
不同农田边界类型的耕地内地表节肢动物功能群生态网络特征分析*
2022-09-20 08:41边振兴王楚翘孙治权
中国生态农业学报(中英文) 2022年9期
边振兴, 王楚翘, 刘 尧, 孙治权, 王 双, 刘 爽, 朱 肃, 于 淼
(1.沈阳农业大学土地与环境学院 沈阳 110866; 2.沈阳农业大学理学院 沈阳 110866; 3.铁岭县自然资源事务服务中心 铁岭 112608)
随着农业用地需求急剧增长, 土地集约化和单一作物种植模式使大量的自然、半自然用地丧失或破碎化。以往研究表明, 集约化是导致农田系统天敌多样性大量丧失和生物害虫控制功能降低的主要原因。在农业景观中, 半自然生境在维持农田天敌多样性, 促进农田生物害虫防治方面有重要作用。植被结构复杂、人为活动干扰低的半自然生境, 可为田间地表节肢动物提供生态源地、避难所等, 有效地增加了田间天敌多样性, 进而提高农田害虫的生物控制能力。同时, 中等景观复杂性假说认为,半自然生境斑块内复杂程度对提高田间物种丰富度的作用最强。当作物生境内出现干扰时天敌可以从作物生境溢出到半自然生境, 当半自然生境内的环境恶化后又回迁到作物生境中, 从而在时空上保持了物种多样性的延续。探索半自然生境对天敌害虫多样性的影响, 利用天敌在半自然生境和农田之间的溢出效应, 有利于加强对天敌的保护, 对于制定可持续的害虫防治策略以及农业景观优化配置具有重要意义。
典型的农业景观一般由农田、半自然生境、人工基础设施及少量的自然生境等镶嵌体构成, 其中,线形的半自然生境被视为农田边界(field margins),即将农田与其他区域相分隔的过渡带, 通常由草带、乔灌木、沟渠、田埂等景观要素组成。近年来, 国内外众多学者对于农田边界特征在害虫生物防控以及天敌多样性保护等方面的生态效益开展了积极探讨。Gayer等证明农田边界有利于天敌类群的栖息和繁衍, 同时, 有利于它们向相邻农田中迁移, 对害虫起到调节和控制的作用; Hoffmann等研究表明, 除木本植物外, 构建临时野花生境与永久半自然生境相结合的边界有助于提升耕地内部地表节肢动物群落多样性; 戴漂漂等研究认为, 农田边界中多花带所占比例为5%~10%时, 可显著增加农田中天敌的密度和丰度, 对提升害虫控制方面具有重要影响。目前针对农田边界类型的研究多集中在灌木篱、草地边缘和小森林斑块等生境, 但对于人为干扰较强, 且硬化程度较高的田间路和生产路等探讨依然不足。因此, 尚需从边界宽度、硬化程度、植被结构等角度划分农田边界类型, 从而更系统性和整体性地分析农田边界类型对耕地内地表节肢动物的影响。
以往关于农田边界和害虫生物防控主要集中于天敌的简单指数, 如多度或多样性。但在生态系统中, 物种之间的相互作用关系同样也是维持生物多样性和生态系统服务功能的重要因素。随着网络分析技术的快速发展, 定量网络拓扑结构的特征参数可以量化物种之间的相互作用关系, 例如天敌害虫之间的竞争、对抗、偏害共栖和共生关系。通过数据采集转化、成对相似矩阵计算等生物信息学过程, 构建生物群落系统的生态网络, 可以揭示节肢动物群落间相互作用, 并考察生态网络的全局拓扑性质。系统认识不同农田边界类型中物种分布格局和物种间相互作用关系是影响害虫与天敌种群发生的基础。以往结合生态网络结构探讨地表节肢动物群落稳定性的研究相对薄弱, 农田边界类型对耕地内部地表节肢动物群落的空间分布以及生态网络特征的影响尚不清晰。
基于此, 本文选取辽宁省昌图县为研究区, 对区内典型农田边界单元开展地表节肢动物调查, 旨在回答以下问题: 1)不同农田边界类型的耕地内地表节肢动物群落分布是否存在差异? 2)不同农田边界类型的耕地内部地表节肢动物功能群的生态网络结构有何不同? 为优化农田边界生境, 提高景观异质性,增强地表节肢动物多样性和天敌类群控害能力提供理论依据, 同时对维护该地区农田生态系统的健康稳定, 更好地发挥生态系统服务功能有重要意义。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于下辽河平原北部, 处辽宁省铁岭市昌图县(123°32′~124°26′E, 42°33′~43°29′N)。气候温和,年均气温7 ℃,年均降水量654.6 mm。土壤为肥力较高的棕壤, 质地较黏重, 是全国最大粮食生产基地, 玉米(Zea mays)种植面积约占耕地面积的56%。经实地调查, 研究样地农田中农作物均为玉米。基于2019年土地利用现状数据, 运用ArcGIS软件, 在较为规整且面积较大的农田中(规格均为500 m×500 m), 筛选出耕作条件、土壤条件、气候条件以及边界形态相对一致的农田—边界单元, 以保证每条边界对农田的影响均相对独立, 将不同边界类型编号为田间路(PR)、沟渠(CD)、林地(WL)、生产路(DR)、草地(GL)。研究区范围及采样单元分布如图1所示。
1.2 农田边界类型划定
随着传统农业向集约化生产的转变, 农业景观中相邻的线性景观要素也发生巨大改变。不同农田边界类型包含不同景观要素, 很大程度上影响环境条件和植被组成。尤其是人类活动对农田边界的干扰, 破坏农业景观中生物多样性和生态系统的稳定性, 严重影响了边缘效应的发挥。在对农业景观进行合理规划和生态景观设计之前, 有必要对现有农田边界进行类型区分。
基于此, 本文以昌图县的农业景观结构特征为分类依据, 考虑到数据完整性将农田边界类型划分为5类。每种农田边界类型随机选择2块规格为500 m×500 m的采样单元, 2块采样单元共4组样带进行取样, 样带中各样点距耕地边界分别为15 m和30 m, 如图1所示, 以保证数据满足统计性检验标准。选取主要的线性半自然生境类型(即农田边界类型)及特征如表1所示。

表1 5种典型农田边界类型及特征Table 1 Five typical types of field margins and characteristics
1.3 地表节肢动物采集与鉴别
2021年8月末采用陷阱法对地表节肢动物进行采集。采样单元内样点布设示意图如图1所示: 垂直于农田边界布设2条样带, 样带之间间隔至少50 m,在耕地内部距交界面0 m、15 m、30 m 3个梯度中布设样点, 每个样点平行布设3个陷阱, 布设间隔至少10 m。陷阱具体布设方法为: 将PP塑料杯(底部直径4.5 cm, 顶部直径9.5 cm, 高11 cm, 容积500 mL)埋入土壤中, 杯口与地表相平齐, 在杯中放入150~200 mL乙二醇溶液(20%)和一滴洗涤剂, 并将杯盖用3根铁丝固定好。

图1 研究区位置、范围及采样单元位置、分布图和采样单元内样点布设示意图Fig.1 Location and scope of the study area and sampling unit and distribution map and map of sample points in the sampling unit
陷阱共布设180个, 在野外放置6 d后取回, 将陷阱瓶里的地表节肢动物放到预先编好编号的PE瓶中, 加入浓度为75%的酒精进行保存, 尽快在实验室进行鉴定分类, 并参考《中国土壤动物检索图鉴》等, 将地表节肢动物鉴定到种。
1.4 数据分析
采用PAST软件计算物种多度、物种数、多样性指数和均匀度指数, 分析地表节肢动物群落的多样性特征。确保数据在满足正态性检验的基础上,利用SPSS 20.0软件进行单因素方差分析(One-way ANOVA), 采用Duncan新复极差多重比较方法对不同农田边界类型和不同距离处的耕地内部地表节肢动物群落多样性差异进行分析。以上分析在Origin软件内进行制图。为更好地理解不同功能群之间的相互联系, 并消除样本大小差异对多样性统计数据的影响, 使用稀疏外推曲线比较地表节肢动物功能群α多样性的差异。为比较不同样本之间功能群结构的相似性, 分别采用ANOSIM分析和非度量多维尺度分析(NMDS)比较不同农田边界类型的耕地内部地表节肢动物群落β多样性的差异性。以上分析均在R软件中进行, 调用iNEXT包和vegan包。
为探索功能群之间的相互关系, 对5种农田边界类型进行生态网络分析, 揭示不同农田边界类型的耕地内部地表节肢动物的共生规律。计算物种之间所有可能的成对Spearman秩相关(), 并根据相关系数的数值大小定义边的权重, 最后利用Gephi软件建立生态网络节点和边的数据库, 以此生成地表节肢动物生态网络关系拓扑图和实证网络拓扑参数, 包括平均路径长度(所有节点之间的平均最短路径)、平均度(连接一个节点到任何其他节点的平均链接数量)、模块指数(物种的互连子集)、平均聚类系数(可通过其他相邻节点达到的节点比例)。基于随机矩阵理论(random matrix theory, RMT)的方法计算随机网络。在生态网络和拓扑特征的基础上计算两个重要的节点特征: 模块内连通度(within-module-connectivity,Z)和模块间连通度(among-moduleconnectivity,P), 计算如下:

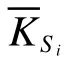
基于模块内连通度( Z)和模块间连通度(P)的网络核心节点识别关键节点, 表征在维持生物群落结构稳定性上可能起重要作用的关键物种。生态网络分析通过Gephi 软件和R 软件, 调用psych 和igraph 包。
2 结果与分析
2.1 地表节肢动物群落组成
通过陷阱法共捕获地表节肢动物4644只, 分属于4纲8目36科。以科为分类单元, 依据食性对地表节肢动物营养功能群进行划分, 分别为: 植食类、天敌类(包括捕食者和寄生蜂)和中性类(包括传粉者和分解者)。其中植食类优势物种(科)为蟋蟀科(Gryllidae)的黄脸油葫芦()和棺头蟋(), 分别占总数的22.49%和53.59%; 天敌类常见物种(科)为步甲科(Carabidae)的蠋步甲()、普通暗黑步甲()、黄斑青步甲(),以及蜘蛛目(Araneae)盲蛛科(Opiliones)盲蛛()和漏斗蛛科(Agelenidae) 漏斗蛛(), 共计707只, 占地表节肢动物总个体数的15.22%; 中性类常见物种(科)包括蚁科(Formicidae)的日本弓背蚁()和黑褐蚁(), 总计203只, 占地表节肢动物总个体数的4.37%。其余27种物种为稀有物种, 占地表节肢动物总个体数的4.33%。地表节肢动物类型及数量可扫描本文首页二维码查看 。
2.2 不同农田边界类型的耕地内部地表节肢动物群落特征
由图2A可知, 草地相邻的耕地内部地表节肢动物物种多度显著高于田间路和林地(=12.081,<0.001); 与草地相邻的耕地内部地表节肢动物物种数显著高于田间路(=5.829,=0.011)。由图2B可知, 林地相邻的耕地内部地表节肢动物多样性指数显著高于其他边界类型(=11.806,<0.001); 与林地相邻的耕地内部地表节肢动物均匀度指数显著高于田间路和生产路(=7.074,=0.006)。

图2 不同农田边界类型的耕地内部地表节肢动物多度、物种数、多样性以及均匀度Fig.2 Abundance, number of species (A), and Shannon-Wiener index and Pielou index (B) of epigaeic arthropods in the arable lands with different types of field margin
在不同农田边界中, 多样性指数随与边界距离增加整体呈递减趋势, 表明农田边界处物种多样性显著高于耕地内部, 验证了边缘效应的存在(图3)。其中, 与田间路(PR)相邻的耕地边界物种多样性指数显著高于耕地内15 m处(=14.51,=0.001), 与林地、生产路和草地(WL、DR、GL)相邻的耕地边界物种多样性指数显著高于耕地内30 m处(=8.793,=0.007;=6.206,=0.02;=4.819,=0.03)。

图3 不同边界类型的耕地内部不同距离梯度地表节肢动物多样性指数Fig.3 Shannon-Wiener index of epigaeic arthropods at different distance gradients within arable land of different types of field margins
2.3 不同农田边界类型的耕地内部地表节肢动物多样性
2.3.1 地表节肢动物功能群α多样性
对比相同农田边界类型的耕地内部天敌类和植食类α多样性特征可知: 所有农田边界类型的耕地内部天敌类物种数、多样性和均匀性均大于或等于植食类和中性类, 除林地(WL)外, 其他农田边界类型的耕地内部植食类物种多度远高于天敌类(表2、图4)。

图4 不同农田边界类型的耕地内部天敌类、植食类、中性类地表节肢动物群落的稀疏外推曲线Fig.4 Sparse extrapolation curves of natural enemies and herbivores and neutral epigaeic arthropod community within arable lands with different types of field margin
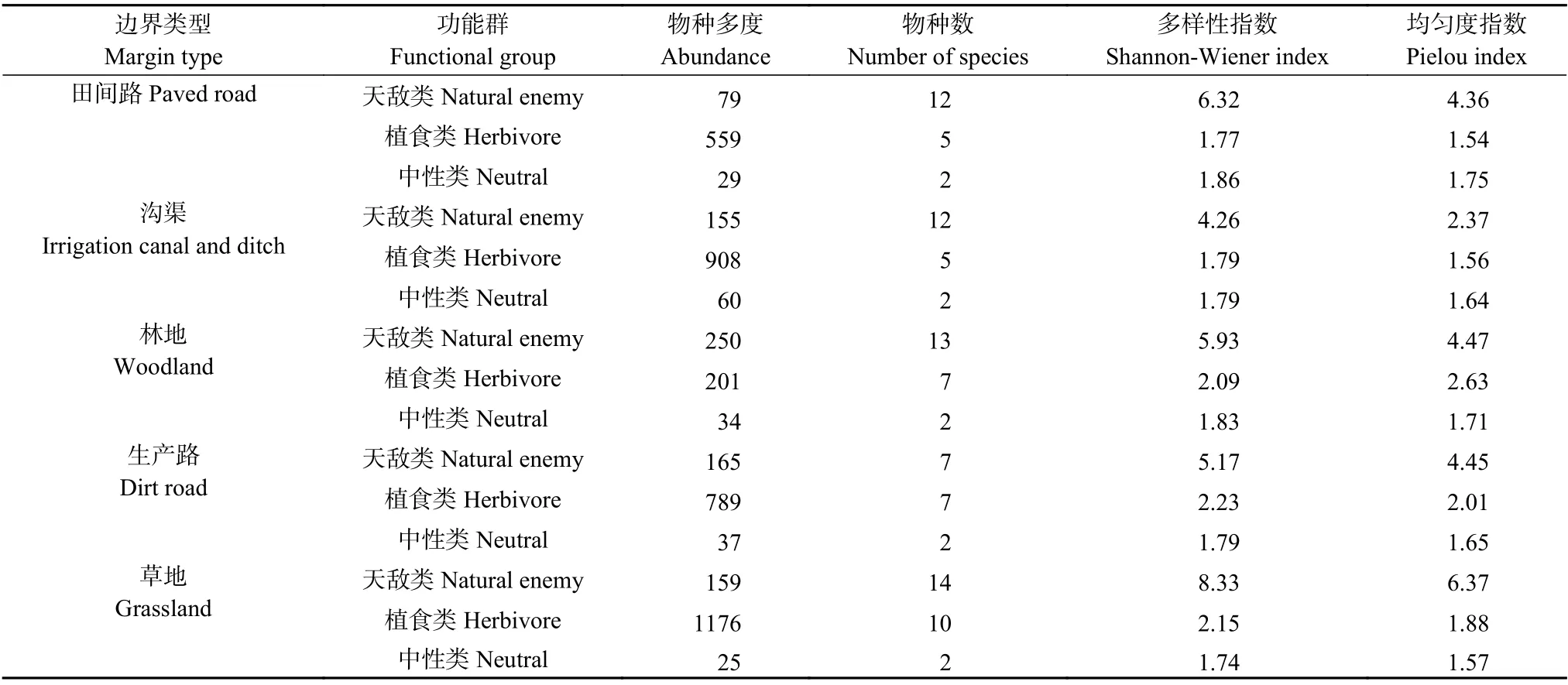
表2 不同农田边界类型耕地内部地表节肢动物功能群多样性Table 2 Functional group species diversity of the epigaeic arthropod community in the arable lands with different types of field margin
对比不同农田边界类型的耕地内部天敌类和植食类α多样性特征可知, 林地(WL)的耕地内部维持着较高的天敌物种多度和较低的植食物种多度, 具有较高的益害比。天敌类群在田间路(PR)、林地(WL)、草地(GL)相邻耕地内部多样性较高, 植食类群在林地(WL)、生产路(DR)、草地(GL)耕地内部多样性和均匀性较高, 中性类群在各边界中多样性指数趋于一致。
2.3.2 地表节肢动物功能群β多样性
ANOSIM分析结果显示, 5种农田边界类型的耕地内部所有地表节肢动物(R=0.613, P<0.001)、天敌类(R=0.495, P<0.01)和植食类(R=0.781, P<0.001)群落存在着较高的异质性。而中性类节肢动物群落没有差异(R=0.049, P=0.62)。
NMDS分析结果显示(图5), 整体上, 沟渠(CD)和生产路(DR)耕地内部的所有地表节肢动物群落结构和物种组成存在一定重叠, 表明地表节肢动物群落具有较高的相似性; 田间路(PR)、林地(WL)、草地(GL)之间分离较为明显, 表明其耕地内部物种组成差异性较大, 群落存在高度异质性。天敌类群和植食类群在林地(WL)、草地(GL)的耕地内部更倾向分布于第一排序轴右侧, 其他边界类型分布在第一排序轴左侧。而所有边界类型相邻的耕地内部中性类群均呈现为高度聚集状态, 体现中性类群的高度均质性。以上排序结果表明, 农田边界植物多样性较高(WL、GL)的耕地内部与农田边界植物多样性较低(PR、CD、DR)的耕地内部的地表节肢动物群落结构有明显差异, 且在植食类群中体现最为明显。

图5 不同农田边界类型的耕地内部所有(A)、天敌类(B)、植食类(C)、中性类(D)地表节肢动物群落非度量多维度(NMDS)分析Fig.5 NMDS of all epigaeic arthropod (A), natural enemies (B), herbivorous (C), and neutral (D) epigaeic arthropod communities within arable lands with different types of field margin
2.4 不同农田边界耕地内部地表节肢动物群落生态网络分析
为了解不同地表节肢动物群落之间的相互作用关系, 分别构建5种农田边界类型的耕地内部地表节肢动物群落生态网络。整体上, 5种农田边界类型的耕地内部物种之间以正相关为主, 天敌和天敌类以及天敌和植食类的相关性较强。与随机网络相比,沟渠(CD)、林地(WL)、草地(GL)在实证网络中的平均路径长度(2.721>2.526、2.796>2.395、4.590>2.377)、平均聚类系数(0.500>0.152、0.656>0.172、0.778>0.150)和模块指数(0.440>0.366、0.580>0.342、0.530>0.338)较高(高于随机网络)(表3), 具有更稳定的生态网络拓扑特征。对实证网络而言(图6), 林地(WL)和草地(GL)的密度(0.183和0.183)、平均度(3.111和4.000)和平均路径长度(2.796和4.590)较高, 而模块指数(0.580和0.530)较低, 地表节肢动物物种之间网络结构更稳定, 天敌和植食类群之间有很强的共生性。沟渠(CD)、林地(WL)、草地(GL)的平均聚类系数较高(0.500、0.656、0.778)节点与相邻节点的连接性较强。田间路(PR)和生产路(DR)具有较高模块指数(0.710和0.660), 表明存在较少的子网络, 生态网络结构相对简单, 天敌和植食类群之间可能存在不稳定性。

图6 不同农田边界类型的耕地内部天敌类、植食类和中性类群落生态网络结构图Fig.6 Ecological network structure diagrams of the natural enemy, herbivorous and neutral epigaeic arthropod communities within the arable land with different types of field margin
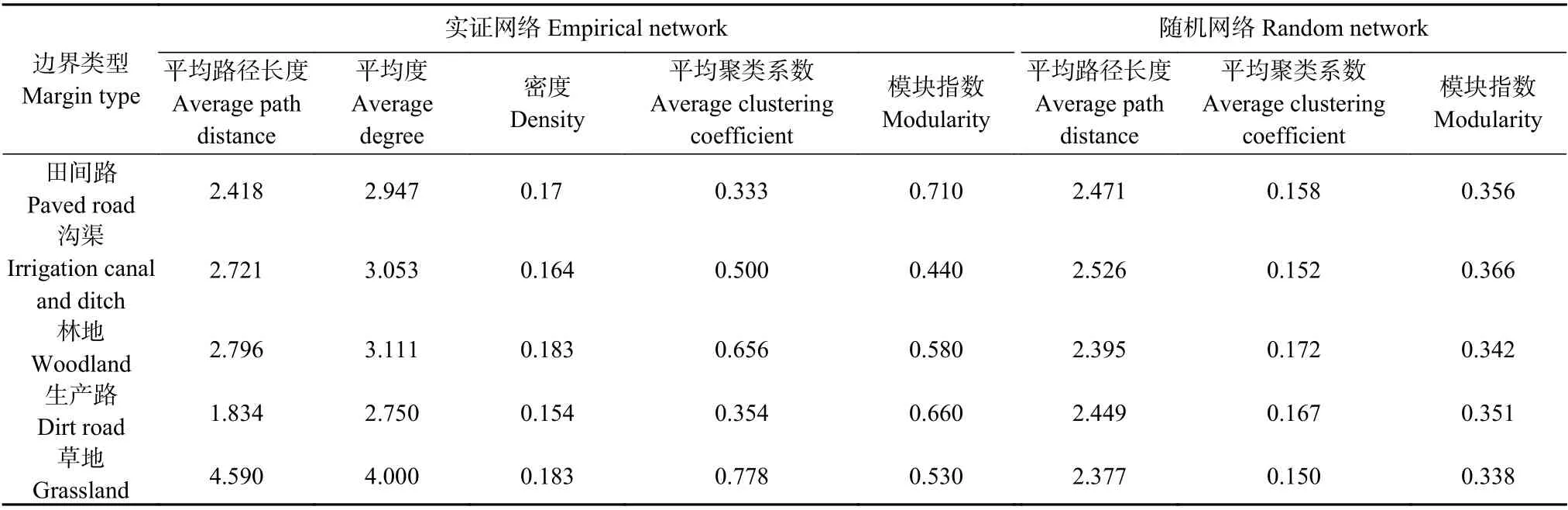
表3 不同农田边界类型的耕地内部地表节肢动物群落生态网络拓扑参数Table 3 Topological parameters of the ecological network of epigaeic arthropod communities within arable lands with different types of field margin
将生态网络划分多个模块, 形成网络中高度连接的区域, 利用各模块表征景观异质性, 识别模块中的节点表征关键种群。利用节点的拓扑特征可将节点属性分为4种类型, 即模块中心点(Z>2.5且P>0.62)、连接节点(Z<2.5且P>0.62)、网络中心点(Z≥2.5且P≥0.62)、外围节点(Z<2.5且P<0.62),其中网络中心点是在整个网络中具有高连通度的节点, 通常将该节点定义为关键物种。结果表明, 田间路(PR)和生产路(DR)存在4个关键物种, 主要属于蟋蟀科黄脸油葫芦和棺头蟋、蚁科黑褐蚁和步甲科蠋步甲; 沟渠(CD)、林地(WL)、草地(GL)存在9个关键物种, 主要属于蟋蟀科黄脸油葫芦棺头蟋、步甲科普通暗黑步甲、中华婪步甲(Harpalus sinicus)、大暗黑步甲(Amara macronota)、黄斑青步甲和蠋步甲以及蜘蛛目盲蛛和漏斗蛛。因此蠋步甲、黄脸油葫芦、棺头蟋在5个生态网络中都是关键物种, 在农业景观中起到维持天敌多样性和稳定生态网络的重要作用。
3 讨论
3.1 不同农田边界类型的耕地内部地表节肢动物群落结构存在差异
草地(GL)相邻的耕地内部地表节肢动物物种多度和物种数以及林地(WL)相邻的耕地内部地表节肢动物多样性和均匀性显著高于其他边界类型, 这可能是由于植被种类丰富的生态环境比种植单一植被的生态环境能为节肢动物群落提供更丰富的物种库, 增加其种类和数量, 为耕地内生物提供更稳定的生存环境以及更长效的资源供给。以往研究多证实边界中草带和林带对耕地内地表节肢动物群落的影响。如边振兴等研究表明, 与消亡型相比, 完整型和残缺型林带相邻的农田物种多度显著较高。张旭珠等的研究同样表明, 人工林地相邻的小麦(Triticum aestivum)田比人工种植草带和自然演替草带相邻的小麦田中维持着显著较高的天敌物种。本文在以往研究基础上, 选取不同植被群落结构和边界宽度的农田边界类型—沟渠、林地、生产路,维持了具有显著差异的地表节肢动物群落结构。植被群落结构的复杂性和物种多样性促进耕地内部地表节肢动物群落的多样性。同时, 宽度>4 m的农田边界(田间路、沟渠、林地、草地)更有利于地表节肢动物的扩散和迁移。因此, 农田边界类型、边界宽度和植被群落复杂程度影响相邻耕地内部地表节肢动物群落的分布, 进而影响天敌对害虫的生物控制效率。
5种农田边界类型均呈现出明显的边缘效应, 这是由于相邻栖息地的物种会向边缘溢出, 由于溢出效应的潜在影响, 农田边界拥有较高的物种活动密度, 产生边缘效应。Blary等研究指出, 农田边界作为生物扩散的重要廊道, 起到提高物种扩散和保护农田中群落稳定性的作用, 对增加农田生物多样性极为重要。因此, 在设计与规划农业景观格局时, 要充分考虑各景观要素对地表节肢动物分布的影响, 合理优化农田边界类型, 提高景观异质性, 保障农业景观格局的健康和稳定。
3.2 不同农田边界类型对耕地内部地表节肢动物群落结构影响显著
林地和草地相邻的耕地内部地表节肢动物群落组成更复杂, 物种多样性更高。天敌类群在田间路、林地、草地的耕地内部多样性较高, 主要由于边界宽度、林下植被和草本植被可以改善环境条件并提供枯枝落叶等食物, 有助于保护天敌多样性, 并增强与植被相关的生态系统服务功能。然而, 沟渠相邻的耕地内地表节肢动物多样性较低, 主要由于采样单元选择的村庄内部沟渠受人为干扰破坏严重,附近自然植被稀缺, 无法为耕地内部地表节肢动物提供良好的栖息地和迁移场所。段美春研究结果表明, 农田边界宽度≥3 m时能够更有效地为天敌创建适宜的栖息地, 进而控制农田中的病虫害。植食类群在林地、生产路、草地的耕地内部多样性和均匀性较高, 一方面是周围景观中半自然植被在提高天敌多样性的同时, 可能也为害虫提供了相应的资源及适宜的微环境, 导致害虫数量大量增加; 另一方面, 硬化度相对较高的生产路会降低景观的连通程度, 且农村道路受人类干扰程度较大, 阻碍天敌活动, 导致植食类群的多样性增加。因此要依据农田种植作物和景观需求确定缓冲带功能, 在保留原有农田边界的基础上, 新建合适的植物组合的农田缓冲带。
中性类节肢动物群落差异不显著, 天敌类群和植食类群在林地、草地的耕地内部的群落结构与其他边界类型有明显的差异, 且在植食类群中体现最为明显。主要由于半自然生境复杂的植被群落结构可为害虫躲避天敌的捕食提供屏障, 导致天敌捕食害虫的难度增加, 害虫捕获效率下降。与Martin等的研究结论相似, 具有高植被多样性的生境中往往具有较丰富的植食类群。植食类群在草地和林地中相似性较高, 因为植物作为植食类群的食物来源, 其空间分布格局及群落结构特征与植食类群多样性有着密切的关系。因此, 在研究半自然生境生物控制服务功能时, 不能仅仅考虑天敌和害虫的发生数量, 还需深入探讨不同的农田边界类型对天敌和害虫群落关系的影响。
3.3 不同农田边界类型对耕地内地表节肢动物功能群的共生关系有较大影响
一般而言, 生物防治效率随着天敌的多样性增加而增加。然而, 由于天敌之间的对抗性作用, 不同的天敌组合可能削弱对害虫的抑制作用。生态网络方法可以加强对天敌类群和植食类群之间关系的理解。研究结果表明, 不同农田边界类型的耕地内部地表节肢动物互作网络的连接紧密程度存在差异,在沟渠、林地、草地3种边界的耕地中物种流动性较强, 当生境内种类、数量发生变化时能够迅速做出调整, 具有较强的调节能力。天敌和植食类群之间有很强的共生性, 具有相对稳定的生态网络结构,整个网络系统中表现出较强的抗性, 是一种有效的害虫抑制系统。这是由于草带、树篱等具有独特的廊道结构特征, 通过连接不同景观斑块和生境, 可作为昆虫迁移扩散的重要廊道, 是形成完整生态功能网络的重要前提。而田间路和生产路的耕地内容地表节肢动物生态网络结构相对简单, 天敌和植食类群之间可能存在不稳定性。主要由于农村道路受到人为干扰较多, 影响物种的迁移和活动, 使整个生态网络处于不确定状态。因此, 为构建持续稳定的农业景观和长期存在的非农生境, 应减少人为活动对农业景观的干扰, 农田边界优先选择本土植物, 种植多样化的乔木, 进行乔灌草搭配等, 进而保障农业景观格局的健康和稳定。
5个农田边界的耕地内部地表节肢动物生态网络中共有的关键物种与优势物种和常见物种相似。表明天敌和植食类群在生态网络稳定性方面发挥重要作用, 生态网络中的关键物种与群落的功能密切联系, 关键物种的存在或消失可能会使群落的结构和功能发生变化。已有研究表明植被因子、生物量、物候差异以及捕食者内部存在的竞争或者捕食关系等都会影响天敌类群对害虫的控制效率。而5个生态网络之间存在显著差异的物种是步甲科和蜘蛛目,表明蜘蛛和步甲是引起网络中共生关系稳定的重要物种, 其中, 植被复杂性对天敌密度会产生正效应和加性效应。与相对硬化农田边界(田间路、沟渠)相比, 植被结构复杂度较高的沟渠、林地和草地可以增强天敌和植食类之间的共生关系, 加强天敌对猎物的流动性和搜索范围, 对天敌的合理保育与害虫的防控消长起到至关重要的作用, 更有利于协调农田生态系统的稳定。因此, 农田周边存在林地、草地等半自然生境斑块有利于增强斑块间的连通度,为天敌提供栖息地及猎物资源, 提高农田生态系统对天敌类群的涵养潜力。
4 结论
通过调查5种典型农田边界的耕地内部地表节肢动物群落和生态网络结构, 得出如下结论: 1)农田边界处呈现明显的边缘效应, 且具有较高植物多样性边界的耕地内部维持着较高的地表节肢动物多样性水平和相对独特的地表节肢动物群落结构。2)林地、草地生态网络结构相对稳定, 面对外界干扰体现出较强的韧性和可调节性。3)田间路和生产路生态网络较为脆弱, 易受外界干扰; 边界中杂草丰富且宽度>4 m的田间路可以为地表节肢动物提供更丰富的食物、栖息场所等资源, 对天敌类群的移动和捕食活动有一定的促进作用。4)沟渠的耕地内部地表节肢动物群落生态网络在面对外界干扰时, 有较强的恢复力, 天敌和植食类群共生关系较强。本文进一步验证了农田周边半自然边界可以增加农业景观异质性, 从而提升生物多样性保护功能。在未来的农业景观构建中, 不仅要注重提升生物多样性, 还应进一步探索物种之间的生态网络关系和共现规律。通过合理使用化学农药、营造合适的植被群落结构以及构建适宜的边界宽度, 增加天敌多样性, 加强耕地内天敌和植食类群之间的生态网络稳定性。
猜你喜欢
山地农业生物学报(2022年3期)2022-05-13
小猕猴智力画刊(2022年3期)2022-03-29
小猕猴智力画刊(2022年3期)2022-03-28
土壤学报(2022年1期)2022-03-08
学校教育研究(2020年7期)2020-04-09
东坡赤壁诗词(2018年1期)2018-03-31
中学生理科应试(2017年6期)2017-09-27
红蜻蜓(2017年2期)2017-03-29
中国新闻周刊(2015年31期)2015-08-28
岁月(2009年10期)2009-10-28
