
等养分投入下冬种紫云英比秸秆还田更有效抑制稻田CH4的产生和排放
2022-09-19 09:26:54陈洪儒鲁艳红廖育林万运帆王开悦张志伟秦晓波
植物营养与肥料学报 2022年8期
陈洪儒,鲁艳红,廖育林,王 斌,万运帆,王开悦,张志伟,3,聂 军,秦晓波*
(1 中国农业科学院农业环境与可持续发展研究所 / 中国农业科学院农业农村碳达峰碳中和研究中心 /农业农村部农业环境重点实验室,北京 100081;2 湖南省土壤肥料研究所,湖南长沙 410125;3 南京信息工程大学,江苏南京 210044)
温室气体浓度的增加已经使全球平均地表温度比工业化前提高1.1℃[1]。甲烷(CH4)是仅次于二氧化碳(CO2)的人为温室气体[2],尽管在大气中的含量远低于二氧化碳,但CH4吸收热红外辐射的效率要高得多,在100年时间尺度上,其单位质量的全球变暖潜力(GWP)是CO2的28倍。水稻是全球最重要的粮食作物之一,水稻种植的淹水环境促进了CH4的大量排放。我国是世界上最大的水稻生产国,稻田CH4总排放量的估算值为7.7 Tg/a, 约占世界稻田CH4总排放量的30%[3]。同时我国还是世界上最大的氮肥生产国和消耗国,化肥的不合理施用促进了农田温室气体大量排放,也加剧了农业面源污染。2015年我国农业农村部明确提出“一控两减三基本”的目标,控制农业灌溉用水,减量化肥和农药使用,对畜禽粪便、农膜、作物秸秆进行综合循环再利用。研究表明,化肥减量配施有机肥能够促进水稻增产、提高氮肥利用率,不同种类有机物料对稻田甲烷排放影响的研究结果不尽相同[4-5]。水稻秸秆和冬种绿肥是我国南方稻田重要的还田有机物料,含有丰富的碳、氮元素,有利于减少化肥施用、增加土壤养分、改善生态环境,实现农田可持续利用[6-7]。关于绿肥与水稻秸秆还田对稻田CH4排放的影响已有较多研究,一般而言水稻秸秆还田会增加CH4排放[4,8],而绿肥所产生的影响还不确定。
稻田CH4排放是土壤厌氧区域产生的CH4经过氧化和传输的净效应。在淹水稻田中,一系列微生物经过水解、发酵等过程将复杂有机物矿化降解成简单小分子化合物,被产甲烷菌利用生成终端产物CH4[9-11]。CH4主要由乙酸发酵和氢气还原二氧化碳(H2/CO2)两种途径产生,其中乙酸发酵途径通常被认为占生物产CH4总量的2/3左右[12-13]。但在不同的稻田土壤中,两种途径对产甲烷的贡献率并非固定不变,实际测定结果和理论值的差异主要取决于土壤微生物种群和土壤有机物料种类的差异[14]。稻田土壤中嗜甲烷菌多分布于有氧气释放的水稻根际、土壤-水界面和淹水溶氧层,只使用CH4作为碳源和能源,稻田土壤厌氧区产生的90%以上的CH4可被有氧区的甲烷氧化菌再氧化[12]。CH4从土壤向大气传输主要包括两种途径:土壤—水稻植株—大气途径和土壤—田面水—大气途径,其中植株传输在总CH4排放中占绝大部分[15]。稳定碳同位素自然丰度法有助于了解稻田CH4排放的主要控制过程,这是由于稻田土壤产生的CH4以不同传输途径向大气排放过程中会发生明显的碳同位素分馏,12CH4较13CH4能更快地被传输而释放到大气中,CH4产生、氧化和传输中CH4的δ13C值具有明显差异。另外,利用该方法,还可通过观测CH4和CO2的δ13C值估算乙酸发酵和H2/CO2途径对总CH4生成的相对贡献[15-16]。
目前关于化肥与绿肥或水稻秸秆还田结合的不同模式对稻田CH4季节排放规律的研究较多,但对其影响CH4排放机制,尤其是在等养分投入下的节肥减排效应研究还有待加强,另外,关于稻田CH4产生、氧化、传输的不同特征及δ13C特征的研究也较少。本研究以超级稻为研究对象,设置等养分投入对比试验,主要利用培养试验、大田监测和稳定同位素等手段,研究全量还田下冬种紫云英和水稻秸秆的节肥减排效应。以厌氧培养试验模拟稻田原位厌氧土层环境,研究绿肥与秸秆处理对稻田CH4产生潜力和氧化潜力的影响,以及两种关键产CH4途径的贡献率,揭示冬种紫云英的节肥和CH4减排机制。
1 材料与方法
1.1 试验点概况
试验于2021年在湖南省农业科学院高桥试验基地进行,该地位于季风气候区,年均温约16.8℃,年降水量约1400 mm,主要集中在春、夏两季。试验前及试验后各处理供试土壤的基本理化性状见表1。

表1 试验地土壤基本性质Table 1 Basic soil properties in this experimental field
1.2 试验设计
试验共设6个处理:1) 不施肥(CK);2) 单施化肥(CF);3) 化肥+秸秆全量还田(S);4) 化肥+紫云英全量还田(M);5) 化肥+秸秆和紫云英全量还田(MS);6) 化肥+秸秆和紫云英全量还田+熟石灰(MSC)。供试紫云英品种为‘湘紫1号’,紫云英处理的小区于上季晚稻收获后(2020年10月14日)播种,2021年4月中旬盛花期收割称重测产,重复小区紫云英均量翻压还田,紫云英鲜草还田量为22500 kg/hm2。秸秆处理的小区于上季水稻收获后称重测产,重复小区均量覆盖还田,稻草还田量为2625 kg/hm2。供试紫云英和稻草的养分含量见表2。MSC处理于水稻移栽前(2021年6月6日) 重复小区撒施熟石灰3.4 kg并旋耕。除CK处理外,各施肥处理养分投入量相同,均为N 210 kg/hm2、P2O575 kg/hm2、K2O 120 kg/hm2,各处理养分投入量详见表3。施用氮、磷、钾肥的种类分别为尿素(含N 46%)、钙镁磷肥(含P2O512%)、氯化钾(含K2O 60%)。磷肥在移栽前(2021年6月7日)作基肥施入;氮肥和钾肥50%作基肥,50%作追肥在分蘖期施入。小区面积30 m2,3次重复,共18个小区,随机区组排列。供试超级稻品种为籼型两系杂交稻‘晶两优华占’,于2021年6月9日移栽。除中期晒田外,其他时期为间歇灌溉,水稻收获前10天自然落干。其他田间管理措施按当地常规方法进行。

表2 供试紫云英和稻草的养分含量Table 2 Nutrient contents of the tested Chinese milk vetch and rice straw

表3 不同处理氮、磷、钾养分和熟石灰投入量(kg/hm2)Table 3 Nutrient and slaked lime input amount in different treatments
1.3 样品采集及测定
1.3.1 稻田CH4排放通量测定 采用拥有专利权的密闭式静态暗箱[17],采样箱由底座和透明顶箱组成。顶箱高度为100 cm,长宽均为37 cm,侧壁包有反光隔热膜以防止采样期间箱内温度变化过大。各小区底座于生长季埋于田间,底座内罩1株水稻植株。取气时间为水稻分蘖初期的2021年6月21日上午9:00—10:00,此时紫云英还田72天,秸秆还田250天,水稻生长较旺盛,CH4排放相对较高。取气时将顶箱垂直安放在底座上部5 cm深的凹槽内并加水密封,使用温室气体自动采样器分别在扣箱后第0、10、20、30 min采集气体收集于500 mL铝箔气袋中,密封恒温保存。为防止人为扰动造成误差,在田间搭设木桥通往各小区底座。气样中CH4浓度和δ13C-CH4由气体浓度/同位素分析仪(Picarro G2201-i,USA)测定。CH4排放通量由箱中浓度随时间的变化率计算得出:
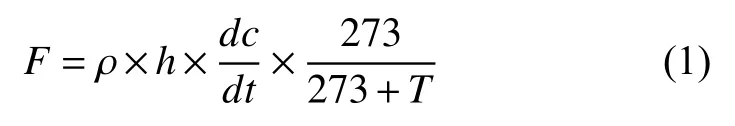
式中:F为CH4排放通量,mg/(m2·h);ρ为标准状态下CH4的密度,取值为0.714 kg/m3;h为采样箱高度,m;dc/dt为单位时间采样箱内CH4浓度变化率,mL/(m3·h);T为采样时箱内温度,℃。
1.3.2 田面水溶存CH4采集与分析 用静态箱取气的同时,使用60 mL注射器在每个底座内取田面表层水(无气泡)。于实验室内将各注射器中水样排出至剩余水样30 mL,将30 mL高纯氦(纯度>99.999%)注入各注射器。室温下将注射器置于振荡器震荡5 min,静置10 min,使注射器内达到气液两相平衡。最后将注射器上部30 mL气体转入12 mL真空瓶内,用气相色谱仪(Agilent 7890A,USA)测定气样中CH4浓度。田间操作及气样的测定分析详见文献[18-19]。
1.3.3 土壤样品采集与分析 培养试验土壤采集自田间取气和田面水取样当天(2021年6月21日)。在每个小区按“S”形随机取0—20 cm土样组成一个混合样,剔除石砾和植物残体,所取土样随即分成2份装入密封袋带回实验室冷冻保存,一份用于测定理化性质,一份用于室内培养实验。供试土壤有机质(SOM) (NY/T 1121.6—2006)、全氮(TN) (LY/T 1228—2015)、全碳(TC)[20]在湖南省土壤肥料研究所农化检测中心测定,可溶性有机碳(DOC)[21]和可溶性总氮(TDN) (HJ 667—2013)在中国科学院亚热带农业生态研究所测定。
1.3.4 CH4产生与氧化潜力培养试验 甲烷产生潜力厌氧培养试验:1) 根据折合干重20 g和土壤含水量计算水土比为2∶1时的湿土重和加水体积,每个处理取3个重复小区混合湿土样称取相应湿土重,置入100 mL培养瓶,加入去离子水,搅拌使土样分散混合均匀,密封瓶口,每个处理4次重复;2) 在30℃黑暗条件下进行10天预培养,预培养结束后进行反复几次抽真空和高纯氮气吹扫循环,最后充满氮气至常压,再于30℃黑暗条件下在大气压下缓慢平衡24 h以消耗培养瓶中的残留氧气;3) 在0 h取气,取气之前将培养瓶轻轻水平旋转1~2 min,注射器取顶部30 mL气样转入12 mL真空瓶中,取气后补充30 mL高纯氮气至常压,放入30℃黑暗培养箱继续培养48 h再次取30 mL气样。
甲烷氧化潜力好氧培养试验:1) 根据折合干重20 g和土壤含水量计算土壤含水量为70%时的湿土重和加水体积,每个处理取3个重复小区混合湿土样称取相应湿土重,置入100 mL培养瓶,加入去离子水,密封瓶口,每个处理4次重复;2) 将培养瓶放入30℃黑暗培养箱中预培养24 h;3) 用微型注射器将1 mL纯CH4注入培养瓶中以形成较高的内部浓度,30℃条件下120 r/min振荡24 h。在0、24、48、72 h取顶部30 mL气样转入12 mL真空瓶中,取气之前将培养瓶轻轻水平旋转1~2 min,每次取完气补充30 mL新鲜空气。
取样完成后用气相色谱仪测定CH4气体浓度,计算甲烷产生/氧化潜力:
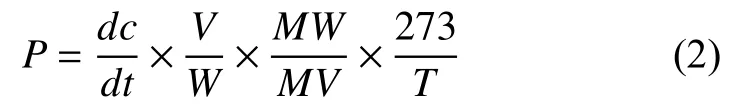
式中:P为 CH4产生/氧化潜力 [μmol/(mol·d)];dc/dt为培养瓶上空CH4浓度单位时间变化率[μmol/(mol·d)];V为培养瓶内气体体积(L);MV为标准状态下1 mol气体的体积(22.4 L);MW为CH4的分子量(g);W为干土重(g);T为培养温度(K)。
1.3.5 δ13C-CH4与关键产CH4途径 用气体浓度/同位素分析仪(Picarro G2201-i,USA)测定田间排放、田面水溶存和厌氧培养试验产生气体的δ13C-CH4和δ13C-CO2,根据CH4和CO2同位素标气拟合曲线计算样品气体中δ13C的实测值。稳定碳同位素组成(‰)利用公式(3)计算:

式中:Rsa和Rst分别表示待测样品和标准样的同位素比值;13C/12C对应于国际标准ViennaPDB,分析精度为1‰。
采用简单混合模型(公式4、5)计算乙酸发酵和二氧化碳还原两种关键产CH4途径占全部CH4的比例[22-23]:

式中:δ13C-CH4是测定的全部产生途径的CH4碳同位素值;δ13C-CH4(CO2)为CO2还原途径产生的CH4碳同位素值(利用公式4计算),CO2分馏系数 αCO2一般采用某一固定值,本研究使用1.079来计算[22],该系数在一个纯培养研究中确定,其中CO2还原是天然湿地中CH4产生的唯一途径[24],已应用于确定湖泊沉积物中的CH4产生途径[22,25];fCO2和face分别为二氧化碳还原和乙酸发酵2个途径占全部CH4的比例 (假设 fCO2+face=1);δ13C-CH4(ace)为乙酸发酵途径产生的CH4碳同位素值,在新鲜有机质充足的假设下,其值为-43‰,而在新鲜有机质极度消耗的假设下其值为-27‰[25],本研究使用二者平均值-35‰来计算。
1.4 数据处理
使用微软Excel (v.2010, Microsoft)整理并初步计算数据均值和标准误差;用SPSS (v.23.0, IBM)进行统计分析,通过单因素方差分析(ANOVA)识别不同施肥处理间稻田CH4排放通量、田面水溶存CH4浓度、稻田土壤CH4产生潜力和氧化潜力的差异,用最小显著性检验(LSD)法比较不同处理间的差异显著性(P<0.05),并对各观测变量做Spearman相关分析,在0.05水平和0.01水平检验相关系数显著性;用GraphPad Prism (v.7, GraphPad Software)和R程序[26]绘图。
2 结果与分析
2.1 稻田土壤培养过程中CH4浓度变化
在厌氧培养的48 h内,各处理CH4浓度均一致呈现升高的趋势(图1A)。S处理的CH4浓度及提升幅度最大,远高于其他处理。M处理的CH4浓度和提升幅度均最小,甚至低于CK。MS和CF的CH4浓度和提升幅度十分接近,而MSC的提升幅度大于CK。
相反,在好氧培养的72 h内,各处理CH4浓度均随培养时间延长而下降(图1B)。CK处理的CH4浓度下降幅度最大,其他处理的CH4浓度下降幅度非常接近。

图1 厌氧培养(A)和好氧培养(B)过程中各处理CH4浓度变化Fig.1 Variation of CH4 concentration during anaerobic incubation (A) and aerobic incubation (B)of different treatments
2.2 稻田土壤CH4产生潜力和氧化潜力
厌氧条件下,不同处理稻田土壤CH4产生潜力范围为 0.0039~0.0427 μmol/(mol·d),无统计学上显著差异(图2A)。S、MS和MSC处理的CH4产生潜力均高于单施化肥(CF)和不施肥(CK)处理。M处理的CH4产生潜力最低,比其它5个处理平均低61.04%,比S处理低83.16%。
好氧条件下,不同处理稻田土壤CH4氧化潜力范围为 2.75~3.85 μmol/(mol·d),CK 处理的 CH4氧化潜力显著高于所有施肥处理(P<0.05) (图2B)。CK、CF、MS、MSC处理的CH4氧化潜力大小顺序与CH4产生潜力相反。M处理的CH4氧化潜力最低,比其它5个处理平均低7.56%,比S处理低5.36%。
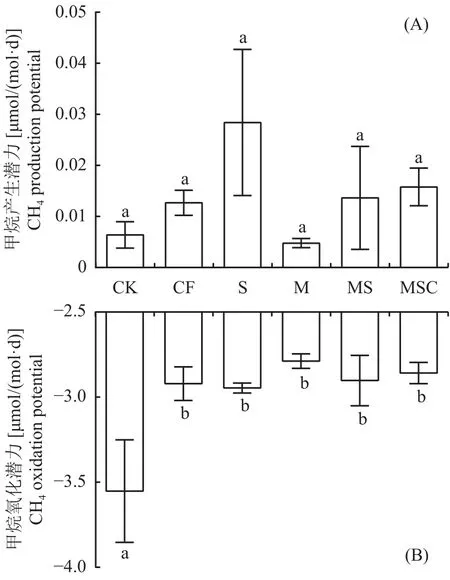
图2 各处理CH4产生潜力(A)和氧化潜力(B)Fig.2 CH4 production potential (A) and oxidation potential (B) of different treatments
2.3 不同处理稻田土壤CH4关键产生途径
CH4产生潜力厌氧培养期间首末两次取的CH4均以H2/CO2还原产生途径为主导(图3)。培养0 h各处理乙酸发酵产CH4途径所占比例范围为9.93%~32.56%,培养48 h后各处理乙酸发酵产CH4途径所占比例范围为22.31%~50.46%,因此相较于0 h,培养48 h后各处理CO2还原产CH4途径所占比例均下降,乙酸发酵产CH4途径所占比例均增加。此外,培养期间各处理乙酸发酵产CH4途径比例平均值大小顺序为 MS>S>CK>CF>MSC>M,M 处理乙酸发酵产CH4途径比例最低,比其他5个处理平均低52.52%,分别比S、MS、MSC处理低53.49%、61.17%、45.65%。

图3 各处理乙酸发酵途径和CO2还原途径产CH4比例Fig.3 The proportion of CH4 produced by AM and HM of different treatments
2.4 稻田CH4排放通量和田面水溶存CH4浓度
不同处理间CH4排放通量范围为5.70 ~ 26.65 mg/(m2·h),各处理差异不显著,所有施肥处理相比不施肥(CK)均增加了CH4排放(图4)。含秸秆还田的3个有机处理的CH4排放通量均高于CF,而没有秸秆还田的M处理比CF降低了5.91%,相比S、MS、MSC分别低12.50%、15.30%和44.40%。

图4 不同处理稻田CH4排放通量和田面水溶存CH4浓度Fig.4 CH4 flux from rice field and dissolved CH4 concentration from surface water in different treatments
不同处理间田面水溶存CH4浓度范围为70.02~163.58 mg/kg,各处理差异不显著(图4)。MSC处理的田面水溶存CH4浓度最高,比其他处理平均提高30.68%。除MSC处理外,其他施肥处理的溶存CH4浓度均低于CK,相比S和MS处理,M处理的田面水溶存CH4浓度较高。
2.5 δ13C-CH4分布特征
各处理CH4产生、排放和田面水传输过程均呈现出一致的δ13C分馏变化趋势:厌氧培养产生<整体排放<田面水溶存。厌氧培养期为CH4的产生过程,初始期(0 h)各处理δ13C-CH4为-102.73‰~-84.00‰,48 h后为-94.65‰~-72.53‰,δ13C-CH4呈富集趋势,这可能是由于发生了一定厌氧氧化,消耗了不稳定的12C,使δ13C相对富集(图5A)。田面水溶存δ13C-CH4值相对产生过程较富集,分布范围也较大(-57.66‰~-20.24‰) (图5B),这说明经过土壤氧化及田面水中的好氧氧化,发生了同位素分馏和δ13C-CH4富集。田间整体排放的δ13C-CH4值(-66.38‰~-61.83‰)同样相对产生过程较富集(图5B),说明土壤产生的CH4经过氧化和传输过程的同位素分馏,最终排放出的12C-CH4较产生过程呈贫化的趋势。

图5 不同处理CH4产生(A)、排放和田面水溶存(B)过程的δ13C-CH4特征Fig.5 Characteristics of δ13C-CH4 of production (A), emission and soluble state (B) of different treatments
2.6 环境要素对CH4产生和排放的影响
各处理甲烷产生、排放与各环境要素间的相关性分析表明,CH4排放通量与CH4产生潜力呈正相关,与CH4氧化潜力呈负相关,CH4产生与排放受到土壤理化因子的综合影响(图6)。CH4排放通量、CH4产生潜力与土壤TDN含量呈显著正相关(P<0.05),各处理TDN含量大小顺序为MSC>S>MS>CF>M>CK (表1),所有施肥处理TDN含量高于CK,与CH4排放通量规律一致。CH4产生潜力与土壤DOC含量呈显著正相关(P<0.05),其中M处理的DOC含量分别比S、MS、MSC处理低25.99%、1.54%、21.18%,与CH4产生潜力变化趋势一致。另外,土壤C/N与CH4排放通量、CH4产生潜力均呈负相关性,各处理土壤C/N大小顺序为M>CK>MS>S>MSC>CF (表1)。综上可见,土壤TDN、DOC和C/N与CH4产生和排放过程密切相关。
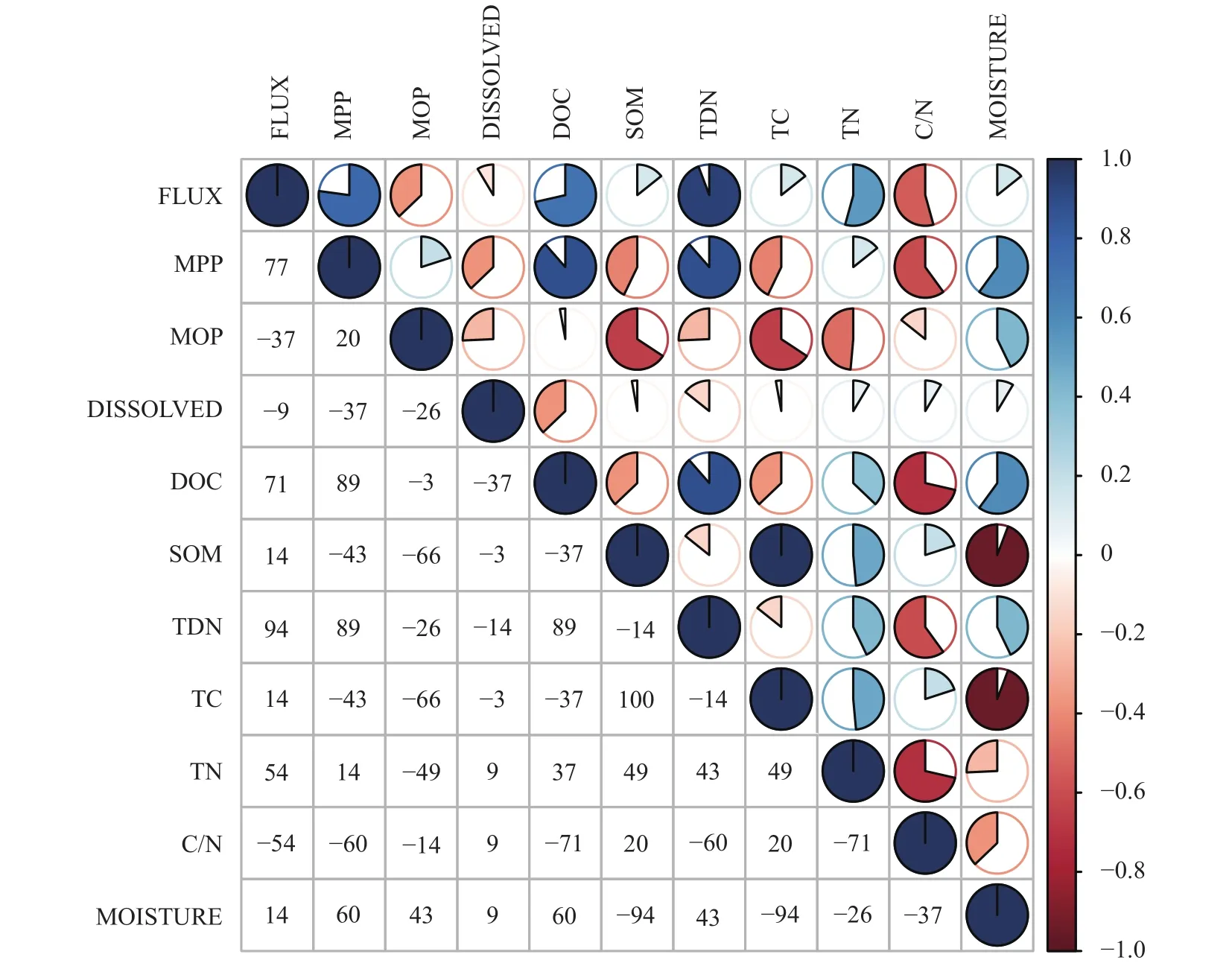
图6 各观测变量相关矩阵Fig.6 Correlation among different measurement indexes
3 讨论
3.1 冬种紫云英减施化肥与CH4减排效应
绿肥是农作物生产的重要物质基础,是绿色农业的有效技术支撑,数据表明,绿肥能有效促进作物养分供给、保障作物稳产高产、全面改善土壤物理、化学、生物性状以及减少温室气体排放[27]。本研究结果表明,冬种紫云英具有较好的氮肥减施和甲烷减排效应。相比单施化肥(CF)处理,化肥+紫云英全量还田(M)减施化肥N 57.01 kg/hm2,节省氮肥27.15% (图7),而相比化肥+秸秆全量还田(S)处理,也能减施13.78%的N,折合化肥N 24.46 kg/hm2。另外,M处理的CH4排放通量低于CF和其他几种组合处理,尤其比S处理低12.50%,表明冬种紫云英可在替代氮肥的同时减少超级稻田CH4排放。田卡等[8]对华南双季稻田的研究表明,等化肥投入下秸秆还田相比单施化肥提高了水稻幼穗分化前稻田CH4排放通量,而冬种绿肥影响不大。张丽[28]在广州双季稻田的研究发现,等化肥投入下秸秆还田比冬种紫云英提高了早稻和晚稻CH4排放通量,但相比单施化肥,紫云英、秸秆还田及二者协同还田均显著增加了双季稻CH4排放。另外,刘威[29]研究了秸秆不同还田方式对江汉平原单季中稻CH4排放通量的影响,发现化肥+秸秆覆盖还田、化肥+紫云英还田处理的CH4排放通量高于不施肥、单施化肥和化肥+秸秆焚烧还田处理。这些研究结果的差异可能是由于研究区域气候、土壤质地、施氮量、研究年限等不同导致的,另外,以往研究多是基于等氮肥基础上添加不同有机物料,这与本研究等养分投入的设定有差异。
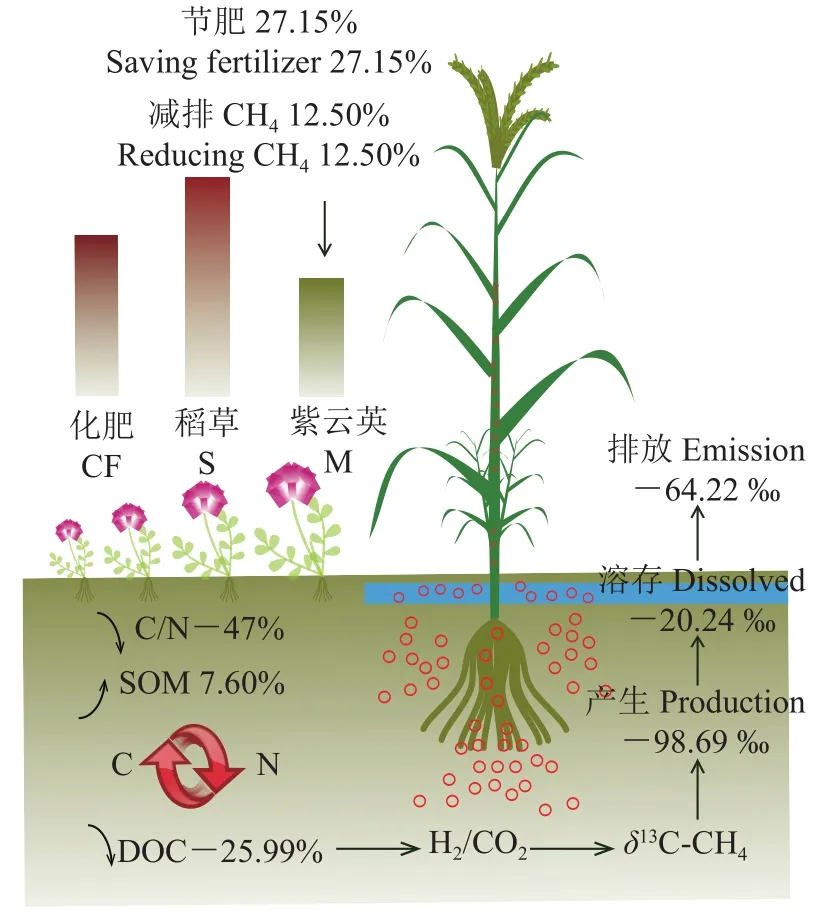
图7 冬种紫云英节肥和CH4减排效应示意图Fig.7 Schematic diagram of the effects of winter-planting Chinese milk vetch on nitrogen input reduction and CH4 mitigation
本研究发现,化肥+紫云英全量还田(M)处理的CH4产生潜力最低,比其他5个处理平均低61.04%,分别比CF和S两个处理的CH4产生潜力低了62.29%和83.16%,表明了冬种紫云英有较强的CH4产生抑制效应。紫云英还田也抑制了部分CH4氧化能力,相比CF和S处理,CH4氧化潜力分别降低了4.53%和5.36%但差异不显著(P>0.05),这说明单独紫云英还田对土壤微生物群落特别是产甲烷菌和嗜甲烷菌分别具有抑制和激发效应,但对CH4产生的抑制明显强于对CH4氧化过程的激发,因此最终显示出对稻田甲烷整体排放的抑制效应。Zhou等[30]的培养试验得出相似的结论,发现添加秸秆残茬的处理CH4产生潜力和氧化潜力均高于添加绿肥残茬处理。实际上,绿肥与稻秸投入直接改变了土壤养分状况,间接改变了产甲烷菌和嗜甲烷菌群落结构和活性,进而改变了土壤CH4产生和氧化潜力,最终导致CH4排放差异[30-31]。本研究相关分析表明,CH4排放通量与CH4产生潜力呈较明显正相关性,但与CH4氧化潜力负相关性不明显。这与前人研究结论相似,如Zhou等[30]研究表明,CH4产生潜力而非CH4氧化潜力对CH4排放具有很强的直接影响。
3.2 冬种紫云英CH4减排机制
由于紫云英植株C/N值显著低于水稻植株,在本研究中,紫云英全量还田处理(M)显著增加了土壤有机质和总碳含量,比秸秆全量还田(S)处理分别提高7.60%和7.55% (表1),但该处理显著降低了土壤活性有机碳(DOC)的含量,相比CF和S处理分别降低了17.45%和25.99%,这就从碳源上抑制了产CH4过程,而更多的碳则储存于土壤中。本研究的相关分析表明,各处理CH4产生潜力与土壤DOC含量呈显著正相关,进一步证实了紫云英对于土壤碳周转和CH4产生与排放的影响。多位学者研究证明CH4排放与土壤DOC呈正相关,DOC增加促进CH4排放,原因在于淹水条件下壮大的厌氧及兼性厌氧微生物群落能够对土壤DOC进行无氧发酵,从而增加土壤CH4产生与排放。另外,研究表明,随有机物料残茬C/N增加,土壤CH4排放随之增大[32-35],因此本研究中不同C/N的紫云英和秸秆因自身分解差异导致土壤DOC含量差异,进而造成紫云英比秸秆还田显著抑制稻田CH4产生与排放,原因可能在于本试验中紫云英于水稻移栽前两个月翻压还田,经过充分的好氧预分解过程促进了土壤活性有机碳彻底分解为CO2,导致在水稻移栽之后的淹水土壤中紫云英处理的土壤可利用碳较低,CH4产生被强烈抑制;相比水稻秸秆,紫云英C/N明显更低(低47.04%,表2),水稻秸秆含有木质素等更多难以降解的物质,分解更缓慢且不完全,在试验淹水期间土壤DOC含量高于其他处理,刺激了CH4产生。
此外,本研究还发现,CH4排放通量、CH4产生潜力也与土壤可溶性总氮(TDN)含量呈显著正相关,所有施肥处理CH4排放通量均高于不施肥处理(CK),而且各处理相比CK均提高了土壤TDN含量,Yu等[36]研究也发现有机物料连续施用10年的稻田土壤TDN显著增加。相比化肥+秸秆全量还田处理(S),化肥+紫云英全量还田处理(M)的总氮含量增加了1.82%,而TDN含量下降了19.95% (表1),可见冬种并翻压紫云英降低了水稻生育早期土壤有效态氮,可能减少了产甲烷菌氮源进而抑制其生长和活性,从而抑制了CH4产生与排放。
本研究发现,土壤C/N与CH4通量和CH4产生潜力均呈负相关性,化肥+紫云英还田提高了土壤C/N (促进提高了碳固存),抑制了CH4产生与排放,反之,化肥+秸秆还田降低了土壤C/N,促进了CH4产生与排放。这可能是由于低C/N的紫云英还田后易于被微生物消耗分解,短期内有多余的氮进行氮矿化,有利于土壤中可利用氮的增加,可直接为植物所吸收,土壤C/N相对提高。高C/N的秸秆含氮量低,等氮条件下比紫云英还田需配施更多氮肥,氮输入能降低土壤C/N,促进土壤有机质分解,增加土壤活性碳含量,促进CH4产生。
另外,本研究发现化肥+秸秆和紫云英全量还田+熟石灰处理(MSC)的稻田CH4排放通量和田面水溶存CH4浓度最高(图4),且CH4产生潜力仅低于S处理。研究表明,土壤pH可能通过影响有机质分解、土壤C/N和CH4产生过程间接影响CH4排放[37-38]。一方面MSC处理的土壤DOC含量较高,土壤C/N较低(表1),可能因此促进了稻田土壤CH4产生与排放(图6);另一方面土壤pH的轻微变化会影响土壤CH4排放,尤其是厌氧条件下CH4产生对土壤pH值变化非常敏感,中性或微碱性土壤最有利于CH4产生[38],因此添加熟石灰可能因土壤pH升高而促进了产CH4菌活性。
本研究中,培养试验土壤取自水稻分蘖初期,研究发现各处理CH4产生过程以H2/CO2还原为主。一些研究也发现冬闲期和水稻生长季大部分时期H2/CO2还原产生途径非常重要(约50%),仅在7、8月水稻生长旺盛时期,乙酸发酵途径占主导地位(约70%)[39-40]。我们也发现,随培养时间延长,乙酸发酵过程的贡献占比增加(图3)。本研究中,乙酸发酵产CH4途径比例平均值[为减少时间误差,本研究中乙酸发酵产CH4途径贡献率采用厌氧培养0和48 h的平均值(图3)来计算]与相应δ13C-CH4平均值(图5A)呈极显著正相关(r=0.943,P<0.01),这可能是由于利于乙酸发酵产CH4的环境中12CH4较快氧化,导致δ13C-CH4值偏正,反之厌氧培养的δ13CCH4值偏负,从而说明此环境利于H2/CO2还原产CH4。另有研究认为,晒田会导致土壤氧化还原电位偏高,乙酸型产甲烷作用在很大程度上优于H2/CO2还原型产甲烷作用[41],这说明氧化作用会使δ13C-CH4值偏正,环境中乙酸型产CH4途径比例增加,氢营养型产CH4途径比例下降,这与本研究的结果一致。此外,本研究发现乙酸发酵产CH4途径比例平均值(图3)与CH4产生潜力(图2A)呈极显著正相关(r=0.628,P<0.01),表明乙酸发酵途径对CH4产生总量的重要影响,厌氧培养期间紫云英全量还田处理乙酸发酵产CH4途径贡献率最低,平均只有16.11%,而秸秆全量还田处理占到34.65%,表明紫云英单独还田抑制了乙酸发酵产CH4途径(相比秸秆单独还田降低53.49%),从而抑制了CH4产生,而水稻秸秆还田可明显促进乙酸发酵过程,这可能是由于介导两种途径的功能微生物类群的差异。
4 结论
甲烷排放通量和产生潜力与土壤可溶性有机碳、可溶性总氮含量呈正相关,与土壤碳氮比呈负相关。在等氮磷钾投入条件下,冬种并翻压紫云英减少了化学氮肥投入,降低了土壤可溶性有机碳含量,提高了土壤碳氮比,因而有效抑制了稻田土壤甲烷的产生和排放。秸秆的作用正好相反,因此,秸秆还田需配合紫云英翻压来抵消甲烷的产生和排放。
厌氧条件下,乙酸发酵产甲烷途径的比例随培养时间延长而升高。厌氧培养的乙酸发酵产甲烷途径比例平均值与甲烷产生潜力呈正相关,紫云英全量还田处理主要抑制了乙酸发酵这一关键产甲烷过程,从而减少了甲烷产生,其乙酸发酵产甲烷途径比例和甲烷产生潜力比其他5个处理平均降低52.52%、61.04%,其中比秸秆单独还田处理降低53.49%、83.16%。
猜你喜欢
诗歌月刊(2024年1期)2024-02-05 01:36:51
中学生数理化·高一版(2022年4期)2022-05-09 15:36:00
中学时代(2018年6期)2018-11-20 05:53:20
海峡姐妹(2018年7期)2018-07-27 02:30:16
知识经济·中国直销(2018年1期)2018-01-31 01:52:45
中国卫生(2016年7期)2016-11-13 01:06:32
儿童故事画报(2016年4期)2016-06-24 07:58:59
儿童故事画报(2016年4期)2016-06-24 07:57:36
浙江大学学报(工学版)(2016年9期)2016-06-05 09:20:57
当代化工研究(2016年5期)2016-03-20 16:21:29
