
刈割和施氮对豆禾混播草地牧草生长特性及品质的影响*
2022-09-20 08:41王辛有曹文侠师尚礼王小军王世林刘婉婷
中国生态农业学报(中英文) 2022年9期
王辛有, 曹文侠**, 师尚礼, 王小军, 王世林, 刘婉婷, 白 洁, 李 文
(1.甘肃农业大学草业学院/草业生态系统教育部重点实验室/中-美草地畜牧业可持续发展研究中心 兰州 730070; 2.青海大学畜牧兽医科学院 西宁 810000)
河西走廊是我国重要的草牧业生产基地, 不合理的利用制度和管理模式导致天然草地退化严重,而干旱缺水的环境条件也使该地区各类人工草地的生产力无法满足畜牧业可持续发展的需求。建植高产优质的放牧型人工草地, 成为发展集约化高效现代草地畜牧业的重要途径。豆科-禾本科混播草地(豆禾混播草地)可在提升草地生产性能的同时,为家畜提供营养均衡的优质混合牧草, 成为建植放牧型人工草地的优先选择。混播草地组分的平衡与群落结构的稳定性是维持豆禾混播草地生产性能与优势的基础, 放牧利用时间、方式和强度, 以及相应的配套管理措施均可能影响混播草地最初设计的草种组分稳定性, 并可能导致草地质量的下降, 使利用年限缩短。因此, 研究放牧型豆禾混播草地的不同利用和配套管理模式对群落稳定性和草地生产性能的影响至关重要。
放牧是草地最直接和最主要的利用方式。合理的放牧利用不但能提高草地生产力和牧草营养品质, 而且能有效维持草地群落稳定性。李振忠等在苇状羊茅(Festuca arundinacea)+红三叶(Trifolium pratense)混播草地上进行的模拟放牧研究表明: 刈前高度(始牧期的草层高度) 30 cm和留茬9 cm能使草地生产力最高; 徐震等认为留茬5.5 cm有利于维持黑麦草(Lolium perenne)+白三叶(Trifolium repens)混播草地的群落稳定性; 刘海泉等则利用模拟放牧的方式对黑麦草+白三叶混播草地的群落结构和生产力进行了综合评价, 认为刈前高度为20 cm、留茬高度为5 cm时, 混播草地群落结构较稳定, 同时草地生产力较高。因此, 不同的刈前高度和刈割强度对草地的影响存在较大差异。近年来, 对豆禾混播草地放牧利用的研究多聚焦于利用强度对草地的影响, 高强度利用会阻碍牧草再生, 加速草地退化, 轻度利用则会浪费优质牧草, 降低草地利用价值。虽然也有学者针对始牧期的选择进行了相关研究, 但以草层高度为衡量标准并搭配适量施肥的研究却十分少见。在混播草地放牧利用过程中, 草层达到何种高度开始利用也同样重要, 刈前高度较低会导致牧草没有足够的营养物质储备, 严重影响其再生; 刈前高度较高则会导致牧草营养价值降低, 不利于家畜采食利用和消化吸收, 使放牧家畜的生产性能下降。此外, 过高的利用强度和频率容易导致土壤有效养分供应不足, 在科学放牧利用的同时对土壤进行适量补氮, 不仅可以提高草地生产性能、维持群落结构稳定, 还可以有效避免养分缺失所导致的草地质量下降。由于在豆禾混播草地中满足禾本科牧草最大生产力所需的氮量远高于豆科牧草向其转入的氮, 对豆禾混播草地进行适量的氮肥补充可以有效促进禾本科牧草的生长。有研究表明, 在放牧条件下对草地施氮有利于牧草地下部分的生长, 能提高牧草水分和营养物质的利用率。因此, 选择合理的刈前高度、刈割强度和配套的施肥管理模式是放牧型豆禾混播草地科学利用的重要基础。
目前有多项研究显示, 刈割是一种快捷有效的模拟放牧技术, 虽不考虑家畜践踏、粪尿归还等多方面的综合作用, 但可以直观有效地研究家畜采食作用对草地生产性能和群落结构的影响。本研究以‘清水’紫花苜蓿(Medicago sativa cv Qingshui)+无芒雀麦(Bromus inermis)+长穗偃麦草(Elytrigia elongate)混播草地为研究对象, 通过刈割模拟放牧利用, 筛选适宜的刈前高度、刈割强度组合以及配套施氮水平, 探讨放牧利用和施氮管理对豆禾混播草地生产性能和群落结构的影响机制, 为豆禾混播草地放牧利用和配套施肥管理提供技术参考依据。
1 材料与方法
1.1 试验地概况
试验地位于河西走廊的甘肃酒泉市肃州区铧尖乡漫水滩村(39°40′12.05″N, 98°49′21.80″E, 海拔1387.20 m), 属大陆性干旱气候, 最高气温38.0 ℃, 最低气温—31.6 ℃,年均温7.9 ℃。年降水量87.70 mm,夏季降水量为48.40 mm, 占全年总降水量的56.70%,年蒸发量为2140 mm, 为降水量的24.40倍。年均日照时数3033.40 h, 平均每天日照时数8.30 h。无霜期短,年平均130 d。试验前土壤总盐含量为7.82 g∙kg, pH为7.83, 有机质含量为3.13 g∙kg, 碱解氮含量为24.87 mg∙kg, 速效磷含量为57.67 mg∙kg, 速效钾含量为118.12 mg∙kg。
1.2 试验设计
根据前期研究结果, 试验对象为2017年4月以刈牧兼用型‘清水’紫花苜蓿、无芒雀麦和长穗偃麦草1∶1∶1建植的豆禾混播草地, 混播方式为同行条播, 行间距20 cm。4月上旬返青, 10月中旬停止生长, 传统方式每年可在6月上旬、7月下旬及9月下旬各刈割1次, 全年可刈割3茬。全生育期保持充分灌水,灌溉方式为喷灌。‘清水’紫花苜蓿草种由甘肃农业大学草业学院提供, 无芒雀麦和长穗偃麦草草种由北京正道种业有限公司提供。本试验所用氮肥为尿素(N≥46%)。
本研究于2020年开展刈割模拟放牧利用与施氮试验。2019年混播草地基本情况见图1, 试验前混播草地中紫花苜蓿的生物量和盖度远高于两种禾本科牧草, 分枝数则为豆科牧草略小于禾本科牧草, 营养品质方面表现为各牧草总体偏低, 豆科牧草略高于禾本科牧草。由此认为混播草地中豆科牧草占比高于禾本科牧草, 且草地生产性能偏低, 需通过模拟放牧和施氮的方式来增加禾本科牧草在群落中的占比并提高草地生产性能, 以达到维持草地组分平衡和提高草地生产性能的目的。

图1 模拟放牧及施氮前混播草地相关指标Fig.1 Related indexes of legume-grass mixed grassland before and simulated grazing and nitrogen addition
本试验采用L(3×4)混合位级正交试验设计,共16个处理, 随机排列, 小区面积为5 m×5 m, 小区间隔行为1 m。根据前期研究筛选结果, 设置刈前高度[20 cm (S)、30 cm (S)、40 cm (S)]、刈割强度[2 cm (E)、5 cm (E)、8 cm (E)、11 cm (E)]和施氮量 [0 kg(N)∙hm(N)、75 kg(N)∙hm(N)、150 kg(N)∙hm(N)、225 kg(N)∙hm(N)]为试验因素。各试验因子的水平值分布如表1所示。以人工刈割模拟放牧, 在第一次利用后施氮, 施肥方式为开沟深施, 深度为10 cm。各小区在草层高度达到试验设计的刈前高度后刈割, 当刈割后的再生草长到试验设计的刈前高度后继续按照试验设计要求进行刈割。

表1 L16 (31×42)正交矩阵表Table 1 L16 (31×42) orthogonal matrix table
1.3 测定指标与方法
于混播草地草层高度达到试验设计的刈前高度后在每个小区选取3个有代表性的1 m×1 m样方, 采用针刺法测定各混播牧草的分盖度。同时选择3个有代表性的1 m条播样段, 记录各混播牧草的分枝数/分蘖数, 按照试验设计的刈割强度取样, 并测定各牧草地上生物量。在每个小区按照试验设计的刈割强度剪取各混播牧草500 g, 用于测定分种牧草营养品质, 将所有植物样品带回实验室在105 ℃下杀青30 min后转到75 ℃烘至恒重。烘干后的植物样品置于粉样机中粉碎并过1 mm筛, 样段牧草用于测定混合牧草营养品质。每次刈割前均按照上述方法对牧草指标及营养品质进行采集分析。
2019年试验小区统一于豆科牧草初花期进行刈割, 留茬高度为5 cm, 全年共刈割3茬, 分别为2019年6月上旬、7月下旬和9月下旬, 全年不施肥, 其他管理措施与2020年一致。每次刈割前按照留茬5 cm对各混播牧草进行采样, 测定其生物量、盖度、分枝数和营养品质, 分析测定方法与2020年一致。文中2019年生物量为3茬总和, 盖度、分枝数和牧草营养品质则为3茬平均值。用于比较处理前后混播草地各指标变化情况。
采用凯氏定氮法测定牧草粗蛋白含量, 采用范氏法测定牧草中性洗涤纤维含量(NDF)和牧草酸性洗涤纤维含量(ADF)。
1.4 数据处理
相对饲用价值计算方法:

式中: RFV表示相对饲用价值; DMI表示干物质采食量, 单位%BW, 含义为占体重的百分比; DDM表示可消化干物质, 单位%DM, 含义为占干物质的百分比; NDF表示牧草中性洗涤纤维; ADF表示牧草酸性洗涤纤维。值相加记为k, 取 k=K, 该因素的极差R为K—
使用 Microsoft Excel 2010 进行数据整理和极差分析, 并使用 Origin 2021和 R 4.0.2 (R Development Core Team)作图。牧草营养物质含量为一年中所有刈割茬次的平均值。生物量为全年各茬次生物量之和, 粗蛋白含量及相对饲用价值为全年各茬次的平均值。极差分析方法为: 将某一因素x水平下的测定K, 优水平为该因素出现K时的水平, 主次顺序为该因素K的大小顺序。
2 结果与分析
2.1 不同处理下草地利用次数和牧草再生天数
不同处理对草地的生长特征产生了明显的影响。随牧草净收获高度(刈前高度与刈割强度的差值)降低, 草地全年可利用次数增加(4~6次)。休牧天数随利用次数增加而先缩短后延长, 由于各处理的牧草净收获高度一定, 因此其生长速度呈现先加快后减慢的趋势, 在第2~4次利用前达最快。其中处理SEN、SEN、SEN和SEN由于净收获高度较小(12 cm和9 cm), 全年利用次数达6次, 导致其生长速度较快, 但生长期较短(图2)。

图2 不同处理混播草地模拟放牧日期、放牧时间、休牧天数及放牧次数Fig.2 Simulated grazing date, grazing time, days of rest grazing and grazing times of legume-grass mixed grassland under different treatments
2.2 模拟放牧及氮素添加对混播草地群落特征的影响
2.2.1 对混播草地盖度的影响
图3为第1次刈割前和最后1次刈割前混播草地中禾本科牧草的分盖度。经过多次模拟放牧后,大多数处理的禾本科牧草盖度较放牧前有了较大提高, 仅有3个处理的禾草盖度发生下降, SEN由29%降至20%, SEN由30%降至20%, SEN由27%降至15%。处理SEN(21个百分点)、SEN(25个百分点)、SEN(26个百分点)、SEN(41个百分点)、SEN(26个百分点)和SEN(25个百分点)的增幅较大。其中, 处理SEN、SEN、SEN、SEN和SEN经过模拟放牧后的豆禾盖度比等于或接近1∶1。
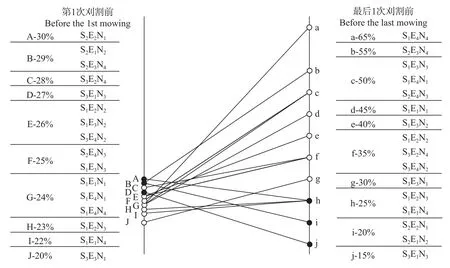
图3 模拟放牧及施氮前后混播草地中禾本科牧草盖度变化Fig.3 Grass coverage of grass in legume-grass mixed grassland before the first simulated grazing (moving) and after the last simulated grazing under nitrogen addition
经过多次模拟放牧与施氮, 豆科牧草盖度较2019年下降部分的降幅为1.22%~73.71%, 上升部分的增幅为5.41%和12.10%; 禾本科牧草盖度下降部分的降幅在2.12%~58.11%, 上升部分的增幅在11.90%~81.90% (图4)。由于豆科牧草盖度降幅大于增幅, 禾本科牧草盖度增幅大于降幅, 所以模拟放牧及施氮能有效增加禾本科牧草在群落中的竞争力。

图4 不同处理下模拟放牧过程中混播草地各牧草分枝及总分枝数变化Fig.4 Changes of branches and total branches of each forage in legume-grass mixed grassland during simulated grazing under different treatments
2.2.2 模拟放牧及氮素添加对混播草地分枝数/分蘖数的影响
由图5可知, 在第1次刈割前各牧草分枝数为紫花苜蓿>无芒雀麦>长穗偃麦草。随刈割茬次增加,无芒雀麦的分蘖数呈上升趋势, 在后期多出现不同程度下降, 这与紫花苜蓿分枝数减少有直接关系。而SEN、SEN、SEN和SEN中无芒雀麦的分蘖数在处理后期仍然出现小幅度增加。在处理后期紫花苜蓿的分枝数除SEN和SEN外, 其他处理均大幅减少, 减少范围在216~642 No.∙m。
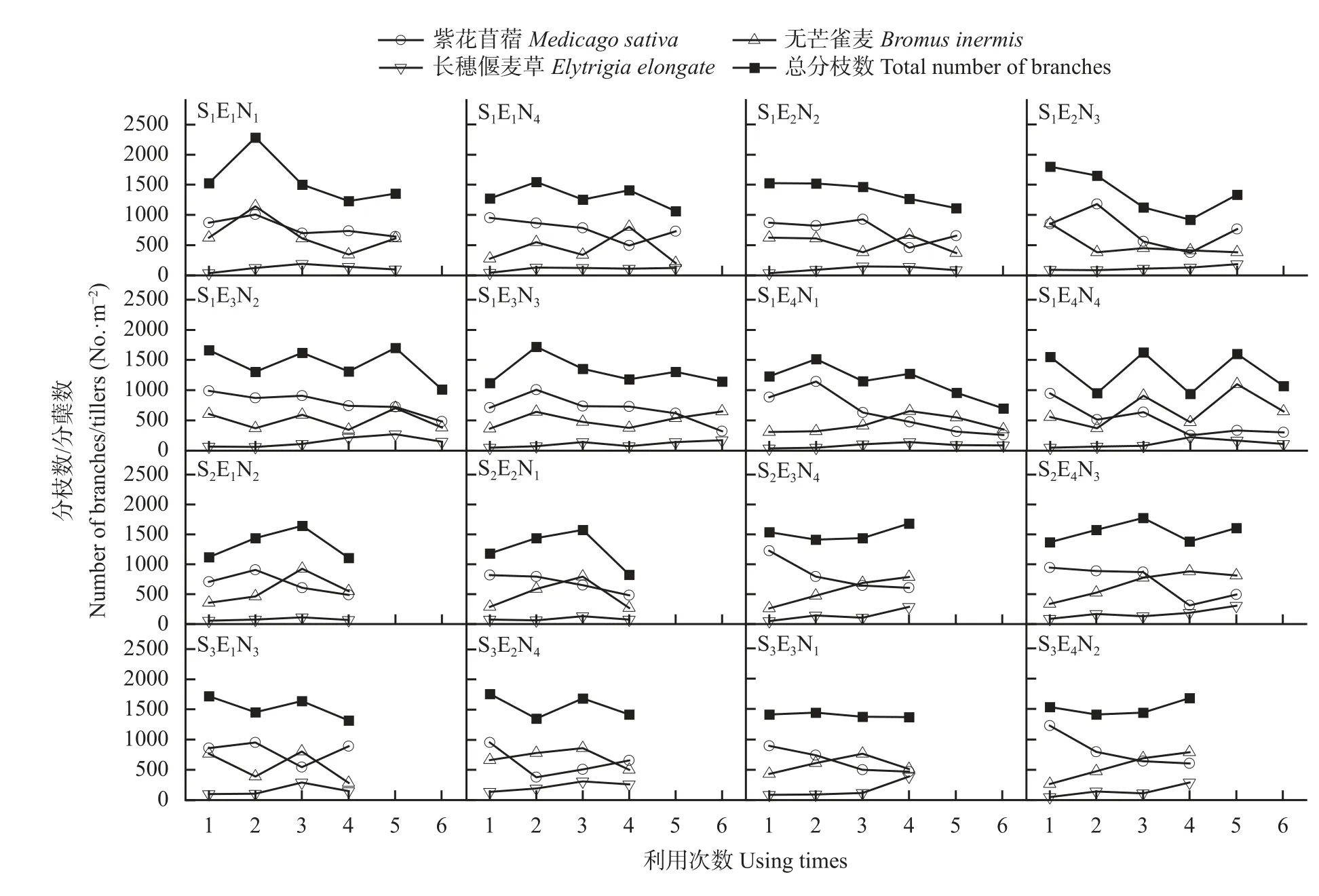
图5 与试验前相比不同处理混播草地牧草盖度和分枝数的变化Fig.5 Changes of coverage and branch number of forages in legume-grass mixed grassland under different treatments compared with those before experiment
在调控后期, SEN、SEN、SEN、SEN和SEN的长穗偃麦草分蘖数均达到250个∙m以上, 恢复效果较好。处理SEN、SEN、SEN、SEN、SEN、SEN、SEN和SEN中的豆科牧草分枝数与禾本科牧草分蘖数相差较多(差值大于400个∙m), 其余处理较为接近。其中SEN、SEN、SEN、SEN和SEN中豆科牧草分枝数与禾本科牧草分蘖数最为接近(差值小于150个∙m)。考虑到对长穗偃麦草的调控作用, 认为处理SEN、SEN和SEN对混播草地分枝数调控作用效果最好。
除SEN中禾本科牧草分蘖数较2019年下降1.91%外, 其余处理的豆科牧草分枝数和禾本科牧草分蘖数均有所增加, 增幅分别为11.62%~82.51%和2.60%~75.21%。由此可见模拟放牧及施氮有利于混播草地中各牧草分枝数的增加(图4)。
2.3 模拟放牧及氮素添加对混播草地生物量结构的影响
对混播草地不同牧草生物量及草地总生物量进行极差分析得出表2。极差(R)越大代表该因素对所讨论指标的影响越大。刈前高度(S)、刈割强度(E)和施氮量(N) 3种因素对紫花苜蓿、长穗偃麦草和草地总生物量产生影响的大小均为S>E>N; 对无芒雀麦生物量产生影响的大小为N>S>E。因此刈前高度对混播草地生物量的影响大于刈割强度, 且高水平施氮更有利于增加无芒雀麦的生物量。

表2 各混播牧草生物量及营养价值的极差(Rj)及各处理因素最佳水平Table 2 Range (Rj) of biomass and nutritional value of each mixed forage and the optimal level of each treatment factor
随刈割茬次增加, 混播草地中紫花苜蓿与无芒雀麦的生物量总体呈下降趋势, 草地总生物量变化趋势与紫花苜蓿基本一致。经过多次刈割调控后,SEN、SEN、SEN、SEN、SEN、SEN、SEN和SEN的豆禾牧草生物量较为接近, 差值小于500 kg∙hm(图6)。
与2019年相比, 豆科牧草生物量除处理13(SEN, 6.21%)和处理14 (SEN, 16.11%)有所上升外, 其他处理降低7.90%~60.91%; 所有处理的禾本科牧草生物量均呈降低趋势, 降幅为4.73%~58.81%。豆科与禾本科牧草生物量下降幅度较小的处理分别为处理11 (SEN)、处理16 (SEN)与处理1(SEN)、处理6 (SEN)、处理8 (SEN)、处理11(SEN)、处理14 (SEN), 降幅均小于10% (图7)。结合图6结果认为: 全年利用5次的SEN处理可使混播草地在高产的同时维持产量组成合理(年总产量可达15 173.41 kg∙hm)。

图6 不同处理下豆禾牧草及混播草地生物量随利用次数的变化趋势Fig.6 Changes of biomass of each forage in legume-grass mixed grassland under different treatments with utilization times

图7 与试验前相比不同处理混播草地牧草生物量的变化情况Fig.7 Changes of biomass of forages in legume-grass mixed grassland under different treatments compared with those before experiment
2.4 模拟放牧及氮素添加对混播草地牧草营养品质的影响
2.4.1 对混播草地牧草粗蛋白含量的影响
3种因素对牧草粗蛋白含量的影响除无芒雀麦为E因素最大外, 其余均为N因素最大, 且所有牧草的“施氮量最优解”均为225 kg(N)·hm, 同时N因素对牧草粗蛋白含量的影响始终大于S因素(表2)。由图8可知各处理中牧草粗蛋白含量的最大值分别为:紫花苜蓿处理14 (SEN, 17.62%), 无芒雀 麦 处理14 (SEN, 15.50%), 长穗偃麦草处理12 (SEN,15.31%),混合草 处 理2 (SEN, 15.51%)。混合草的粗蛋白含量以处理2 (SEN, 15.50%)、处理14(SEN(14.62%)、处理8 (SEN, 14.31%)和处理12(SEN, 14.31%)较高。
经过模拟放牧及施氮对混播草地的调控, 豆禾牧草的粗蛋白含量较2019年有较大程度增加, 增幅分别为15.51%~51.22%和10.90%~45.13%。由此可见在高频率利用的同时对草地施氮可有效提高混播草地牧草营养价值, 且对豆科牧草的促进作用更明显(图9)。
2.4.2 对混播草地牧草相对饲用价值的影响
3因素对紫花苜蓿和无芒雀麦相对饲用价值影响的大小为S>N>E, 对长穗偃麦草相对饲用价值影响的大小为N>S>E, 对混合草相对饲用价值影响的大小为E>N>S (表2)。由图8可知各处理中牧草相对饲用价值的最大值分别为: 紫花苜蓿处理4(SEN, 223.68), 无芒雀麦处理11 (SEN, 139.10), 长穗偃麦草处理16 (SEN, 131.83), 混合草处理13(SEN,184.93)。混合草相对饲用价值以处理13(SEN,184.93)、处 理2 (SEN,176.48)、处 理3(SEN, 173.29)和处理4 (SEN, 172.83)较高。这4个处理下3种混播牧草的相对饲用价值也都处于较高水平。

图8 模拟放牧及施氮对牧草营养价值的影响Fig.8 Effects of simulated grazing and nitrogen addition on the nutritional value of herbage
经过模拟放牧与施氮的调控, 牧草相对饲用价值较2019年有了较大提高。豆禾牧草相对饲用价值的增幅分别为50.20%~125.70%和29.21%~49.42%,豆科牧草增幅更大, 效果更明显(图9)。

图9 不同处理混播草地牧草营养品质增幅情况Fig.9 The increase of nutritional quality of forage in mixed grassland under different treatments
3 讨论
当前河西走廊地区牧草种植收获方式单一(多为一年刈割3~4茬的紫花苜蓿单播草地)。由于没有适合放牧的人工草地, 导致当地天然草地进一步退化, 现代畜牧业始终无法快速高效发展。采用根茎型‘清水’紫花苜蓿建植刈牧兼用型豆禾混播草地既能满足农户收获干草的需求, 也能为牧户提供良好的放牧草场。关于该草地应如何科学利用, 前期已针对刈割施肥制度进行了较详尽的研究, 本文着重研究了放牧配合施氮的利用管理模式。
选择合理的刈前高度对维持混播草地生产性能和群落结构的稳定十分重要。刈前高度决定了牧草株高, 而其对生物量具有较大的贡献率, 所以单株生物量的增加促进了草地总生物量的提升。而牧草发育程度往往与株高呈正相关。提高刈前高度会使牧草木质化程度加剧, 降低牧草营养价值。同时,研究区气候干旱, 土壤盐碱化严重的自然环境更适合根系发达、抗逆性强的紫花苜蓿生长, 过高的刈前高度会使紫花苜蓿对禾本科牧草产生遮阴作用,降低禾草光合速率, 使其生长发育受限, 这是造成禾草盖度降低的主要原因。
刈割强度更多影响了草地利用率和牧草再生,不同牧草对刈割强度的响应也存在差异。有研究表明, 低留茬能降低紫花苜蓿茎叶比, 同时减少残留在地上的枯枝, 促进新鲜枝条从地下发生, 从而提高牧草粗蛋白含量和相对饲用价值。但高留茬却能起到促进禾本科牧草生长、抑制豆科牧草生长的作用, 对于紫花苜蓿占优的混播草地来说, 能有效调控其群落结构的稳定性。留茬越高, 能保留的禾草光合作用组织就越多, 从而增加其分蘖数; 同时紫花苜蓿茎叶比增加、分枝数减少, 削弱了其对水分、光照等自然资源的竞争力, 由此来达到平衡群落结构的目的。
鉴于研究区土壤肥力条件较差, 且高频率、高强度利用会带走大量土壤养分。因此, 适量施氮能补充土壤养分的缺失, 满足植物生长对营养的需求。有研究发现, 禾本科草对草地生产性能的贡献极大。因为禾本科草生长过程中对氮素需求量较高, 而从豆科牧草中转移的氮量又无法满足, 所以施氮能有效促进禾草生长并提高其营养价值, 改善混播草地产量组成。另外, 增施氮肥也可促使禾本科牧草在群落中占据优势地位, 当土壤养分无法满足牧草生长发育时, 适量施氮能补足禾本科牧草对养分的需求, 提升其竞争力, 使混播草地群落的结构更平衡稳定。
科学地放牧利用搭配合理的施肥管理是解决河西走廊地区因常年单播紫花苜蓿所导致的利用方式单一、利用年限短和产量下降速度快等问题的关键措施。模拟放牧利用制度决定了牧草净收获高度及草地利用次数, 随利用频率和茬次的增加, 牧草分枝数开始减少, 草地地上生物量发生了不同程度的下降; 而牧草营养价值则因为苜蓿茎叶比的降低而获得较大幅度提升, 同时有效增加禾本科草盖度和分蘖数, 这是紫花苜蓿不耐牧而禾草适宜多次利用的特性所导致的。
4 结论
通过对河西走廊地区豆禾混播草地群落结构与生产性能的研究发现: 1)增加放牧频率或减小刈割强度, 有利于增加禾本科牧草在群落中的占比, 对比例增长过快的紫花苜蓿有较好的抑制作用, 30 cm刈前高度, 8 cm刈割强度配合225 kg(N)·hm的施氮量可使混播草地群落结构最稳定, 豆禾比最接近1∶1。2)刈前高度和刈割强度是混播草地产草量的主要影响因素, 与刈割利用相比, 放牧使草地产量发生一定程度的下降, 但牧草营养品质则会有较大提升。当刈前高度为40 cm, 刈割强度为5 cm, 施氮量225 kg(N)·hm时草地生物量最大(17 707.80 kg·hm), 当刈前高度为20 cm, 刈割强度为2 cm, 施氮量225 kg(N)·hm时牧草营养价值最高(混合草粗蛋白含量和相对饲用价值分别为15.46%和176.48)。3)综合混播草地群落结构及生产性能的研究结果认为: 河西走廊地区豆禾混播草地最适放牧利用及施氮方式为: 30 cm刈前高度, 8 cm刈割强度以及225 kg(N)·hm的施氮量。
猜你喜欢
农业工程学报(2022年10期)2022-08-22
山地农业生物学报(2022年3期)2022-05-13
土壤学报(2022年1期)2022-03-08
新传奇(2020年17期)2020-08-04
安徽农学通报(2020年7期)2020-05-26
学校教育研究(2020年7期)2020-04-09
环境与发展(2019年8期)2019-10-18
中学生理科应试(2017年6期)2017-09-27
中国中药杂志(2017年17期)2017-09-23
农产品市场周刊(2015年39期)2015-10-31
