
位于植物细胞膜表面的类受体激酶(receptor-like kinases,RLKs)在介导外界信号向细胞内传递过程中发挥着重要作用.典型的RLKs由胞外域,跨膜结构域和胞内激酶结构域组成.一般认为当胞外域与特异的配基结合后,其激酶活性会被激活或抑制,胞内激酶结构域与下游蛋白相互作用并启动其生化反应(如磷酸化),调控下游基因表达进行信号输出,从而将外界信号传递到胞内,实现对环境的快速变化作出反应[1-2].因此对RLKs胞内激酶域活性的研究成为了分析其生物学功能及鉴定下游底物分子的基础.
凝集素样类受体激酶(lectin-receptor like kinases,LecRKs)最初在拟南芥中被发现,因胞外域含有一个保守的凝集素结构域而得名,是仅次于富含亮氨酸重复序列类受体激酶(leucine-rich repeat receptor-like kinases,LRR-RLKs)的第二大亚家族,其在水稻中有超过250个成员,在拟南芥中有超过75个成员[3-4].由于在动物和酵母中均未鉴定到此类蛋白,所以推测它们是植物中所特有的一类RLKs.
与LRR-RLKs相比,目前关于植物的LecRKs报道较少,仅对少部分基因的功能进行了研究.由于LecRKs的胞外域与动物的凝集素蛋白相似,科学家推测其可能具有识别和转糖的功能[4-5].近些年的研究表明,LecRKs在植物抵御生物和非生物胁迫中发挥重要作用[6-8].例如,拟南芥LecRK Ⅴ.5,LecRK Ⅸ.1和LecRK Ⅸ.2能增强对疫霉的抗性[9];LecRK Ⅵ.2是气孔先天免疫的正调节因子[10];LecRK Ⅸ.2或LecRK Ⅳ.3能增强对丁香假单胞菌或葡萄孢菌的抗性;SDS2与RLCK118/176相互作用,通过磷酸化调控NADPH氧化酶OsRbohB,刺激活性氧(Reactive oxygen species,ROS)产生,从而正向调控水稻免疫反应[11-12];病原体入侵时会造成胞内ATP释放到胞外,被受体LecRK1.9/DORN1感知,从而磷酸化RBOHD,导致ROS爆发,关闭气孔[13].笔者所在课题组前期研究也发现,LecRK Ⅳ.1/SIT1参与盐胁迫的早期应答过程.SIT1激酶活性被盐激活后,通过激活下游的MAPK3/6,进而促进乙烯的合成和ROS的产生,引起植物对盐的敏感反应[14-15];最新研究表明番茄SlLecRK1.9通过诱导ERFs表达提高番茄对尖镰孢番茄颈腐根腐病专化型病原菌的抗性[16].
LecRK Ⅳ.3属于拟南芥LecRLKs家族中的一员,其编码序列全长2 025 bp,编码674个氨基酸,其在拟南芥中的分子功能未有详细报道.笔者初步的研究数据表明,LecRK Ⅳ.3在植物盐胁迫、紫外线胁迫的应答过程中发挥着重要作用(数据未发表).因此,本研究拟对LecRKⅣ.3基因胞内域(intracellular domain,ID)进行体外原核表达纯化,获得正确的LecRK Ⅳ.3ID蛋白,并验证其具有激酶活性,为进一步揭示LecRK Ⅳ.3的生物学功能及其所调控的信号通路奠定基础.
1 材料与方法
1.1 实验材料
大肠杆菌克隆感受态细胞DH5α,表达感受态细胞BL21,Rosetta,BL21(AI),Rosetta(DE3)pLysS,OverExpress C43(DE3)和pET-30a载体;BamH Ⅰ和HindⅢ限制性内切酶、T4 DNA连接酶、预染蛋白质分子量标准(10~170 ku)、化学发光的底物(Dura)(美国:Thermo Fisher Scientific);基因克隆Pfu高保真酶(中国:北京全式金生物);DNA分子量标准、PCR产物胶回收和质粒快速提取试剂盒(中国:天根生物);一步重组试剂盒(中国:诺维赞生物);菌落PCR鉴定Extaq酶(日本:Takara);His纯化Ni柱(美国:GE);IPTG、溶菌酶(美国:Sigma);辣根过氧化物酶标记的His-HRP抗体(中国:康为世纪);放射性同位素32P(中国:同辐).
引物合成和测序均由Invitrogen公司完成.
1.2 试验方法
1.2.1LecRKIV.3ID基因克隆与His-LecRKⅣ.3ID原核表达载体的构建
用TRIzol法从生长7 d的拟南芥幼苗中提取植物总RNA,通过反转录获得cDNA.设计LecRKⅣ.3ID特异引物ID-F/ID-R,利用Pfu高保真DNA聚合酶进行扩增获得片段长度为1 059 bpLecRKⅣ.3-ID的PCR产物.产物胶回收后经一步重组法连入预先BamH Ⅰ和HindⅢ酶切过的pET-30 a空载体中,转化大肠杆菌克隆感受态细胞DH5α.挑取单克隆菌落,利用基因ID-F/载体T7-term引物进行菌落PCR鉴定,阳性克隆进行测序,最终获得序列正确融合的His-LecRKⅣ.3ID表达载体.
1.2.2His-LecRKⅣ.3的转化和蛋白纯化
将1 μL融合成功的His-LecRKⅣ.3ID分别转化至不同的表达感受态细胞宿主中(见表1).根据在不同表达感受态细胞中的转化效率和生长情况,探究His-LecRK Ⅳ.3ID蛋白激酶表达的最适宿主.最后挑取最适宿主进行下一步实验.

表1 常用大肠杆菌表达感受态细胞类型及特点Tab.1 E. coli Expression Competent Cell Types and Characteristic
挑取平皿上单克隆菌落,接菌至3 mL液体LB培养基(kan+),37 ℃过夜摇培,次日1∶100(体积比)扩培至OD值=0.8~1.0(λ=600 nm),收集1 mL菌液为诱前菌体.取50 mL诱导后菌液,其中取出1 mL菌液为诱后菌体,其余低温6 000 r/min,离心15 min,收集菌体;加入平衡缓冲液(20 mmol/L Tris-HCl pH=7.4,500 mmol/L NaCl,1 mmol/L PMSF,1 mmol/L MgCl2,溶菌酶 1 mg/mL,蛋白酶抑制剂cocktail,20 mmol/L 咪唑)重悬菌体,移液枪吹吸充分打散,冷室孵育30 min;使用超声波破碎仪,30 %强度、超声3 s、停止8 s进行破碎,至菌液澄清透明,一般不超过5 min;低温14 400 rpm离心10 min,将上清转移至新的离心管中,上清和沉淀分别收集1 mL留样;其中上清中加入100 μL预先平衡的Ni-NTA Agarose beads,冷室垂直摇床上孵育1 h以内;低温1 500 r/min,离心5 min,收集琼脂糖凝胶微球,取出1 mL 流穿液(flow through,FL)留样;富集的琼脂糖凝胶微球中加入洗涤缓冲液(20 mmol/L Tris-HCl pH=7.4,500 mmol/L NaCl,50 mmol/L 咪唑),洗涤3次,每次5 min;将琼脂糖凝胶微球转移至重力柱内,再洗涤2次,低温1 500 r/min离心10 s,去掉残留的洗涤缓冲液;加入100 μL洗脱缓冲液(20 mmol/L Tris-HCl pH=7.4,500 mmol/L NaCl,100 mmol/L 咪唑),孵育5 min;低温1 500 r/min离心1 min,收集蛋白,共洗脱3次,留样Elution1(E1),Elution2(E2),Elution3(E3).
诱前,诱后,上清,沉淀,FL,E1,E2和E3样品分别用20 μL 2× SDS蛋白上样缓冲液重悬,100 ℃水浴10 min,顶速离心,十二烷基硫酸钠聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE)检测蛋白纯化情况.
1.2.3 His-LecRK Ⅳ.3ID融合蛋白的鉴定.
为了验证纯化的蛋白是否为LecRK Ⅳ.3ID,利用Anti-His-HRP抗体通过Western blot免疫印迹检测.将His-LecRK Ⅳ.3ID蛋白进行SDS-PAGE电泳,转膜,5 %脱脂奶粉封闭,孵育Anti-His-HRP,化学发光法检测目的条带.
1.2.4 LecRK Ⅳ.3的激酶活性分析
将0.5 μg His-LecRK Ⅳ.3ID加入激酶反应缓冲液中(50 mmol/L HEPES-KOH pH=7.5,10 mmol/L MgCl2,1 mmol/L DTT,5 mmol/L MnCl2,100 μmol/L ATP,5 μCi同位素标记ATP),30 ℃反应3 h;反应完毕后,加入2×蛋白上样缓冲液,70 ℃温浴15 min,顶速离心,预制胶进行SDS-PAGE;电泳完毕后用考染液拷染3 h以上,再用超纯水脱色7 h 以上,照相留图;胶贴住磷屏放至压片暗盒内,利用Typhone仪器扫描信号.
2 结果与分析
2.1 LecRK Ⅳ.3ID基因克隆及His-LecRK Ⅳ.3ID原核表达载体构建
以拟南芥的cDNA为模版,LecRKⅣ.3 ID-F/ID-R为特异性引物进行PCR反应,产物经1 %琼脂糖凝胶电泳,在UV-B灯下可观察到一条约1k bp的特异高亮条带(图 1a).通过一步重组法将胶回收的目的基因片段与预先用BamH Ⅰ和HindⅢ双酶切后的pET-30a空载体进行连接反应.转化DH5α后进行菌落PCR鉴定,对得到的阳性克隆进行测序,最后获得序列融合成功的His-LecRKⅣ.3ID环状质粒(图 1b,c).
2.2 不同表达感受态细胞宿主对His-LecRK Ⅳ.3ID转化的影响
将融合成功的His-LecRKⅣ.3ID分别转化至不同的表达感受态细胞宿主中.结果发现,转化BL21和Rosetta宿主中的大肠杆菌均不能正常生长,而转化BL21(AI),Rosetta(DE3)pLysS,OverExpress C43(DE3)宿主中的大肠杆菌均可以正常生长.其中,OverExpress C43(DE3)宿主中的大肠杆菌生长菌落数最多,转化效率最高(图2).
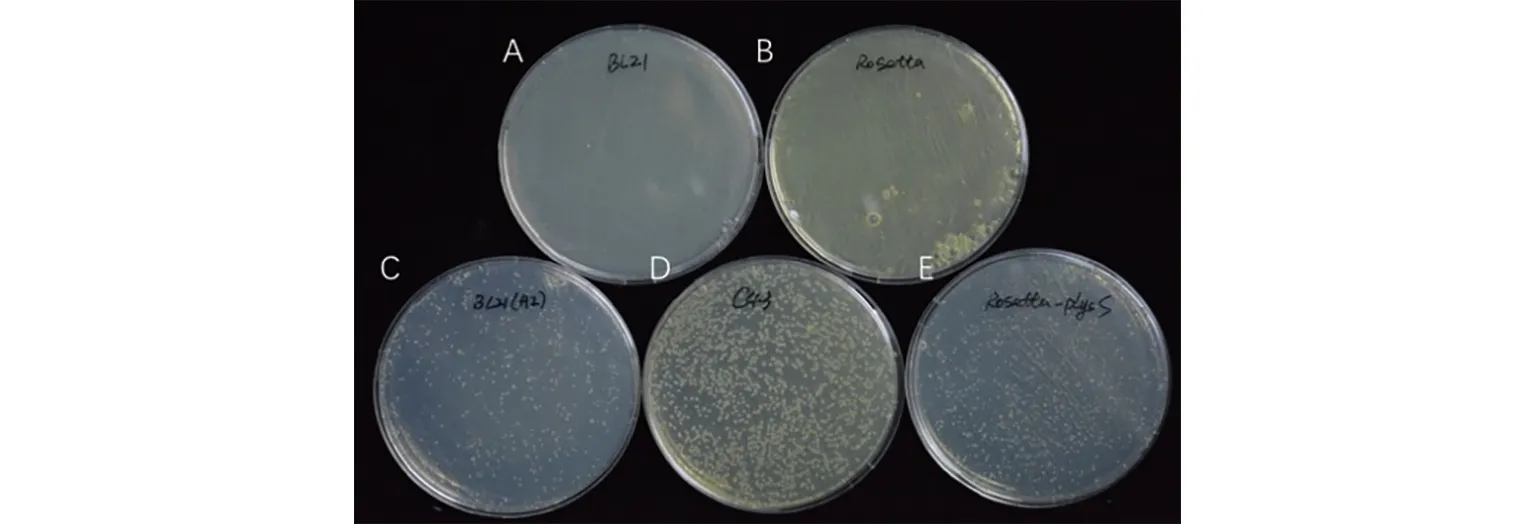
A~E宿主分别是:BL21,Rosetta,BL21(AI),OverExpress C43(DE3),Rosetta(DE3)pLysS.图2 His-LecRK Ⅳ.3ID在不同表达感受态细胞宿主中的生长情况Fig.2 The Growth of His-LecRK Ⅳ.3ID in Different Host Cells
将转化His-LecRKⅣ.3ID的宿主BL21(AI)和OverExpress C43(DE3)菌株分别进行接菌、扩培,使用0.5 mmol/L IPTG 在18 ℃诱导12 h.结果表明,OverExpress C43(DE3)宿主中得到的His-LecRK Ⅳ.3ID融合蛋白主要在可溶性蛋白中表达(图3a),而BL21(AI)宿主中得到的His-LecRK Ⅳ.3ID融合蛋白主要在包涵体中(图3b).
2.3 Western blot免疫印迹鉴定His-LecRK Ⅳ.3ID蛋白
Western blot结果显示,在48 ku分子量位置有一条特异条带,理论分子量与实际分子量相符合,所以纯化获得的蛋白是融合的His-LecRK Ⅳ.3ID蛋白(图4).
2.4 放射性同位素法证实His-LecRK Ⅳ.3ID具有正常的激酶活性
His-LecRK Ⅳ.3ID蛋白通过和放射性32P标记的ATP进行激酶反应,放射性自显影实验发现His-LecRK Ⅳ.3ID能发生自我磷酸化(图5),这说明它具有激酶活性.
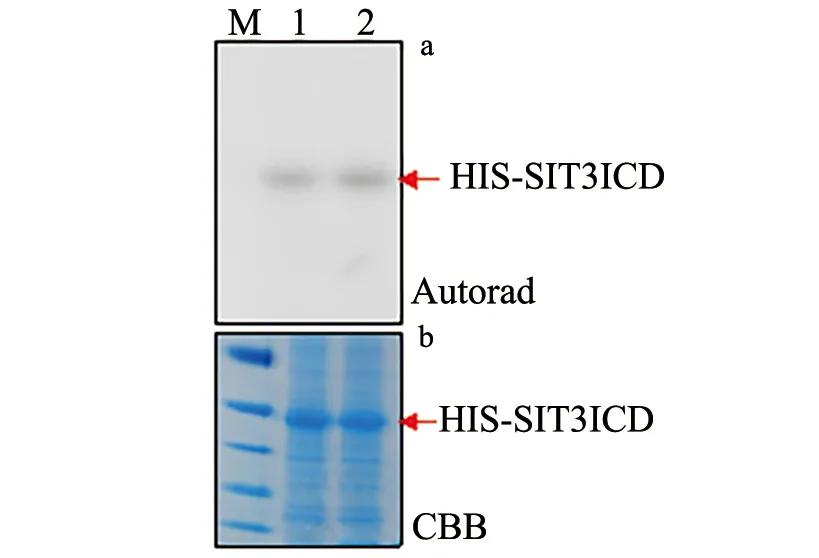
1,2.OverExpress C43(DE3)宿主中纯化His-LecRK Ⅳ.3ID蛋白;M为蛋白分子量指示Maker.a.放射性自显影(Autorad)结果,b.考马斯亮蓝(coomassie brilliant blue,CCB)染色结果.图5 LecRK Ⅳ.3ID具有激酶活性Fig.5 LecRK Ⅳ.3ID Has Kinase Activity
3 讨 论
类受体激酶被认为是调节植物环境适应性的关键信号分子,它能够通过一系列的信号转导来实现对外界环境快速变化作出的反应[1-2].近些年LecRKs家族基因的功能也被越来越多的发现和报道.本课题组研究发现,LecRK Ⅳ.3在拟南芥抗盐胁迫及UV-B胁迫过程中发挥着非常重要的作用,因此成功表达出该蛋白的激酶域并对活性进行初步分析为进一步研究LecRK Ⅳ.3的生物学功能并阐明它介导的信号通路奠定了基础.
蛋白的原核表达是获得蛋白质最常见方法之一[17-18].平常实验中使用较多的标签一般会选择GST,MBP,His等.His标签由于标签小、等电点低、纯化介质成本低而受到科研工作者的青睐[19];而MBP和GST分子量较大,后续Pull-down(拉下)或蛋白质体外结合等实验中容易出现标签本身与潜在蛋白的非特异结合、蛋白融合标签后出现翻译蛋白断裂等问题.作为膜蛋白,类受体激酶的蛋白纯化相对于胞质蛋白比较困难.其一是他一般都具有较强的激酶活性,对表达宿主有毒性,在常规的大肠杆菌表达菌株中转化效率较低,甚至不生长;其二是诱导过程中它特别容易进入包涵体中,不易后续的纯化;其三是其诱后和诱前相比,卡马斯亮蓝染色实验不易观察到其菌体蛋白表达的明显差异,只能通过后续获取超声破碎后的上清和沉淀再进行分析.
本文中,通过转化不同大肠杆菌表达感受态细胞宿主找到了His-LecRK Ⅳ.3ID蛋白纯化的最佳宿主,并在18 ℃低温条件下成功纯化了正确的、有激酶活性的His-LecRK Ⅳ.3ID融合蛋白.这不仅为研究LecRK Ⅳ.3的生物学功能奠定了基础,还为将来纯化具有较强激酶活性的蛋白激酶提供理论参考.在His-LecRK Ⅳ.3ID纯化过程中,发现His标签固有的缺点,由于His标签融合蛋白洗脱需要用到咪唑,而高浓度咪唑容易使蛋白变性,所以蛋白不适合于冷冻保存.笔者通过实验证实,较稳定的蛋白可于4 ℃短暂保存一周,若需要长期保存需要通过透析或脱盐柱去除咪唑,以免影响后续的实验.最后建议蛋白激酶类蛋白纯化要现用现制,以此保证蛋白的正常生物学功能.
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
天津医科大学学报(2021年3期)2021-07-21 09:03:46
世界科学技术-中医药现代化(2021年12期)2021-04-19 12:31:40
河北北方学院学报(自然科学版)(2020年8期)2020-01-18 01:38:48
生命科学研究(2018年1期)2018-05-29 01:12:47
生物工程学报(2017年4期)2017-05-06 07:09:55
上海农业学报(2017年3期)2017-04-10 12:39:14
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
安徽农业科学(2016年17期)2016-08-11 11:22:38
广西医学(2016年11期)2016-02-17 03:34:58
