
胆汁酸受体在非酒精性脂肪性肝病中的作用
2022-09-07 02:18田盟,严俊,2),3),4),5),李汛,2),3),4),5)*
中国生物化学与分子生物学报 2022年5期
田 盟, 严 俊,2),3),4),5), 李 汛,2),3),4),5)*
(1)兰州大学第一临床医学院, 兰州 730000;2)兰州大学第一医院普外科, 兰州 730000;3)甘肃省生物治疗与再生医学重点实验室,兰州 730000;4)兰州大学医学院肿瘤防治中心, 兰州 730000;5)甘肃省肝胆胰外科研究所, 兰州 730000)
非酒精性脂肪性肝病(non-alcoholic fatty liver disease, NAFLD)是指每天饮酒少于20 g的病人中肝脂肪浸润占全部肝细胞的5%以上,可以表现为单纯性肝脂肪变性、非酒精性脂肪性肝炎(non-alcoholic steatohepatitis, NASH)、肝纤维化和肝硬化,甚至肝细胞癌(hepatocellular carcinoma, HCC)这一系列肝的疾病[1]。目前,全球NAFLD患病率约为25.24%[2]。而在肝细胞癌病人中,NAFLD是Medicare数据库中最常见的潜在病因[3],因此逐渐引起了各国的关注和重视。针对NAFLD的发病机制提出了“两次打击假说”和“多次打击假说”[4,5]。肝内甘油三酯 (triglyceride, TG)蓄积为第一次打击,主要的调节机制有4个:游离脂肪酸 (free fatty acid, FFA)摄取增加、新生脂肪生成增加、脂肪酸氧化 (fatty acid oxidation, FAO)减少和以极低密度脂蛋白形式从肝细胞输出减少,而胰岛素抵抗被认为是脂质内流的主要驱动力,高胰岛素血症和高血糖症也加速脂肪生成和抑制脂肪酸氧化[5];脂质的积累使肝容易受到额外的肝毒性影响(第二次或多次打击),然后导致肝的胆汁酸代谢紊乱、肝的炎症和纤维化[4]。由上述可知,脂质和糖代谢异常以及随后引起的胆汁酸代谢异常、肝的炎症和纤维化都是导致NAFLD的重要因素,但是其上游的具体发病机制仍未完全明确。近些年来,胆汁酸被证明可作为机体信号分子,激活胆汁酸受体来调节代谢和炎症等,本文对这些胆汁酸受体在NAFLD中的作用进行综述。
1 胆汁酸的肠肝循环
在正常情况下,胆汁酸是在肝中合成的,以胆固醇为原料,通过胆固醇7α-羟化酶 (cholesterol 7α-hydroxylase, CYP7A1)的作用产生鹅去氧胆酸(chenodeoxycholic acid, CDCA)和胆酸(cholic acid, CA)。为了增加胆汁酸的溶解度并降低细胞毒性,肝中的初级游离胆汁酸会与甘氨酸或是牛磺酸结合,形成的结合型胆汁酸主要由胆盐输出泵(bile salt export pump, BSEP)从肝分泌到胆管中,经胆囊储存,进餐后胆囊收缩将胆汁酸释放到肠腔。进入肠腔的结合型胆汁酸会在肠道微生物的作用下形成次级胆汁酸(石胆酸(lithocholic acid, LCA)和脱氧胆酸(deoxycholic acid, DCA))。大约95%的胆汁酸在回肠远端被肠上皮细胞重吸收,并通过门静脉转运回肝,其余5%胆汁酸从粪便中排出。上述整个过程被称之为胆汁酸在机体的肠肝循环,每天在人体循环大约6次。这些合成的胆汁酸种类繁多,生理功能各异,不仅在促进肠道脂质以及脂溶性维生素的吸收中发挥作用,而且通过激活胆汁酸受体在介导细胞和分子信号方面也是至关重要。

Fig.1 Pathogenesis of the nonalcoholic fatty liver disease Liver triglyceride (TG) accumulation was the first hit, and there were four main regulatory mechanisms: The intake of FFA increased, the production of new fats increased, the FAO decreased, and the output from liver cells in the form of VLDLs decreased, while insulin resistance is considered to be the main driving force of lipid influx. The accumulation of TG renders the liver susceptible to additional hepatotoxic effects (second or multiple hits), such as oxidative stress, mitochondrial dysfunction and so on, which in turn leads to dysfunction of bile acid (BA) metabolism, liver inflammation and fibrosis. TG: triglyceride; FFA: free fatty acid; FAO: fatty acid oxidation; VLDLs: very low density lipoproteins; NASH: non-alcoholic steatohepatitis; HCC: hepatocellular carcinoma
2 胆汁酸受体与非酒精性脂肪性肝病
NAFLD被认为是代谢综合征在肝中的表现,主要是由于机体代谢紊乱和炎症引起的。胆汁酸受体广泛分布于机体中,包括核受体(法尼醇X受体(farnesoid X receptor, FXR)、孕烷X受体(pregnane X receptor ,PXR)、组成性雄烷受体(constitutive androstane receptor, CAR)和维生素 D 受体(vitamin D receptor, VDR))以及细胞表面受体:跨膜G 蛋白偶联胆汁酸受体(transmembrane G protein-coupled receptor 5, TGR5)、鞘氨醇-1-磷酸受体2(phingosine-1-phosphate receptor 2,S1PR2)和毒蕈碱胆碱受体3(M3 muscarinic receptor, M3R),作为关键代谢过程的转录调节因子被胆汁酸激活后,调节机体的胆汁酸代谢、脂质和糖代谢和能量消耗等。这暗示胆汁酸受体在NAFLD的发生发展中发挥重要作用。
2.1 法尼醇X受体与NAFLD
法尼醇X受体在肝和回肠中广泛表达,也可在胃、心血管、巨噬细胞及白色和棕色脂肪组织中表达,它最有效配体是鹅去氧胆酸,其次是胆酸,脱氧胆酸和石胆酸[6]。FXR的激活除调节胆汁酸代谢和转运外,在调节脂质和葡萄糖代谢、炎症反应和纤维化等方面亦发挥关键作用,影响NAFLD发展的各个阶段。因此,调控FXR及相关因子的表达,是NAFLD治疗药物研究的潜在靶点。
2.1.1 FXR调节胆汁酸代谢 胆汁酸代谢包括胆汁酸合成以及胆汁酸转运。FXR通过抑制胆汁酸合成和促进胆汁酸排出来减少胆汁酸在肝内淤积而引起的肝毒性作用,例如肝纤维化、肝硬化和肝癌等。因此,FXR对胆汁酸代谢的调节可减少NAFLD的发生。FXR可以通过检测肝和肠道中胆汁酸浓度的升高来负反馈抑制肝中CYP7A1基因的表达,减少胆汁酸合成。在肝中,激活的FXR诱导小分子异源二聚体伴侣受体(small heterodimer partner, SHP)表达。SHP会干扰其他核受体,包括肝受体同源物1(liver receptor homologue1, LRH-1)和肝X受体(liver X receptors, LXR),从而抑制CYP7A1基因的表达[5];而在回肠末端,激活的FXR会诱导人类肠道成纤维细胞生长因子 19 (fibroblast growth factor-19, FGF19)(人FGF19,小鼠FGF15)的表达,使FGF15/19通过门静脉血到达肝内,与肝细胞表面FGFR4/β-klotho异二聚体复合物结合,触发JNK1/2和ERK1/2信号级联反应,也抑制了CYP7A1的表达[2]。除了影响胆汁酸的合成,FXR还可调节胆汁酸转运体,激活FXR可导致胆汁酸向小肠的转运增加,从血液中的摄取减少,促进胆汁酸随粪便和尿液排出体外,防止NAFLD的发生。在回肠中,当胆汁酸浓度增加时,会通过位于回肠末端的钠依赖性胆汁酸转运蛋白(apical sodium-dependent bile acid transporter, ASBT)转运到回肠上皮细胞,从而激活肠道FXR,继而在回肠胆汁酸载体蛋白质和肠OST-α/β的帮助下转运入门静脉[7],抑制位于肝细胞上的钠依赖性牛磺胆酸盐共转运多肽(normal tissue complication probability, NTCP),它介导了门脉循环中大多数胆汁酸的吸收,并使肝细胞上的BSEP上调,胆汁酸排出增加[8]。由此可见,来自肝和肠道的胆汁酸信号可通过负反馈对自身进行调节,激活FXR下调胆汁酸浓度,防止肝内胆汁淤积,在保护肝免受毒性损害方面发挥重要作用。
2.1.2 FXR促进脂质和糖代谢 肝是脂质储存和处理的重要场所。脂质在肝中的蓄积是造成NAFLD的第一次打击,FXR可以通过抑制肝对脂肪酸的摄取、抑制肝内新生脂肪的生成、促进FAO和促进脂质的排出来调节脂质代谢。肝对脂肪酸 (fatty acids,FAs)的摄取主要依赖于位于质膜上的特定脂肪酸转运体,例如CD36[9]。在NAFLD患者中,CD36的表达与肝中的TG浓度呈正相关。激活FXR可导致 CD36 的表达下降[10]。新生脂肪的生成受多种转录因子调控。其中,甾醇调节元件结合蛋白1c (sterol regulatory element-binding protein-1c, SREBP-1c),以及糖类反应元件结合蛋白(carbohydrate-responsive element binding protein, ChREBP)是主要调节因子。FXR通过诱导SHP的表达抑制肝X受体和肝X受体诱导的SREBP-1c的激活,进而抑制了造脂酶(lipogenic enzymes)的表达[11];另外,FXR通过促进ChREBP的释放,阻碍维甲酸和甲状腺激素受体募集到L型丙酮酸激酶 (L-type pyruvate kinase, LPK)启动子的核心部位,来抑制葡萄糖诱导的LPK的表达,从而减少新生脂肪生成所需的乙酰辅酶A的产生,减轻肝的脂肪变性[12]。这提示,FXR可以通过抑制脂肪酸的摄取和新生脂肪的生成,从源头上减少肝内脂肪的合成量,实现了“节源”。对于肝中已经存在的FAs,FXR可以通过多种途径使其氧化和排出。野生型C57BL/6J小鼠经CA处理后,FAO基因表达增加,可能是由于FXR通过诱导过氧化物酶体增殖物激活受体α (peroxisome proliferator-activated receptor alpha, PPARα)的表达来实现的[13];除此之外,FXR还能直接或是通过FGF19增加调节脂肪酸氧化的重要细胞因子FGF21的表达和分泌[5]。而血清TG浓度的降低部分是由于FXR介导脂蛋白脂酶(lipoprotein lipase, LPL)调节因子的转录,LPL可以水解极低密度脂蛋白颗粒和乳糜粒中TG[5]。载脂蛋白C-II (apolipoprotein CII , ApoC-II)是LPL的激活因子,载脂蛋白C-III (apolipoprotein CIII, ApoC-III)和血管生成素样蛋白 3(angiopoietin-like protein 3, Angptl3)是脂蛋白脂酶抑制剂,FXR通过诱导ApoC-II的表达,抑制ApoC-III和Angptl3的表达,促进脂蛋白脂酶的过表达,从而促进TG的清除。上述表明,FXR不仅可以做到对肝内脂质的“节源”,还能做到“开流”。因此,有效地激活FXR可以被开发用来治疗脂肪肝疾病。进一步深入研究发现,FXR还会在基因水平上调节糖脂代谢。肥胖患者中包括 miR-802在内的microRNA表达异常升高,FXR可诱导SHP的表达级联调节miR-802的表达,以维持葡萄糖和脂质稳态[14]。由此可见,FXR影响脂质代谢的全过程,从基因水平-摄取-合成-氧化-排出,对FXR的控制可有效地防止肝内脂质蓄积,限制NAFLD的发生。
胰岛素抵抗或是糖异生造成的高血糖会促进脂肪酸内流,加速脂质的形成,导致肝脂肪变性。FXR通过维持葡萄糖稳态而抑制了NAFLD的发生。磷酸烯醇式丙酮酸羧激酶(phosphoenolpyruvate carboxykinase, PEPCK)和葡萄糖-6-磷酸酶(glucose-6-phosphatase, G-6-Pase)是糖异生的相关基因,FXR激活SHP可通过抑制他们来调节糖异生[15];而FXR激活的FGF15/19似乎也在葡萄糖调节中起关键作用。在餐后状态下,FGF15/19从小肠释放出来,通过cAMP调节元件结合蛋白的去磷酸化和失活来抑制肝脏糖原的分解[16]。FXR抑制糖异生的作用为NAFLD的预防和治疗提供了新见解。
2.1.3 FXR调节肝的炎症、纤维化 肝的炎症、纤维化是NAFLD发展的严重阶段,FXR被证明直接抑制脂多糖(lipopolysaccharide, LPS)诱导的促炎细胞因子和巨噬细胞趋化因子的产生,包括TNFα、IL-1β、IL-6和MCP-1[17]。鉴于其在疾病发病机制中的重要性,这种减少很可能改善肝的炎症;除了抑制肝炎症的发生,FXR还通过诱导肝星状细胞(hepatic stellate cells, HSCs)表达抗纤维化基因而直接抑制肝纤维化,或通过诱导肝星状细胞PPARγ的表达使HSC失活[18]。FXR肝内的高表达具有直接的抗炎和抗纤维化作用,防止其进一步恶化为肝细胞癌。
2.2 孕烷X受体、组成性雄烷受体、维生素 D 受体与NAFLD
PXR和CAR主要在肝和肠道中表达,由石胆酸激活[19]。PXR和CAR同样可以调节机体代谢和炎症反应。但 PXR在NAFLD的发生发展中扮演的角色尚无定论,起着双重作用。首先,PXR和CAR除了通过抑制胆汁酸合成和促进胆汁酸排出减少其肝毒性外,还直接具有解毒作用。其中,PXR能够促进FGF15/19分泌进而抑制CYP7A1,减少胆汁酸的合成[20];而PXR和CAR都可诱导细胞色素P450 3A4 (cytochrome P450 3A4, CYP3A4)和细胞色素P450 2B6 (cytochrome P450 2B6, CYP2B6)介导的羟基化作用,使胆汁酸解毒,或诱导胆汁酸结合酶的表达来保护肝免受毒性。还能诱导多药耐药相关蛋白 3 (multidrug resistance-associated protein 3, MRP3)和多药耐药相关蛋白 4 (multidrug resistance-associated protein 4, MRP4)表达,促进胆汁酸排入体循环,再经肾由尿液排出[21]。除调节胆汁酸代谢以外,PXR和CAR还可调节糖和脂质代谢,但PXR缺失通过抑制肝脂肪生成、增加耗氧量和线粒体β氧化,以及增加胰岛素的敏感性来减少肝脂肪变性和胰岛素抵抗[22]。故激活PXR反而会促进肝的脂肪变性,对肝发挥损害作用。而CAR恰恰相反,被证明作用于小鼠Pnpla3启动子间接调控肝新生脂肪形成,同时结合并抑制叉头盒蛋白O1(forkhead box protein O1, FOXO1),阻止PEPCK1和G-6-Pase的表达,从而影响胰岛素受体信号转导,阻止糖异生[20,23];抑制肝糖异生也可以归因于CAR促进泛素化和过氧化物酶体增殖物激活受体γ共激活因子 1 α(peroxisome proliferator-activated receptor γ coactivator 1α, PGC-1α)的降解[24]。另外,PXR和CAR都具有抗炎的特性。PXR和CAR能阻断许多促炎的NF-κB靶基因的产生,PXR还会上调分泌型白细胞介素1受体拮抗剂的产生。这通过减少IL-6和TNFα的产生,以及阻断IL-1β在细胞表面的作用来抑制LPS诱导的炎症[25,26]。因此,CAR主要能防止肝脂肪变性、NASH和肝纤维化等。而PXR因为其促进肝脂肪变性的特性对机体调节是复杂的,对肝总体是发挥保护还是破坏作用,仍需要根据具体情况具体分析。
VDR在肠道、成骨细胞、肾和多种类型的免疫细胞中高度表达,在肝中低水平表达,可被LCA激活[27]。通常VDR被激活后,也能够诱导CYP3A4和胆汁酸结合酶的表达,抑制CYP7A1的表达[28]。但最新的研究发现,脂肪生成基因Angptl3和血管生成素样 8 (angiopoietin-like 8,Angptl8)是人肝细胞中可被胆汁酸诱导的新型VDR靶基因,NAFLD患者肝内VDR表达和功能增加,同时ANGPTL3和ANGPTL8基因上调,有助于TG的积累[27,29]。这项研究为肥胖中ANGPTLs介导的异位脂肪蓄积和NAFLD发育的潜在机制提供了新颖的见解。以上研究表明,胆汁酸核受体的功能是非常复杂的,正确且全面地认识PXR、CAR和VDR的分子功能,对于进一步研究NAFLD新的治疗靶点具有长远意义。
2.3 G蛋白偶联胆汁酸受体与NAFLD
TGR5几乎在人体和动物所有的组织中均表达,以胆囊表达最高,其次是回肠和结肠,肝 (Kupffer细胞,不是肝细胞)、棕色脂肪组织、肌肉和中枢神经系统中的表达丰度较低。它主要被石胆酸激活,其次是DCA、CDCA和CA[30]。TGR5主要通过增加 cAMP 的细胞内浓度来传递其信号,导致下游激酶的快速磷酸化,涉及抗炎症反应和能量代谢等方面,也可直接调节胆汁酸和糖脂代谢,进而防止肝的脂肪变性等NAFLD的发生发展。
2.3.1 TGR5调节胆汁酸代谢 对于胆汁酸的调节,TGR5可以通过抑制胆酸合成和改变胆汁酸成分来实现。Liu等[31]研究发现,TGR5抑制小鼠肝细胞中CYP7A1的表达,胆汁酸池明显减少;TGR5还调节胆汁成分,当喂食致石饮食时,TGR(-/-)小鼠可以免受胆结石疾病的影响,这可能是因为TGR5激活可诱导胆管细胞分泌碳酸氢盐和氯化物改变胆汁酸成分[5]。但具体改变成分仍不太清楚。激活TGR5可抑制胆汁酸合成,但是作为调节机体代谢的重要胆汁酸受体,其在胆汁酸转运方面的作用仍不明确,需要进行深入研究。
2.3.2 TGR5调节脂质和糖代谢 激活肠内分泌L细胞上的TGR5可调节线粒体氧化磷酸化,导致的ATP / ADP比值的增加以诱导钙动员以及膜的去极化,诱导胰高血糖素样肽-1(glucagon-like peptide 1, GLP-1)释放,这不仅可以抑制肝脂肪生成,与胰腺中高度表达的GLP-1受体结合,还可调节葡萄糖代谢,促进胰岛新生和存活[32],增强胰岛B细胞中胰岛素分泌,改善胰岛素抵抗,并由于其抑制胰岛B细胞中胰高血糖素的分泌而减少了肝的内源性葡萄糖的产生[33]。这也被认为有助于其在改善胰岛B细胞功能和血糖控制方面的作用。同时,有研究[34]发现,FXR其实是和TGR5在L细胞中共定位,并且FXR可诱导TGR5刺激GLP-1分泌。这表明就抑制肝脂肪变性而言,FXR和TGR5之间存在联合作用或协同作用机制。
2.3.3 TGR5调节能量代谢 正常情况下,脂肪占人体供能的20%~25%。在棕色脂肪和骨骼肌中,胆汁酸和TGR5之间的相互作用增加了细胞内cAMP。刺激cAMP依赖的蛋白激酶A(protein kinase A, PKA)激活ChREBP,诱导甲状腺激素脱碘酶2(thyroid hormone deiodinase 2, D2)的表达,将失活的甲状腺素(thyroxine4, T4)转化为活性的3,5,3-三碘甲状腺原氨酸(thyroxine3, T3),增加棕色脂肪组织和肌肉中能量的消耗和线粒体的氧化磷酸化[35]。因此,和其他胆汁酸受体不同,TGR5还能增加肥胖小鼠的能量消耗,防止高脂血症诱导的肥胖。
2.3.4 TGR5抑制肝的炎症 TGR5在巨噬细胞中高表达,胆汁酸可调节这些细胞的炎症反应。TGR5增加了大鼠肺泡巨噬细胞内cAMP的表达,从而降低了巨噬细胞的吞噬活性,抑制了LPS诱导的促炎细胞因子的产生[5]。另外,TGR5激活可抑制内毒素激活的Toll样受体4(toll-like receptor 4, TLR4)引起的炎症介质的表达,但在TGR5-/-小鼠肝中无此作用[36],可以确定TGR5是肝炎症的负调节因子。
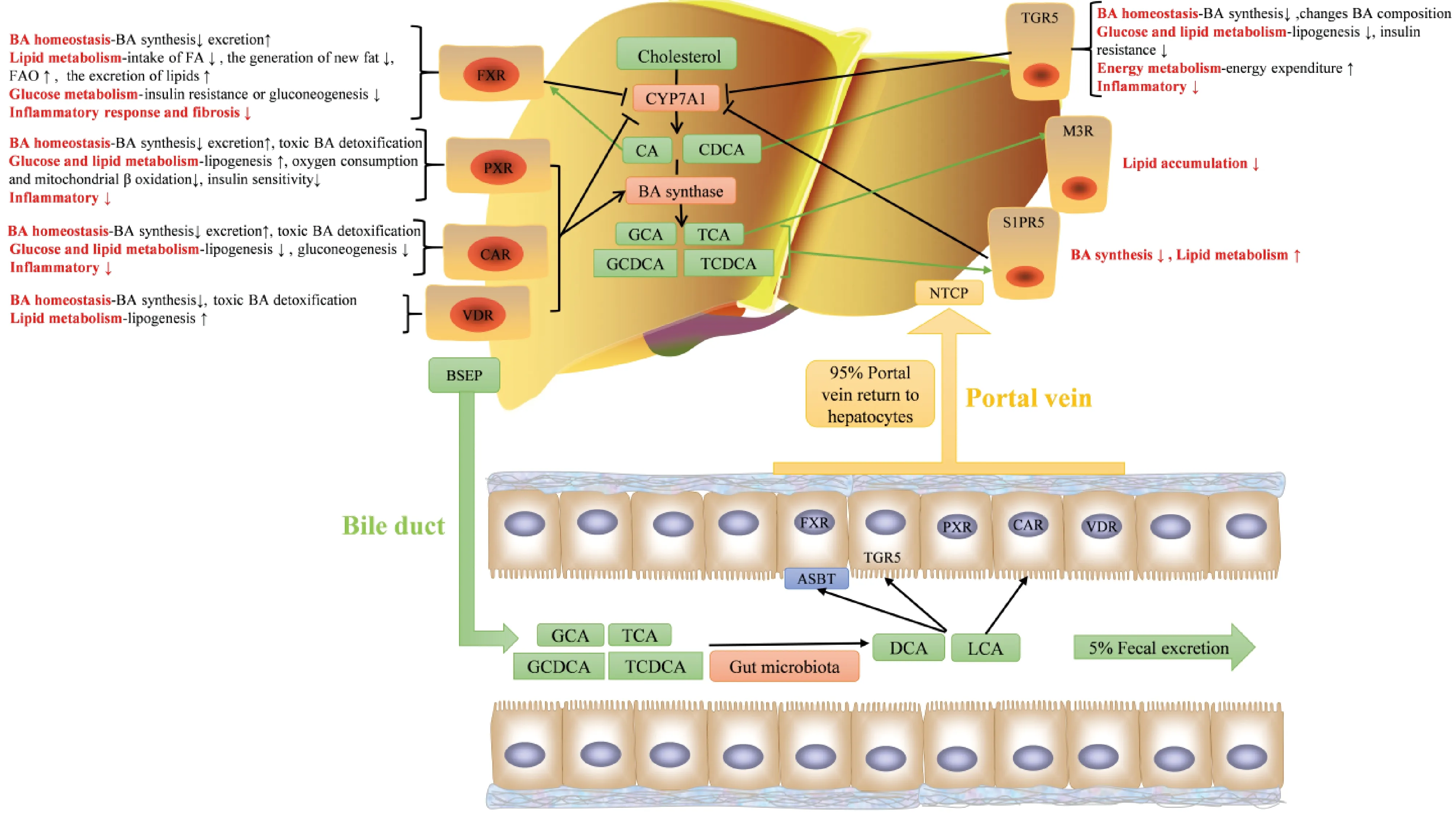
Fig.2 Enterohepatic circulation of bile acid (BA) and Schematic representation of the action of BA receptors in several tissues Bile acids are synthesized in the liver and enter the intestine through the bile ducts, and 5% are excreted in the faeces and 95% are reabsorbed into the blood. The nuclear receptors, FXR, PXR, CAR, VDR, are mainly found in the liver and intestines, while cell surface receptors, TGR5, are mainly found in the liver and intestines. S1PR2 and M3R in the intestines, have several downstream effects, including BA homeostasis, glucose and lipid metabolism, energy metabolism, inflammatory response and fibrosis. FXR: farnesoid X receptor; PXR: pregnane X receptor; TGR5:transmembrane G protein-coupled receptor 5; S1PR2:phingosine-1-phosphate receptor 2; M3R: M3 muscarinic receptor; BSEP: bile salt export pump; NTCP : normal tissue complication probability; ASBT: the apical sodium-dependent bile acid transporter; CA: cholic acid, CDCA: chenodeoxycholic acid, DCA: deoxycholic acid, LCA: lithocholic acid
2.4 鞘氨醇-1-磷酸受体2、毒蕈碱胆碱受体3与NAFLD
S1PR2分布广泛,高表达于肝、心、肾、脾、肺、脑和血管平滑肌细胞,结合型胆汁酸是它的有效配体[28]。S1PR2除了调节胆汁酸合成外,还能促进脂质代谢,在肝的病理生理中发挥独特而关键的作用。有研究[37]表明,当血清中结合型胆汁酸的浓度高达156 μmol/L,可以激活S1PR2,从而导致Jnk1/2激活,SHP上调和CYP7A1抑制。在原代啮齿动物肝细胞中,S1PR2可激活下游ERK1 / 2和AKT信号通路,它们在肝糖和脂质代谢的调节中发挥重要作用;并且最近Nagahashi等[38]研究表明,在原代小鼠肝细胞中,编码S1PR2基因的过表达增加了SphK2,S1PR2-/-和SphK2-/-小鼠均在高脂饮食下迅速发育出脂肪肝。上述结果表明,结合型胆汁酸-S1PR2途径在调节肝脂质代谢中的重要性。
M3R是毒蕈碱受体存在于肝细胞中的主要亚型,可被TCA激活[39]。有研究[40]表明,M3R通过激活AMPK通路,对肝细胞脂质堆积具有保护作用。因此,肝细胞中的胆碱能信号在维持肝的正常代谢中发挥重要作用,M3R是防止肝脂肪变性的潜在治疗靶点。
3 胆汁酸受体作为非酒精性脂肪性肝病治疗靶点
鉴于胆汁酸受体的上述生物学效应,胆汁酸受体成为治疗NAFLD的潜在目标。其中,以FXR和TGR5为主。奥贝胆酸(obeticholic acid,OCA)是强效的FXR激动剂,可以通过激活 FXR降低肝的脂肪生成和增强VLDLs在外周组织的清除,以及增加胰岛素敏感性并减少肝内糖异生。在FLINT研究[41]中,分析了NAFLD病人用OCA组与安慰剂组相比,每d口服25 mg的OCA持续72周,可改善组织学NASH评分至少降低2分,而无纤维化恶化。但是,已观察到OCA会增加血浆脂质浓度并引起瘙痒,因此,必须要通过进一步研究进行评估。其他FXR激动剂,例如GW4064和WAY-362450也可以通过激活FXR参与肝脂肪形成的过程,从而减轻了肝的脂肪变性[42]。此外,TGR5激动剂INT-777可改善高脂饮食诱发的肥胖小鼠的肝脂肪变性和肥胖症,并改善其胰岛素敏感性[43]。使用FXR/TGR5双重激动剂INT-767的肥胖小鼠减少了肝脂肪变性,减少了促炎细胞因子的表达[44]。因此,除了FXR激动剂以外,TGR5 激动剂对于治疗NAFLD也具有非常大的潜力,但目前尚缺乏TGR5激动剂治疗NALFD的相关临床研究,能否成为NAFLD治疗靶点仍需要更多的研究支持。
4 问题与展望
综上所述,胆汁酸激活的胆汁酸受体在胆汁酸代谢、脂质和糖代谢、能量代谢、肝的炎症和纤维化等方面具有重要作用,进而影响NAFLD的发生与发展,进一步完善了NAFLD的发病机制。胆汁酸受体数量众多且作用广泛,虽然大多数胆汁酸受体对于NAFLD的发展是发挥保护作用,但随着对其研究的深入,也会发现例如PXR、VDR等防止肝内胆汁淤积的同时会促进肝内脂肪蓄积。对于NAFLD起着双重作用,有待对其目前已知的上、下游信号通路进行更加深入的研究和其他代谢通路的探索,以更加全面地了解他们的作用。另外,不同的胆汁酸受体之间可能存在交互作用,例如FXR可以和TGR5协同调节糖脂代谢。因此,不同胆汁酸受体的联合研究非常有价值。并且,目前应用于临床的多为单一胆汁酸受体激动剂。探寻例如INT-767的双重或是多重受体激动剂,能够更好地治疗NAFLD,更好地投入临床。最后,我们认为,胆汁酸受体具有极好的研究和运用前景,未来可逐步发掘出其在NAFLD预防、诊断和靶向治疗等各个环节的临床运用价值。
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中国药学药品知识仓库(2022年9期)2022-05-23
中国典型病例大全(2022年13期)2022-05-10
家庭医学·下半月(2021年1期)2021-03-28
家庭科学·新健康(2020年6期)2020-07-06
科学导报(2019年45期)2019-09-23
保健与生活(2019年23期)2019-09-10
家庭医学(2018年2期)2018-07-14
分析化学(2018年1期)2018-01-18
黄河黄土黄种人(2017年11期)2017-11-27
