
玉米大斑病菌犬尿氨酸途径关键酶基因的鉴定
2022-09-14 04:04:54曹丽萍刘英姿张福源邢继红董金皋
河北农业大学学报 2022年4期
李 白,曹丽萍,刘英姿,张福源,张 康,邢继红,董金皋
(1.省部共建华北作物改良与调控国家重点实验室,河北 保定 071000;2.河北省植物生理与分子病理学重点实验室,河北 保定 071000;3.黑龙江省农业科学院 齐齐哈尔分院,黑龙江 齐齐哈尔 161000;4.河北省承德市园林管理中心,河北 承德 067000)
玉米大斑病(NCLB)是世界范围内玉米上的重要叶部病害,其病原菌为玉米大斑病菌(Setosphaeria turcica),该病菌侵染玉米叶部组织并诱导其坏死病变,严重影响叶片的光合作用,造成产量损失[1]。因此,挖掘玉米大斑病菌的生长发育和致病相关的基因,深入研究其作用机制将为玉米大斑病的防控提供理论依据。
犬尿氨酸途径(Kynurenine pathway,KP)是色氨酸分解代谢的主要方式[2-3],从酵母到人类的整个真核生物谱系中高度保守,能够影响多种重要的生物过程[4]。色氨酸在肝脏中由色氨酸-2, 3-双加氧酶(Tryptophan-2, 3-dioxygenase, TDO)或者其它组织里的吲哚-2, 3-双加氧酶(Indoleamine-2,3-dioxygenase, IDO)的作用下生成N-甲酰基犬尿氨酸(N-Formylkynurenine),犬尿氨酸甲酰胺酶(Kynurenine formamidase, KFA)将其水解后,得到犬尿氨酸(Kynurenine)。犬尿氨酸位于该途径的一个重要的分支位点,依赖于NADPH,并能在犬尿氨酸上发挥羟基化,最后产生3- 羟基犬尿氨酸[5-6]。
犬尿氨酸途径(KP)在以烟酰胺腺嘌呤二核苷酸(NAD+)的形式产生细胞能量方面起着关键作用[7]。NAD(P)+是所有生命体内氧化还原反应的辅助因子,同时也在DNA 修复和组蛋白去乙酰化过程中起重要作用[8-9]。在许多微生物中,犬尿氨酸途径是其NAD+生物合成的唯一来源[10]。在哺乳动物中,犬尿氨酸单加氧酶(KMO)作用于犬尿氨酸,生成具有神经毒性的3-羟基犬尿氨酸,与多种神经退行性疾病密切相关。而在细菌和部分真菌体内,犬尿氨酸途径与真核生物类似,不同的就是犬尿氨酸酶替换了真核生物中的犬尿氨酸单加氧酶,由犬尿氨酸生成邻氨基苯甲酸和L-丙氨酸。邻氨基苯甲酸被氧化酶氧化成邻苯二酚,经过多步反应,最终转变成CO2[11]。玉米大斑病菌犬尿氨酸途径以及此途径中的关键酶基因的功能都未见报道。本研究利用生物信息学方法,对玉米大斑病菌犬尿氨酸途径中的关键酶基因进行系统分析,确定玉米大斑病菌犬尿氨酸途径的关键酶基因;利用玉米大斑病菌的表达谱数据,确定犬尿氨酸途径关键酶基因在病菌生长发育和侵染过程中的表达规律,为阐明玉米大斑病菌犬尿氨酸途径及其途径关键酶基因在病菌生长发育和致病过程中的功能奠定基础。
1 材料和方法
1.1 玉米大斑病菌犬尿氨酸途径关键酶基因序列的获得
利用人和小鼠的犬尿氨酸途径关键酶基因的氨基酸序列,从Ensembl fungi(http://fungi.ensembl.org/index.html)中用Blast 搜索得到玉米大斑病菌(Setosphaeria turcica)中的同源序列。
1.2 犬尿氨酸途径关键酶基因的系统进化分析
利用人类和小鼠的犬尿氨酸单加氧酶(Kynurenine 3-monooxygenase,KMO)、犬尿氨酸酶(Kynureninase,KYN)、吲哚-2, 3-双加氧酶(Indoleamine-2, 3-dioxygenase,IDO)、犬尿氨酸氨基转移酶(Kynurenine aminotransferase,KAT)的氨基酸序列,在Ensembl Fungi 真菌数据库中搜索玉米大斑病菌的全基因组序列;对搜索的结果进行深入的分析,并利用MEGA 7.0 软件构建系统发育树,进行基因的多序列比对,并用邻接法进行系统进化分析,设置 bootstrap 值为 1 000。
1.3 犬尿氨酸途径关键酶基因的结构域分析
利用玉米大斑病菌犬尿氨酸途径关键酶基因StKMO、StKYN、StIDO、StKAT的氨基酸序列,通过InterProScan(http://www.ebi.ac.uk/InterProScan/)网站,对其功能域进行分析,用IBS 软件进行结构域的绘制。
1.4 犬尿氨酸途径关键酶基因在玉米大斑病菌生长发育不同时期的表达分析
从公共平台GEO(https://www.ncbi.nlm.nih.gov/geo/),获取玉米大斑病菌生长发育不同时期的表达谱数据,数据编号为SRR9997710、SRR9997709、SRR9997708、SRR9997715 和SRR9997714;对犬尿氨酸途径关键酶基因StKMO、StKYN、StIDO、StKAT在病菌生长发育的不同时间的数据进行分析,取log2 值后确定目的基因在病菌不同生长发育时期的表达水平。
1.5 犬尿氨酸途径关键酶基因在玉米大斑病菌侵染过程中的表达分析
从公共平台GEO(https://www.ncbi.nlm.nih.gov/geo/),获取玉米大斑病菌侵染过程中的表达谱数据,数据编号为SRR12131452、SRR12131453、SRR12131454 和SRR12131455;对犬尿氨酸途径关键酶基因StKMO、StKYN、StIDO、StKAT在病菌侵染不同时间的数据进行分析,取log2 值后确定目的基因在病菌不同侵染时间的表达水平。
2 结果与分析
2.1 犬尿氨酸单加氧酶(KMO)的系统发育树和功能域分析
利用人类和小鼠的犬尿氨酸单加氧酶(KMO)的氨基酸序列在Ensembl Fungi 真菌数据库中搜索玉米大斑病菌的氨基酸序列,使用MEGA 7.0 软件构建系统发育树,发现SETTUDRAFT_165798与其他物种的KMO 相似性较高(图1A);利用InterProScan[12]网站对其功能域进行分析,发现它们都具有犬尿氨酸单加氧酶(kynurenine 3-monooxygenase) 的功能域( 图1B)。 表明SETTUDRAFT_165798 为玉米大斑病菌的犬尿氨酸单加氧酶(KMO)基因,命名为StKMO。

图1 犬尿氨酸单加氧酶KMO 的系统发育树和功能域分析Fig.1 Phylogenetic tree and functional domain analysis of kynurenine 3-monooxygenase
2.2 犬尿氨酸酶(KYN)的系统发育树和功能域分析
利用人类和小鼠的犬尿氨酸酶(KYN)的氨基酸序列在Ensembl Fungi 真菌数据库中搜索玉米大斑病菌的氨基酸序列,使用MEGA 7.0 软件构建系统发育树,发现SETTUDRAFT_165216 与其他物种的KYN 相似性较高(图2A);利用InterProScan网站对其功能域进行分析,发现它们都具有一个犬尿氨酸酶(Kynureninase)的功能域,并具有高度相似性(图2B)。表明SETTUDRAFT_165216 为玉米大斑病菌的犬尿氨酸酶(KYN)基因,命名为StKYN。

图2 犬尿氨酸酶KYN 的系统发育树和功能域分析Fig.2 Phylogenetic tree and functional domain analysis of kynureninase
2.3 犬尿氨酸氨基转移酶(KAT)的系统发育树和功能域分析
利用人类和小鼠的犬尿氨酸氨基转移酶(KAT)的氨基酸序列在Ensembl Fungi 真菌数据库中搜索玉米大斑病菌的氨基酸序列,使用MEGA 7.0 软件构建系统发育树,发现SETTUDRAFT_177407与其他物种的KAT 相似性较高(图3A);利用InterProScan 网站对其功能域进行分析,发现它们都具有一个犬尿氨酸氨基转移酶(Kynurenine aminotransferase) 的功能域( 图3B)。 表明SETTUDRAFT_177407 为玉米大斑病菌犬尿氨酸氨基转移酶(KAT)基因,命名为StKAT。

图3 犬尿氨酸氨基转移酶KAT 的系统发育树和功能域分析Fig.3 Phylogenetic tree and functional domain analysis of kynurenine aminotransferase
2.4 吲哚-2, 3-双加氧酶(IDO)的系统发育树和功能域分析
利用人类和小鼠的吲哚-2, 3-双加氧酶(IDO)的氨基酸序列在Ensembl Fungi 真菌数据库中搜索玉米大斑病菌的氨基酸序列,系统发育分析发现SETTUDRAFT_102413、SETTUDRAFT_171749与其他物种的IDO 相似性较高(图4A);利用InterProScan 网站对其功能域进行分析,发现它们都具有吲哚-2, 3- 双加氧酶(Indoleamine-2,3-dioxygenase)的功能域,并具有高度相似性(图4B)。表明SETTUDRAFT_102413、SETTUDRAFT_171749 为玉米大斑病菌的吲哚-2, 3-双加氧酶(IDO)基因,命名为StIDOs。

图4 吲哚-2, 3-双加氧酶IDO 的系统发育树和功能域分析Fig.4 Phylogenetic tree and functional domain analysis of indoleamine-2, 3-dioxygenase
2.5 犬尿氨酸途径关键酶基因在玉米大斑病菌生长发育不同时期的表达分析
利用公共平台GEO 中玉米大斑病菌的表达谱数据,对犬尿氨酸途径关键酶基因StKMO、StKYN、StKAT、StIDOs在病菌的附着孢时期、萌芽时期,早萌时期以及分生孢子发育时期的数据进行分析。结果发现,StKMO基因在病菌萌芽和分生孢子发育时期高水平表达,在附着胞时期和早萌时期表达相对较低;StKYN基因在病菌生长发育的4 个时期均呈现高水平表达;StKAT基因在病菌分生孢子发育时期高水平表达,在附着胞时期、萌芽时期和早萌时期表达相对较低;StIDO-1(SETTUDRAFT_102413)基因在病菌生长发育的4 个时期均呈现高表达,StIDO-2(SETTUDRAFT_171749)基因在病菌萌芽时不表达,其他时间均表达(见图5)。
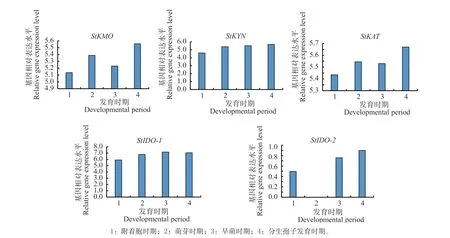
图5 犬尿氨酸途径关键酶基因在玉米大斑病菌生长发育中的表达Fig.5 Expression of key enzyme genes of the kynurenine pathway in growth and development of Setosphaeria turcica
2.6 犬尿氨酸途径关键酶基因在玉米大斑病菌侵染过程中的表达分析
利用公共平台GEO 中玉米大斑病菌的表达谱数据,对犬尿氨酸途径关键酶基因StKMO、StKYN、StKAT、StIDOs在病菌侵染的3、5、9、13 d 的数据与对照(0 d)进行对比分析。结果发现,玉米大斑病菌StKMO基因在病菌侵染3、9 和13 d 时高水平表达,在侵染0 d 和5 d 时不表达;StKYN基因和StKAT基因在病菌侵染9 d、13 d 时高水平表达,在病菌侵染的前5 d 均不表达;StIDOs(SETTUDRAFT_102413 和SETTUDRAFT_171749)基因只在病菌侵染9 d 时表达,其他时间均不表达(见图6)。

图6 犬尿氨酸途径关键酶基因在玉米大斑病菌侵染过程中的表达Fig.6 Expression of key enzyme genes of kynurenine pathway in infection of Setosphaeria turcica
3 结论与讨论
在哺乳动物中,KMO 作用于犬尿氨酸,使其转变为具有神经毒性的3-羟基犬尿氨酸,与亨廷顿病(Huntington’s, HD)、阿尔茨海默病(Alzheimer’s, AD)和帕金森病(Parkinson’s, PD)等多种神经退行性疾病密切相关[13-16]。研究报道,细菌以及部分真菌中也存在与哺乳动物相似的犬尿氨酸途径。如,铜绿假单胞菌(Pseudomonas aeruginosa)通过犬尿氨酸途径分解代谢色氨酸[17]。玫瑰孢链霉菌(Streptomyces roseosporus)达托霉素生物合成基因簇中含有色氨酸2, 3-双加氧酶基因dptJ,dptJ基因敲除后,达托霉素的产量显著降低;将玫瑰孢链霉菌中的TDO 敲除后,达托霉素的产量也显著降低;分别遗传回补后产量得到增加,表明犬尿氨酸途径在达托霉素生物合成中的重要作用[18]。相关研究发现,在人体炎症条件下,犬尿氨酸途径中的关键酶,特别是犬尿氨酸3-单加氧酶(KMO)会有明显的上调[19]。
目前,玉米大斑病菌犬尿氨酸途径及其途径关键酶基因的功能均未见相关报道。本研究通过对Ensembl fungi 数据库进行搜索,获得了玉米大斑病菌中犬尿氨酸途径关键酶的编码基因,包括1 个犬尿氨酸单加氧酶(KMO)的编码基因StKMO、1 个犬尿氨酸酶(KYN)的编码基因StKYN、1 个犬尿氨酸氨基转移酶(KAT)的编码基因StKAT以及2 个吲哚-2, 3-双加氧酶(IDO)的编码基因StIDOs。利用生物信息相关方法对玉米大斑病菌的犬尿氨酸途径关键酶基因进行了系统发育分析和保守结构域分析,证明了玉米大斑病菌犬尿氨酸途径的存在。利用玉米大斑病菌的转录组数据,对犬尿氨酸途径关键酶基因StKMO、StKYN、StKAT和StIDOs在病菌生长发育和侵染过程中的表达规律进行分析,发现这些基因在病菌不同生长发育和侵染时期呈现高水平表达,推测这些基因在玉米大斑病菌生长发育以及侵染过程中发挥重要功能。但是玉米大斑病菌犬尿氨酸途径及其关键酶基因在病菌生长发育和致病过程中功能以及作用机制还需进一步的深入研究。
猜你喜欢
幼儿100(2023年18期)2023-05-29 08:34:36
课堂内外·小学版(低年级)(2022年4期)2022-06-24 09:47:10
当代水产(2022年1期)2022-04-26 14:35:30
世界科学技术-中医药现代化(2021年5期)2021-11-05 06:56:34
孩子(2019年8期)2019-08-26 05:43:32
广东饲料(2016年7期)2016-12-01 03:43:36
西南农业学报(2016年5期)2016-05-17 05:42:33
西南农业学报(2016年6期)2016-04-16 05:12:51
江苏调味副食品(2015年1期)2015-02-28 01:56:34
聪明泉·少儿版(2014年2期)2014-09-22 09:54:50
