
玉米杂交种及其亲本苗期性状对低氮胁迫的动态响应
2022-09-13 07:39徐世英冯万军
作物杂志 2022年4期
徐世英 王 宁 程 皓 冯万军
(山西农业大学农学院/黄土高原特色作物优质高效生产省部共建协同创新中心,030801,山西晋中)
杂种优势(hybrid vigor或heterosis)是指2个遗传背景不同的亲本杂交产生的杂种F1代,在生物量、生长速率、生活力及在生物或非生物胁迫的抗性等性状上优于亲本的现象[1-2]。目前,常用于评估特定性状杂种优势表现的指标包括中亲优势(MPH)和超亲优势(BPH),即分别为同一性状杂交种与亲本中亲值(MPV)和高亲值差值的比值[3]。杂种优势在玉米生产上已经广泛应用,但其遗传基础尚不清楚。研究表明,玉米在众多性状上均有杂种优势表现,如生物量或产量等成株性状[4-6],以及胚[7-8]和幼苗[9-10]发育早期的多个性状。玉米幼苗初生根和种子根对幼苗早期活力起关键作用,因此其幼苗根系是研究杂种优势遗传基础的理想材料[11]。Hoecker等[10]对12个玉米杂交种及其亲本自交系研究发现,不同幼苗根系性状的杂种优势表现有较大差异,其中初生根长度、侧根密度和种子根数等性状优势值较高。
杂种优势表现与遗传、环境和生长发育时期等众多因素有关[12]。玉米杂交种比亲本生长更快、产量更高,这可能与其具有更优异的营养环境适应能力有关[13],也因此比亲本更耐密植[14]。研究发现,在干旱胁迫下,耐旱性强的玉米杂交种及其亲本体内的抗氧化酶活性升高幅度大于耐旱性弱的玉米杂交种及其亲本,杂交种的耐旱性受亲本的加性效应作用[15],但随着干旱时间推进,杂交种及其亲本体内的抗氧化酶活性有一定变化,母本对杂交种的保护酶活性和抗膜脂过氧化伤害能力的影响较大[16]。在低磷胁迫下,玉米自交系幼苗体内的超氧化物歧化酶(SOD)活性下降,而在杂交种中却升高,杂交种在多个性状上都有比亲本更强的耐低磷性[17]。陈范骏等[18]研究指出,在低氮和高氮水平下,玉米杂交种的氮素利用效率(NUE)均有中亲或超亲优势,且在低氮水平下玉米杂交种的NUE仅受母本影响。在不施氮的情况下,玉米杂交种在生殖生长期的氮素吸收量与总氮素吸收量的比值相较于亲本显著增加[19]。这些研究表明,在逆境下,杂交种具有比亲本更好的耐受性,但杂种优势表现因遗传背景和逆境下生长时间不同而存在较大差异。
氮素是保证作物产量的重要矿质元素之一[20-21]。在农业生产中,由于氮肥的过量施用,作物的产量并未持续提高,并且导致一系列严重的环境问题[22-23]。因此,如何在减少氮肥用量的同时保证作物产量是目前农业生产面临的主要矛盾之一,而解析作物的氮素利用机制,培育氮高效利用的作物新品种,可能是解决这一矛盾较为有效的手段[24]。目前,关于缺氮条件下杂交种及其亲本苗期性状如何进行动态响应,且苗期杂种优势的形成是否受到影响的研究未见报道。为此,本研究采用水培法,通过设置低氮和正常供氮2个处理,测定处理不同时间后2个玉米杂交种及其亲本幼苗期地上部和根系性状,分析其杂种优势表现的动态变化,为解析玉米杂种优势形成机制以及氮高效玉米杂交种选育提供理论参考。
1 材料与方法
1.1 试验方法
以玉米杂交种豫玉22(YY22)和先玉335(XY335)及其亲本自交系[豫玉22母本综3(Z3)、父本87-1,先玉335母本PH6WC、父本PH4CV]为试验材料,于2020年在山西农业大学农作站经过严格的人工套袋授粉繁育,选取饱满的种子在10%双氧水中消毒30min,用蒸馏水冲洗干净后用蒸馏水浸泡12h,后转移到含有双层滤纸的培养皿中,放入25℃培养箱中进行催芽,待初生根长至约1.0cm,挑选出长势均匀的幼苗,去除胚乳后全部移植到装有44L含1/2营养液的黑色育苗箱中,置于人工气候室进行蹲苗处理,待第2个叶片完全展开,挑选长势一致的幼苗,将其随机分为2组,分别移植到盛有正常氮素(2mmol/L,CK)和低氮(0.04mmol/L,LN)水平营养液的黑色育苗箱中进行培养,3次重复。营养液配方参考赵泽群等[25]的方法,氮用Ca(NO3)2·4H2O供给,并以1.8mmol/L CaCl2·2H2O补充不足的Ca2+,营养液pH设为6.0±0.1,每天进行标定,每3d换1次营养液,同时随机更换水培箱的位置。在幼苗培养过程中,人工气候室昼夜光照长短及温度分别设定为16h/8h和26℃/18℃,光照强度14 000lx,期间用电动气泵连续通气。分别在培养3、7和14d时,3次重复中各选取5株长势一致幼苗用于表型和全氮含量测定。
1.2 测定项目与方法
1.2.1 表型性状 用解剖刀截取幼苗的第1(L1)和第2(L2)片叶片,用扫描仪Epson V850扫描拍照,利用ImageJ软件分析叶面积。
将地上部与根系手动分离,根系用水清洗,把每个材料的根系放入装有双蒸水的有机玻璃盘(30cm×20cm)中,逐一分开侧根,防止重叠,然后进行扫描。利用WinRHIZOTM软件分析单株总根长、根体积、根表面积、根平均直径、分枝数和根尖数等根系性状[26]。测定完成的根系与地上部分开放置于烘箱中120℃杀青30min,80℃烘至恒重,最后称干重。并计算根冠比,根冠比=根系干重/地上部干重。
1.2.2 生理指标 在取材当天,利用叶绿素仪(SPAD-502,Minolta Camera Co.Ltd.,日本)测定幼苗的第1叶(L1)和第2叶(L2)的相对叶绿素含量(SPAD值)。
地上部和根系样品烘干研磨后,称取50mg粉末,采用H2SO4-H2O2-靛酚蓝比色法测定氮素浓度[27]。根据公式:氮素积累量(mg)=生物量×氮素浓度,计算地上部、根系、整株的氮素含量及地上部与根系中氮与整株氮含量之比;根据公式:耐低氮指数=低氮处理下性状值/对照性状值[28],计算各性状的耐低氮指数。
1.2.3 杂种优势指标 用以下公式[3]计算各性状的中亲优势和超亲优势:中亲优势(MPH,%)=[F1-双亲均值(MP)]/MP×100;超亲优势(BPH,%)=[F1-高亲值(HP)]/HP×100。
1.3 数据处理
用Excel 2010进行数据整理,利用SPSS 16.0软件对整理好的数据进行统计分析,用TBtools和Origin 2021软件进行聚类分析。
2 结果与分析
2.1 玉米杂交种及其亲本幼苗叶片和根系形态对低氮胁迫的反应差异
在LN处理7d时,6个基因型的L1和L2尚未明显衰老,但在LN处理14d时,先玉335及其亲本的L1和L2均已萎蔫,且以先玉335萎蔫程度更为严重;而在LN处理14d时,豫玉22及其亲本仅L1衰老严重,其L2尚未明显衰老,且在3个基因型间差异不明显(图1a)。在根系形态上,LN处理3d时,豫玉22根系形态与对照相比已有明显变化,侧根数和侧根长均比对照明显增加,且在7d时这些变化更为明显,而至14d时其生长受到抑制,但其两亲本根系在2个氮素水平处理早期(3和7d)变化并不明显,仅87-1在LN处理14d时其根系才明显比对照更为庞大;与之不同,先玉335及其父本PH4CV的根系形态在LN处理7d时才与对照相比有明显变化,表现为侧根的数目增加,根系更为庞大,而在LN处理14d时与CK处理相比差异不明显,但其母本PH6WC在两氮素水平处理间始终变化并不明显(图1b)。上述结果表明,玉米苗期根系对低氮的反应要早于地上部,且杂交种对低氮的反应比亲本更为迅速,但这种反应在不同基因型上有较大差异,并且会随着时间有一定变化。
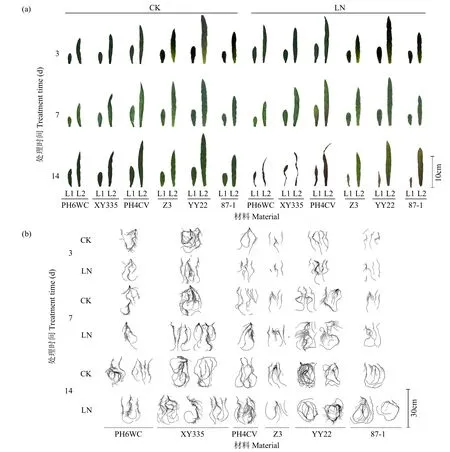
图1 2个氮素水平下玉米杂交种及其亲本幼苗叶片(a)和根系(b)形态的动态变化Fig.1 Dynamic morphology changes on leaves(a)and roots(b)of seedlings among maize hybrids and their parents under two nitrogen levels
2.2 2个氮素水平下玉米杂交种及其亲本幼苗性状的方差分析
从表1可以看出,除根表面积和根体积在氮素水平上差异不显著外,其余15个性状在培养时间、氮素水平和基因型3个因素上均有显著或极显著差异,且仅根平均直径未受到培养时间与氮素水平及3个因素间互作的显著影响,而其余性状在任意2个或3个因素间互作上均有显著或极显著差异。另外,从方差值来看,氮素水平对单株氮积累量、地上部氮积累量、叶片的SPAD值和根系氮积累量等性状影响较大,其次是根冠比、地上部干重和根尖数,而对其余7个性状的影响相对较小,说明不同性状对氮素的反应有一定差异,但这种反应差异受培养时间、基因型及其相互作用的影响。
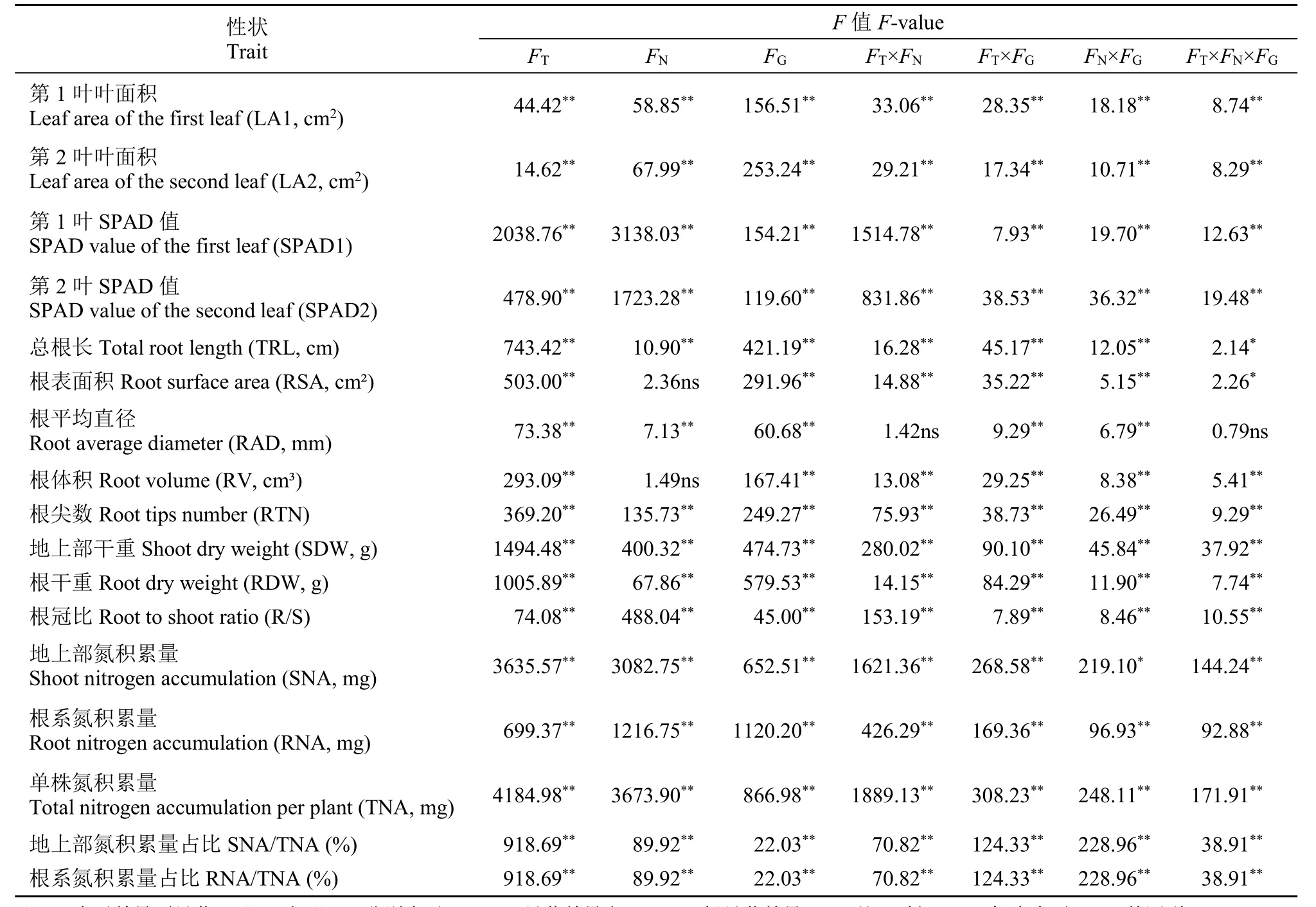
表1 2个氮素水平下不同培养时间玉米杂交种及其亲本幼苗性状的方差分析Table 1 The variance analysis of the seedling traits among maize hybrids and their parental lines at different days after two nitrogen treatments
2.3 玉米杂交种及其亲本幼苗杂种优势及聚类分析
结合杂种优势测定结果与聚类分析(图2)发现,2个杂交种所有性状的中亲和超亲优势在各时间点的变化趋势基本一致,均以总根长、根表面积、根体积、根尖数、根干重、地上部干重、地上部氮素积累量、根系中氮素积累量、单株氮素积累量9个性状在各时间点具有较大中亲和超亲优势值,而L1和L2的叶面积、SPAD值、根平均直径、根冠比、地上部和根系的氮积累量占比等8个性状在各时间点的中亲和超亲优势值相对较小,甚至个别性状表现出负优势,因此根据17个性状的杂种优势表现差异大致可以分为2组。另外,从优势值来看,LN处理7d时,豫玉22根尖数的中亲和超亲优势值最大,分别达到了413.5%和401.2%,其次是CK处理7d的豫玉22的总根长和根表面积,其总根长中亲和超亲优势值分别为383.3%、365.5%,根表面积为342.3%、323.0%;豫玉22其余性状在CK处理7d的中亲优势在-38.2%(7d根系氮素积累量占比)~383.3%(7d总根长),LN处理中亲优势值7~14d在-19.6%(14d的L2的SPAD值)~413.5%(7d根尖数),CK处理7d的超亲优势值为-52.4%(7d根系氮素积累量占比)~365.5%(7d总根长),LN处理7d和14d的超亲值为-27.3%(14d的L2的SPAD值)~401.2%(7d根尖数);而先玉335所有性状在CK处理不同时间的中亲优势则在-28.7%(3d的根系氮素积累量占比)~171.8%(14d的地上部氮素积累量),而LN处理14d时中亲优势值在-71.3%(14d的L2的叶面积)~171.7%(14d的总根长),CK处理3d和14d超亲优势值为-35.4%(3d的根系氮素积累量占比)~129.1%(14d的地上部氮素积累量),LN处理的超亲优势值为-82.7%(14d的L2叶面积)~158.7%(14d总根长)(图2)。
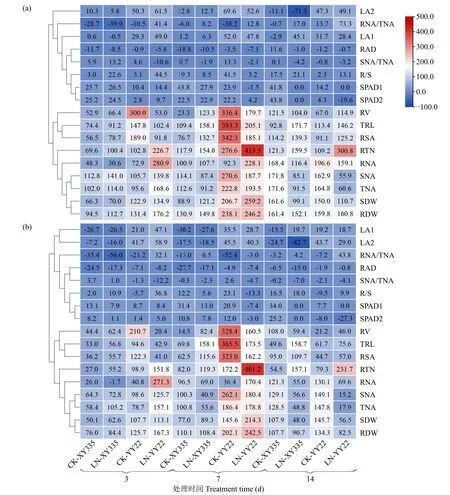
图2 2个氮素水平处理不同天数后玉米幼苗各性状中亲(a)和超亲(b)优势的聚类Fig.2 Cluster of the mid-parent(a)and better-parent(b)heterosis of all traits of maize seedlings at different days after two nitrogen level treatments
由图2可以看出,相比于CK处理,在LN处理下2个杂交种有一些性状的中亲和超亲优势值均升高或降低。其中,在LN处理3d时,先玉335的根体积、总根长、根表面积、根尖数和总氮积累量的增加幅度在11.9%~46.8%,而豫玉22的L1和L2的叶面积、根尖数、根干重、根冠比、地上部氮积累量、根系氮积累量、单株氮积累量及根系氮积累量占比的中亲和超亲优势均升高,增加幅度在11.2%~230.5%;LN处理7d时,先玉335的根体积、总根长、根表面积、地上部干重和根系氮积累量占比的中亲和超亲优势值均提高,增加幅度在12.3%~100.0%,而豫玉22的根系氮积累量、根系氮积累量占比、地上部干重和根尖数的中亲和超亲优势值均升高,增加幅度在为49.4%~229.0%;LN处理14d时,先玉335的总根长、根尖数、根表面积和L1叶面积的中亲和超亲优势值均提高,增加幅度在14.7%~109.1%,而豫玉22的总根长、根表面积、根体积、根尖数、根冠比、根系氮积累量占比的中亲和超亲优势值均升高,增加幅度在10.8%~191.6%。由此表明,豫玉22的根尖数、根系氮积累量占比和先玉335的总根长、根表面积及根尖数的中亲和超亲优势值在3个时间点均比CK有所升高;而在LN处理下,豫玉22的地上部氮积累量和先玉335的叶面积的中亲和超亲优势在3个时间点均比CK有所降低。
综上,在CK和LN处理下,杂交种幼苗的多个性状均存在中亲和超亲优势,且低氮对玉米幼苗的杂种优势表现有一定影响,多个根系性状及其根系中的氮素含量的杂种优势在LN处理下会提高,而叶面积和地上部氮素积累量的杂种优势则会降低,但这种差异表现会因性状、培养时间和杂交组合不同而存在较大变化,由此表明杂种优势的遗传机制异常复杂。
2.4 玉米杂交种及其亲本幼苗性状耐低氮性分析
对各性状的耐低氮指数分析(表2)发现,随着胁迫时间推进,17个性状的耐低氮性上,地上部干重、L1和L2的叶面积和SPAD值、地上部氮积累量、根系氮积累量和总氮积累量呈递减的变化,根冠比和根系氮积累量占比呈递增趋势,总根长、根表面积、根体积、根平均直径、根尖数和根干重先升高后降低,而地上部氮积累量占比则先保持不变后有所降低。同时,基于3个时期和所有基因型,除根平均直径和地上部氮积累量占比整体的变异系数较小以外,其余性状的变异系数均在20.2%~75.0%。另外,随着胁迫时间推进,6个基因型在所有性状上的变异系数呈逐渐升高的变化,说明培养时间对各性状的耐低氮性的影响较大。
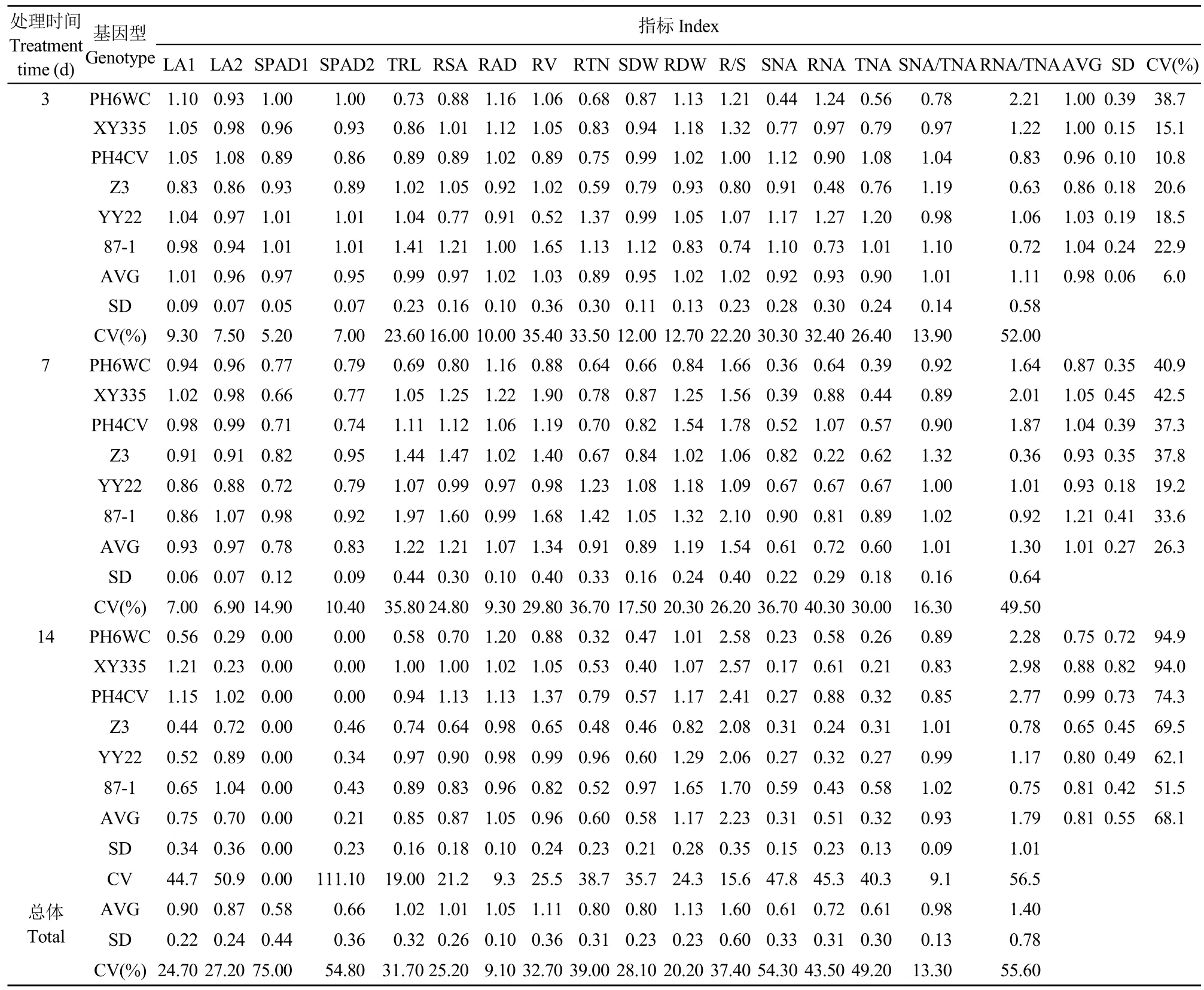
表2 不同培养时间玉米杂交种及其亲本幼苗耐低氮指数Table 2 Low nitrogen tolerance indexes of seedling among maize hybrid and their parents at different days after cultivation
基于各性状的耐低氮指数的平均值,对3个时间点下6个玉米基因型幼苗的耐低氮性进行分析发现,在处理3d时,除综3耐低氮性较差外,其余5个基因型的整体耐低氮性差异不大;在7d和14d,6个玉米基因型的耐低氮性的排序分别为:87-1>先玉335≥PH4CV>综3≥豫玉22>PH6WC和PH4CV>先玉335>87-1≥豫玉22>PH6WC>综3(表2)。综合3个处理时间的结果可以看出,6个基因型的材料耐低氮性依次为87-1(1.02)>PH4CV(0.99)>先玉 335(0.98)>豫玉 22(0.92)>PH6WC(0.87)>综3(0.82)。由此表明,6个基因型玉米的耐低氮能力随着时间推进会发生变化,而各时间点下杂交种的耐低氮能力在2个亲本之间或与其中1个亲本接近。
2.5 基于耐低氮指数对各性状间的相关性分析
相关性分析(表3)表明,根尖数的耐低氮指数与总根长、根表面积、根系氮积累量和单株氮积累量等性状的耐低氮指数呈显著或极显著正相关;而根冠比的耐低氮指数与根系氮积累量占比的耐低氮指数呈极显著正相关,但与L1和L2的SPAD值、地上部干重、地上部氮积累量、单株氮积累量、地上部氮积累量占比等性状的耐低氮指数呈显著或极显著负相关。结果表明,低氮下,侧根数目的增加有助于吸收有限的氮素资源,而根系中分配的氮素越多,越有利于促进根系生长,从而造成根冠比增大。
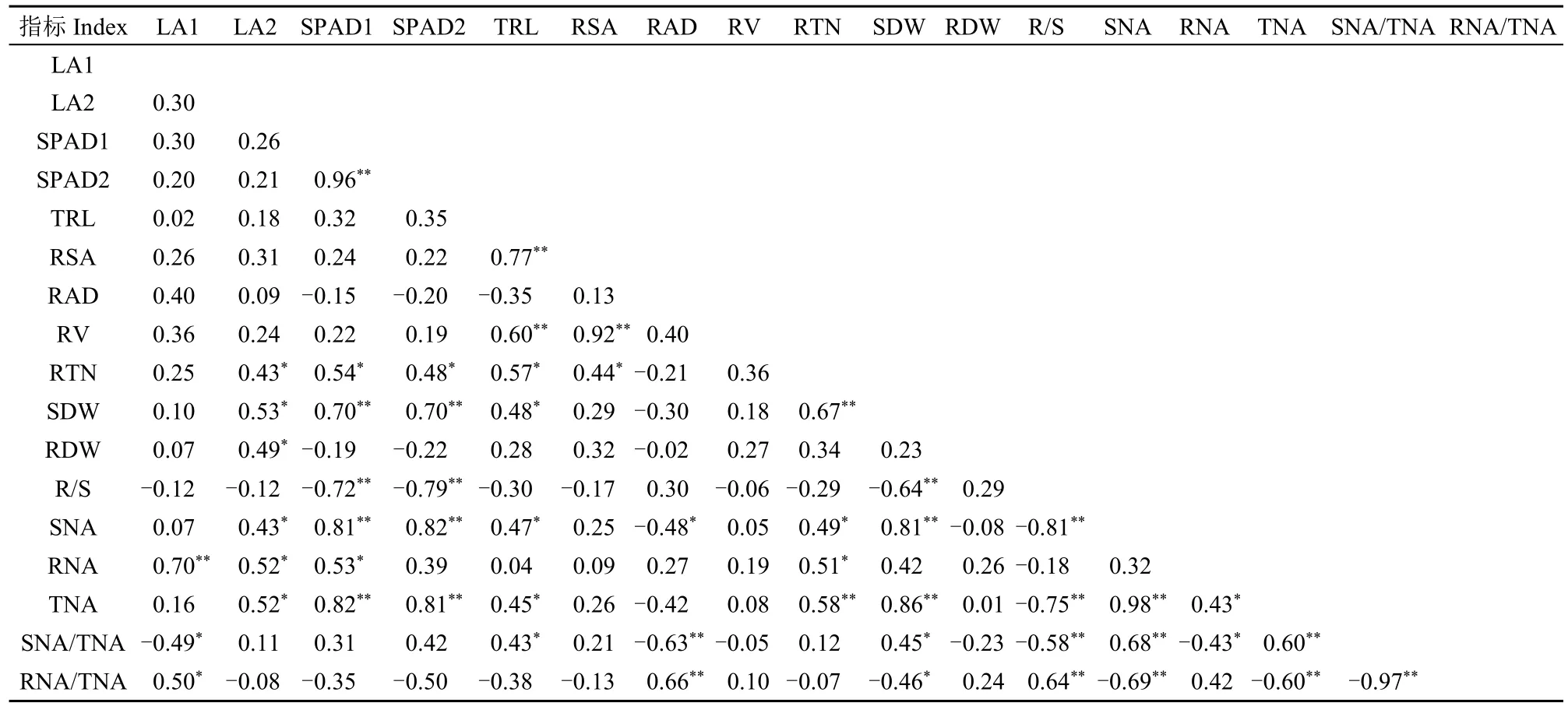
表3 不同性状耐低氮指数的相关性分析Table 3 Correlative analysis of low nitrogen tolerance indexes of the different traits
3 讨论
杂种优势是复杂的数量性状,是整个作物生长周期中各杂种优势性状相互作用的结果[13]。Springer等[12]总结前人研究发现,不同杂交组合同一性状的杂种优势不一致,同一组合不同性状的杂种优势也不相同,在研究的众多成株性状中以产量和穗粒重的超亲优势值较大,均在60%以上。玉米幼苗期对环境非常敏感,是研究其对逆境响应机制的理想时期[29]。研究[9]表明,不同玉米杂交组合发芽后3~7d初生根长度的中亲优势值在17%~25%,而侧根密度的中亲优势值变化范围为51%~130%。水培8d的玉米杂交种幼苗根系在总长度、表面积、体积、根尖数和干重等性状上,中亲和超亲优势均分别在60%和30%以上[26]。本研究中,在相同氮素水平下,豫玉22和先玉335在水培3、7和14d时,均以总根长、根表面积、根体积、根尖数和根干重等根系性状及地上部和根系的氮素积累量的中亲和超亲优势值较大,多数均在50%以上,而叶片面积和叶绿素含量等地上部性状的中亲和超亲优势值较小,多数均在30%以下,甚至有些性状存在负优势,这与前人研究[9,12,26]结论相近。
杂种优势受环境和发育时期双重影响[5-6]。在盐胁迫下,杂交种的膜稳定性和叶绿素含量均高于亲本,而超氧自由基和Na+含量均低于亲本,说明杂交种在耐盐胁迫上存在杂种优势[30]。Betrán等[31]研究指出,低氮会造成玉米产量的杂种优势表现低于正常环境。本研究发现,在低氮胁迫早期(3d和7d),在杂种优势表现上,玉米杂交种先玉335的总根长、根表面积、根平均直径和根体积等性状均高于对照,但其根尖数和根冠比则先后低于对照;而豫玉22仅根干重和根尖数高于对照,但其总根长、根表面积、根平均直径和根体积均低于对照。在低氮胁迫后期(14d),2个杂交种的根冠比、总根长、根表面积和根尖数的中亲优势和超亲优势值均高于对照,先玉335的根干重、根平均直径和根体积的中亲优势和超亲优势值均低于对照,而豫玉22仅根干重的超亲优势值低于对照。由此表明,氮素对玉米幼苗杂种优势的形成有一定影响,且低氮下各个性状的杂种优势表现还会随胁迫时间及基因型的不同而有所改变,低氮会提高一些苗期根系性状的杂种优势,而也有些苗期根系性状的杂种优势形成受低氮抑制。
根系是植物首先感知土壤中水分和矿质元素的器官,对氮素的反应早于地上部[32]。植物的根系对氮素表现出很强的可塑性,可以通过改变根系构型和增大根冠比来适应不同的氮素环境[33]。一般短期缺氮会促进根系生长,而高氮则抑制根系生长[34]。氮素缺乏会促使侧根大量发生,增大根表面积和根体积,从而有利于对水分和营养元素的吸收[35-36]。本研究发现,随着胁迫时间推进,6个基因型玉米根冠比的耐低氮指数从1.02增至2.23,且2个杂交种根冠比的耐低氮指数均与其母本接近,说明低氮导致玉米幼苗根冠比增加,玉米杂交种幼苗根冠比对低氮的响应可能受母体遗传。在根尖数上,在胁迫处理3d和7d时,仅豫玉22及其父本87-1的耐低氮指数在1.0以上,这意味着这2个基因型玉米侧根数目在低氮胁迫下有所增加;另外,仅在胁迫处理14d时,玉米杂交种先玉335的耐低氮指数介于双亲之间,其余2个时间点下均高于任一亲本,而杂交种豫玉22则在任一时间点均高于其双亲,这与表型观察结果基本一致(图1和表3),表明玉米杂交种幼苗侧根数目对低氮的响应可能多受加性或显性效应作用。
氮素调动的变化通常伴随着植株表型性状的改变[19]。研究[25]发现,相比于正常氮素水平,当氮素匮乏时,玉米幼苗根系吸收的氮素在根系中的分配比例大于地上部,甚至氮高效型玉米自交系XY4在根系中的氮素积累量显著高于对照,其根系构型更为庞大,从而保障玉米幼苗的根系生长。在本研究中,在低氮胁迫3、7和14d,玉米杂交种豫玉22的根系氮素积累量占比的耐低氮指数均在1.0左右,这意味着其在2个氮素水平间差异较小,但其2个亲本的根系氮素积累量占比相比于对照均有不同程度降低;随着胁迫时间延长,先玉335及其亲本的根系氮积累量占比相比于对照均不断提高,且在胁迫处理7d和14d时,先玉335的根系氮积累量占比均高于其双亲。由此表明,低氮胁迫下,氮素在玉米幼苗根系中的分配比例会提高,杂交种在根系中分配的氮素要远多于其亲本,从而提高其多个根系性状的杂种优势表现,但这些根系性状对低氮的响应会因遗传背景不同而有较大差异。
4 结论
氮素缺少的早期,玉米幼苗将氮素优先积累在根系中,促进侧根的发生,保证根系生长,根冠比增大,有利于对有限氮素资源的有效吸收。玉米杂交种与其亲本相比,具有更强的氮素平衡能力,多个根系性状存在明显差异,其中侧根数目对低氮的响应可能受加性或显性效应作用,根冠比对低氮的响应可能受母体遗传,但众多根系性状的杂种优势表现对低氮的响应会因遗传背景、生长发育时间的不同而存在较大差异。因此,若想深入研究玉米在氮素利用上的杂种优势机制,还应充分考虑这些影响因素,首先应清楚亲本自交系的遗传背景,然后分析各性状对低氮的响应差异,而在氮高效杂交种选育过程中应注意亲本间各性状的优势互补。
猜你喜欢
西南农业学报(2022年5期)2022-06-06
今日农业(2021年14期)2021-11-25
三农资讯半月报(2020年13期)2020-07-31
科学导报(2018年47期)2018-05-14
农家科技下旬刊(2017年6期)2017-07-05
江苏农业科学(2016年4期)2016-06-14
江苏农业科学(2015年1期)2015-04-17
中学生物学(2009年11期)2009-07-16
发明与创新·大科技(2009年5期)2009-05-31
