
基于渔业数据的南极磷虾48渔区渔场时空分布*
2022-09-05 02:00赵国庆罗俊荣唐峰华宋学锋
渔业科学进展 2022年4期
赵国庆 罗俊荣 唐峰华 樊 伟 宋学锋 杨 超 张 衡
基于渔业数据的南极磷虾48渔区渔场时空分布*
赵国庆1,2罗俊荣3唐峰华2樊 伟2宋学锋2杨 超1,2张 衡2①
(1. 上海海洋大学海洋科学学院 上海 201306;2. 中国水产科学研究院东海水产研究所 上海 200090;3. 福建正冠渔业有限公司 福建 福州 350500)
根据我国2010—2019年南极磷虾()捕捞渔船的生产资料,分析了南极海域48渔区南极磷虾渔场的分布特点,采用重心迁移轨迹模型和标准差椭圆(SDE)模型探讨了南极磷虾的渔场变动特征和规律。结果显示,南极磷虾捕捞量主要集中在48.1亚区,占比为70.30%,48.2亚区和48.3亚区的产量相差很小,占比分别为14.28%和15.42%;年间单位捕捞努力量渔获量(CPUE)曲线上升,最小值为2012年,最大值为2019年;月间CPUE先增后降,最小值为1月,最大值为6月。48.1亚区的年间和月间渔场重心均往西南方向移动;48.2亚区年间的渔场重心东移,但移动范围较小,月间规律不强;48.3亚区年间渔场重心南移,月间渔场重心向西北移动。经SDE分析可知,48.1亚区渔场分布范围最广、离散程度最大,48.3亚区渔场方向性最强、向心力最明显。48.1亚区渔场重心主要分布于布兰斯菲尔德海峡,48.2亚区渔场重心分布于南奥克尼群岛东侧,48.3亚区渔场重心分布于南乔治亚群岛东北侧。聚类结果表明,48.1亚区年间渔场重心均较为集中,48.2和48.3亚区除2017年外,其他年间渔场重心较为集中。
南极磷虾;渔场重心;标准差椭圆分析;聚类分析;时空分布;48渔区
南极磷虾通常指南极大磷虾(,以下称磷虾),属于甲壳类浮游动物,广泛分布于南极大陆周围,既是浮游植物的主要捕食者,又是鱼类、头足类、企鹅、海豹、鲸鱼等的主要饵料,在食物网中起着承上启下的作用,是整个南大洋生态系统物质和能量流动的关键物种(Siegel, 2005; Atkinson, 2012; Stowasser, 2012)。在地球已知的多细胞动物中,磷虾生物量最高,据估算,其资源量可达3.0×108~5.0×108t (Atkinson, 2009)。磷虾脂类物质含量很高,富含长链多不饱和脂肪酸(Ericson, 2018),被称为全球最大的动物蛋白质库,另外,还富含丰富的矿物元素和胡萝卜素等(Wang, 2011)。因此,磷虾具有十分巨大的开发价值。磷虾捕捞业始于20世纪60年代早期,中国于1984年首次开展南极科学考察时即将磷虾列为重点考察目标,而正式商业性探捕开发始于2009—2010年度(国家海洋局极地专项办公室, 2016)。目前,我国磷虾捕捞技术和捕捞产量都取得了重大进展,由2009—2010年度4000 t上升到2019年50 000多t (CCAMLR, 2020)。
掌握磷虾渔场时空分布对磷虾商业性捕捞具有重大意义。目前,对磷虾分布特征的研究,主要通过大型渔船的生产资料(Krafft, 2015)、科学调查资料(Marrari, 2008)、暂养实验(Kawaguchi, 2010a)或远程监测(Saenz, 2020)等进行,相关研究多结合环境数据(Atkinson, 2019)或借助声学探测(Seok, 2018; 张吉昌等, 2012)来进行,侧重于某一时空尺度下磷虾的分布特征或已知因子如何影响磷虾分布,并探讨磷虾分布是否存在统计意义上的聚集或离散性质。Krafft等(2015)通过商业捕捞渔船对磷虾丰度和垂直结构等种群动态进行研究后认为,使用商业捕捞渔船作为研究平台,对了解渔业动态及量化生物资源量水平具有很大的优势。Patricio等(2020)通过2011—2016年磷虾捕捞数据总结了智利磷虾渔业现状,展示了磷虾捕捞深度、单位捕捞努力量渔获量(CPUE)、网次地理分布等。张瑛瑛等(2020)利用2010—2015年1—6月的磷虾捕捞数据研究了渔场重心总体上的轨迹变化,并探讨了CPUE与海表温度和叶绿素(Chl-)的关系。但是,目前基于长时间序列的磷虾渔业数据对磷虾渔场的变动时间、变动方向和变动格局的研究鲜有报道。因此,本研究依据我国2010—2019年磷虾捕捞渔船的生产资料,探讨了48区磷虾渔场重心轨迹的迁移和分布格局情况,以期为磷虾合理开发利用提供借鉴。
1 材料与方法
1.1 数据来源及捕捞海域
渔业数据来源于我国磷虾捕捞渔船(中国水产总公司、上海开创远洋渔业公司、辽宁远洋渔业公司等10艘渔船)的渔捞记录资料(图1),包括作业日期(年、月、日)、作业位置(经度和纬度)、渔获产量、拖网次数、拖网时间、拖速、网具类型、拖网作业时的网口高度和网口水平扩张。时间跨度为2010—2019年,主要作业月份为每年的1—9月。作业区域主要分布在南极海域48.1区、48.2区和48.3区,空间分辨率为1°×1°(图2)。
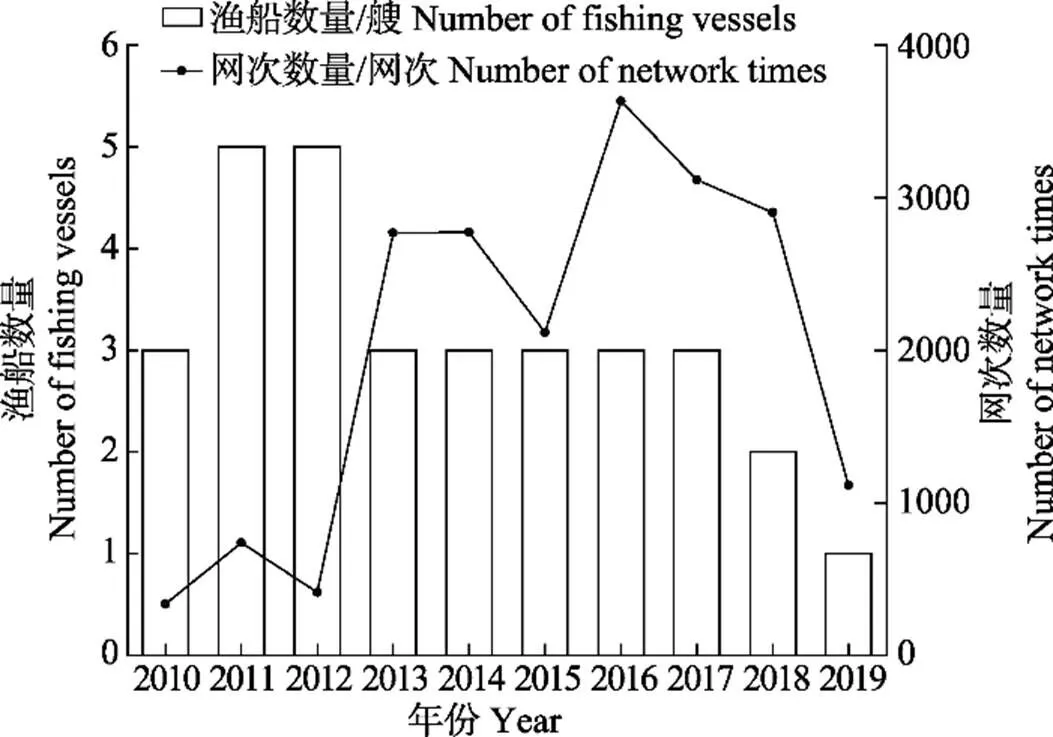
图1 南极磷虾拖网渔船数量和拖网次数统计
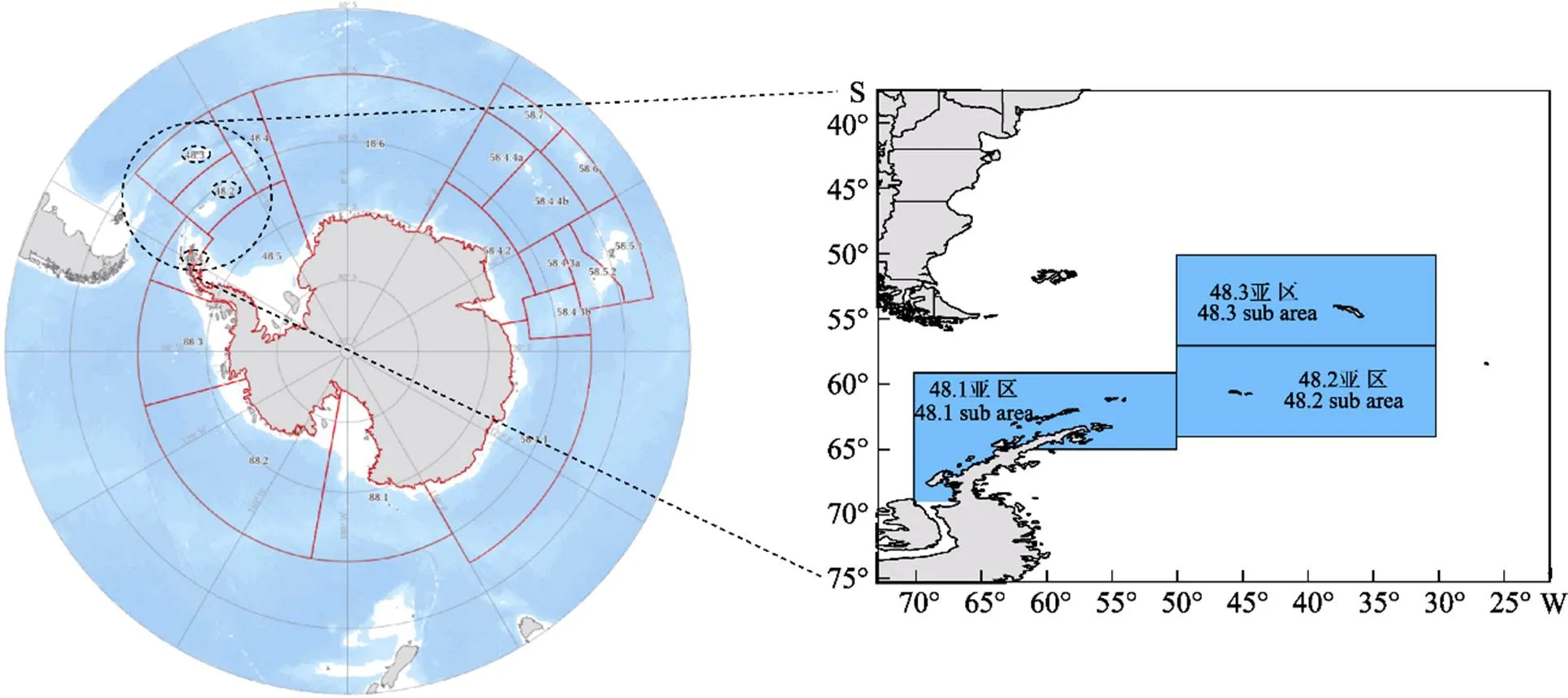
图2 调查海域示意图
1.2 数据处理
1.2.1 渔获数据处理 对渔获数据进行标准化处理,采用标准化后的CPUE来表示磷虾渔场资源丰度,单位为t/h,计算公式为:
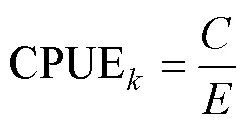
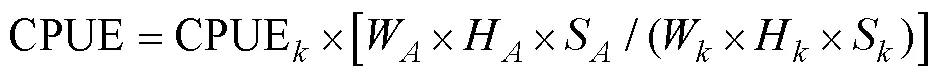
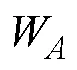
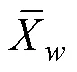
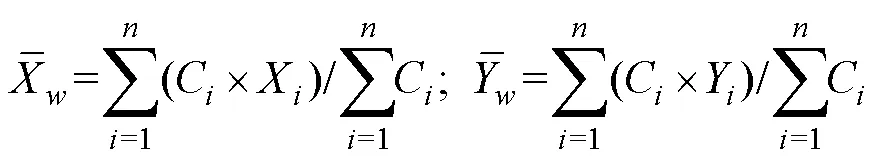
1.2.3 标准差椭圆分析 标准差椭圆(standard deviational ellipse, SDE)是一种定量描述研究对象空间分布整体特征及时空演变过程的空间格局统计分析方法,能够精确表达地理要素空间分布的整体特征(Lefever, 1926),本研究采用产量作为权重字段,用其来揭示磷虾渔场的变动方向和分布格局。SDE相关参数计算公式如下:
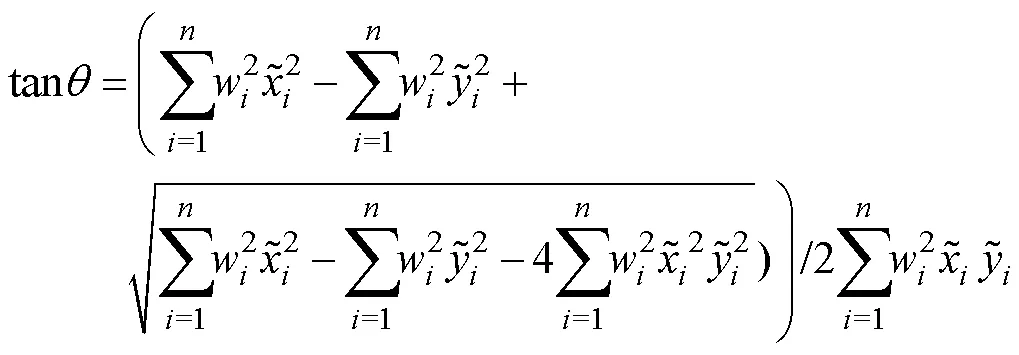
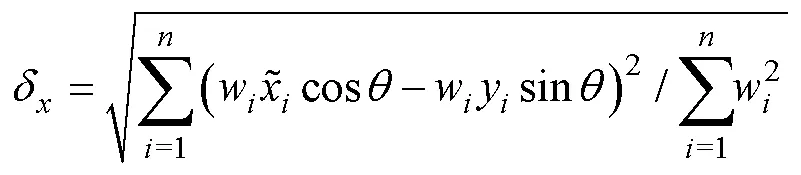
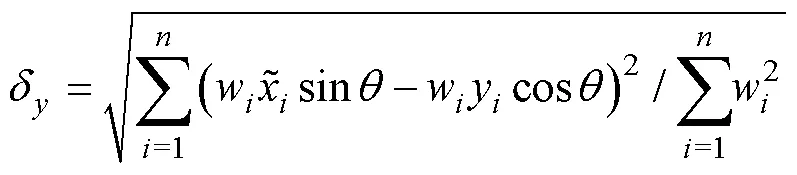
1.2.4 渔场重心聚类分析 利用欧式距离来计算年产量重心的距离,比较渔场重心的变化情况,公式如下(Carlo, 2021):
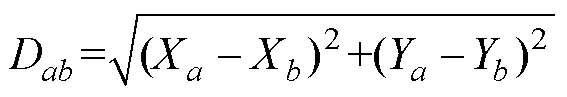
式中,D为年和年产量重心之间的距离,、分别为第年的经度和纬度,X、Y分别为第年的经度和纬度。根据欧式距离,对2010—2019年各年的产量重心按照最小距离法进行聚类,分析比较年份和月份之间的差异性。
2 结果与分析
2.1 磷虾产量和CPUE时空变化
将捕捞区域分为1°×1°若干单位小渔区,统计2010—2019年每单位渔区的磷虾总产量,产量分布见图3A,CPUE分布见图3B。2010—2019年的产量集中于48.1亚区,48.2、48.3亚区的产量占比和有产量的单位小渔区分布数量相差较小,且产量多集中在各亚区的岛屿周围。48.1、48.2和48.3亚区的产量分别占总产量的70.30%、14.28%和15.42%。各亚区CPUE的变化与产量的变化一致,各区域中平均CPUE值以48.1亚区最高,其次为48.2亚区,48.3亚区最低,分别为19.95、12.31和11.69 t/h。
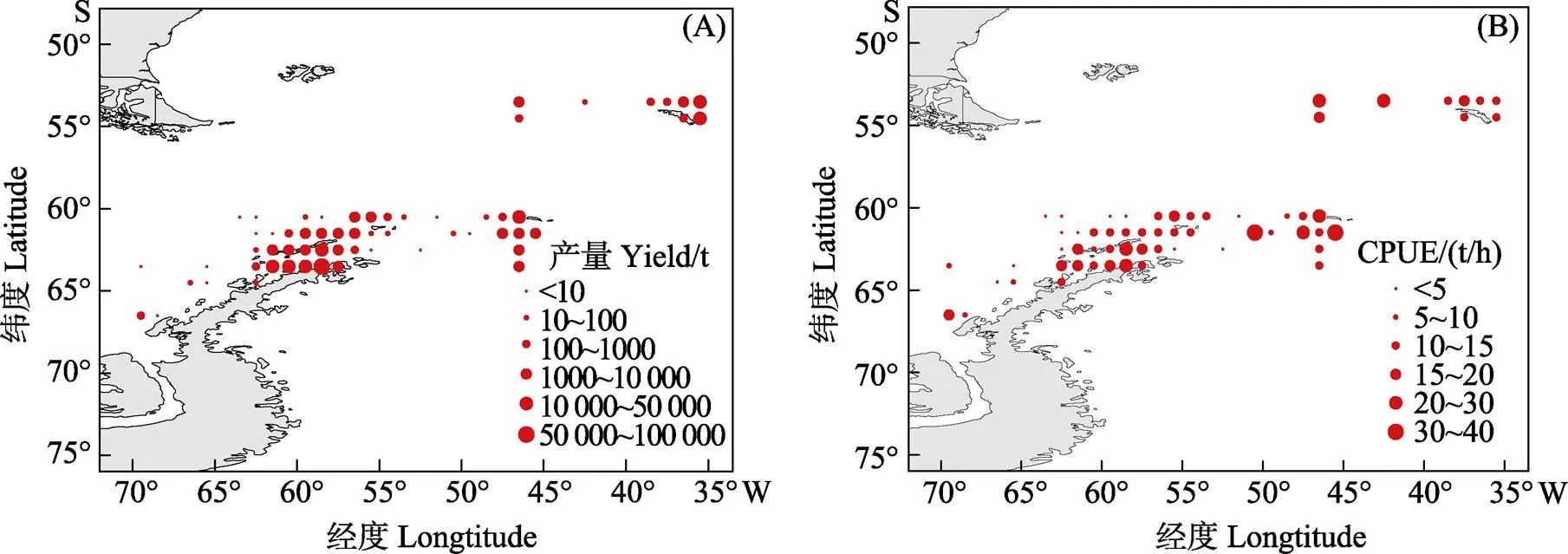
图3 2010—2019年磷虾产量空间变化(A)和CPUE空间变化(B)
年间捕捞力量的投入存在较大差异,年间产量差距较大,因此,本研究仅对CPUE进行统计分析。从年间变化来看,CPUE呈上升趋势,在2019年达到最大值,最小值发生在2012年。网次占比的变化情况基本与CPUE的变动趋势相似,最小值和最大值分别发生在2010年和2016年(图4A)。从月间变化来看,1—9月CPUE和网次占比均呈先增后降的趋势,且均在9月达到最低值(图4B)。
2.2 磷虾产量重心的时空变化
从年间变化来看,48.1亚区的产量重心呈现西南–东南–西南的变动趋势,产量重心总体上向西南方向移动(图5A);48.2亚区的产量重心在2010—2016年基本上在东西方向上来回移动,在2017年往南移动,2018年和2019年又基本移动到2013年重心的经度上,但在纬度上东移,产量重心总体向东移动(图5B);48.3亚区的产量重心除2017年外总体上在南北方向上移动,且整体上往南移动(图5C)。
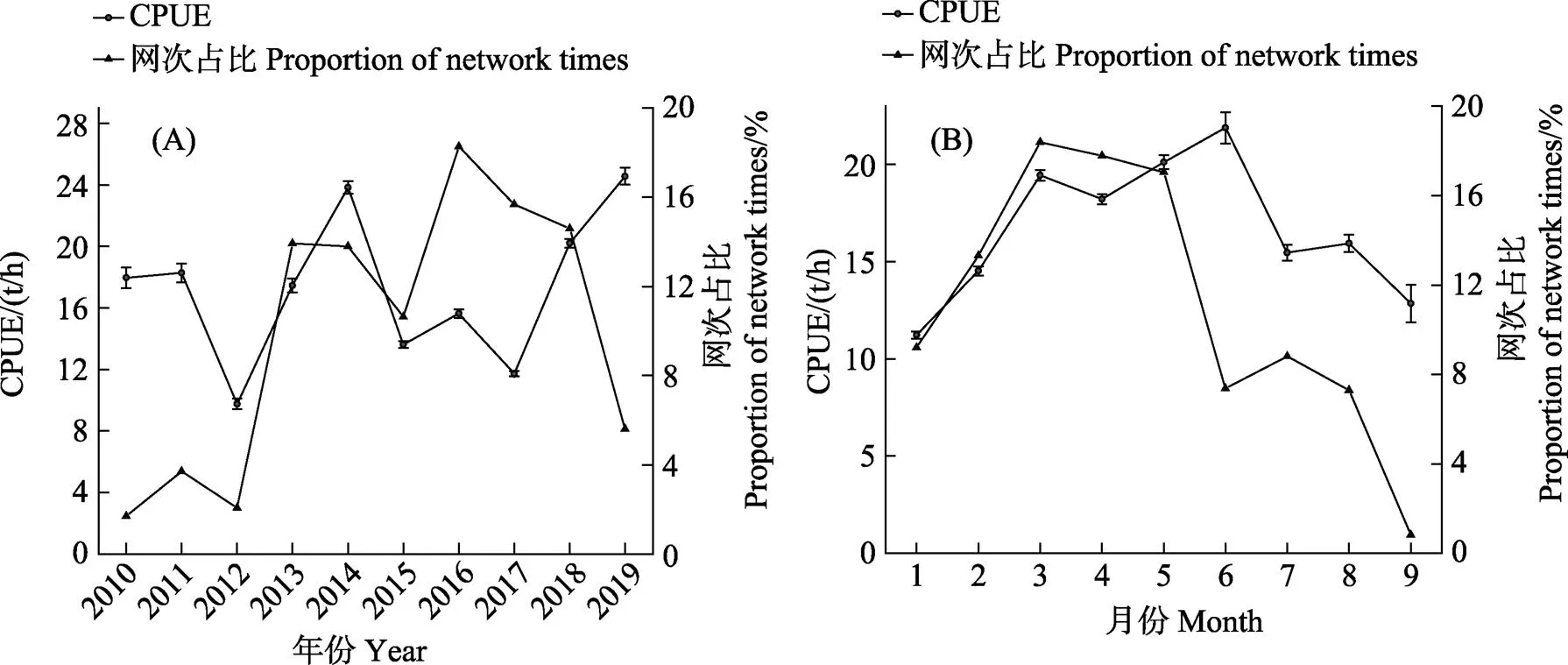
图4 2010—2019年磷虾平均CPUE的年间(A)变化和月间(B)变化
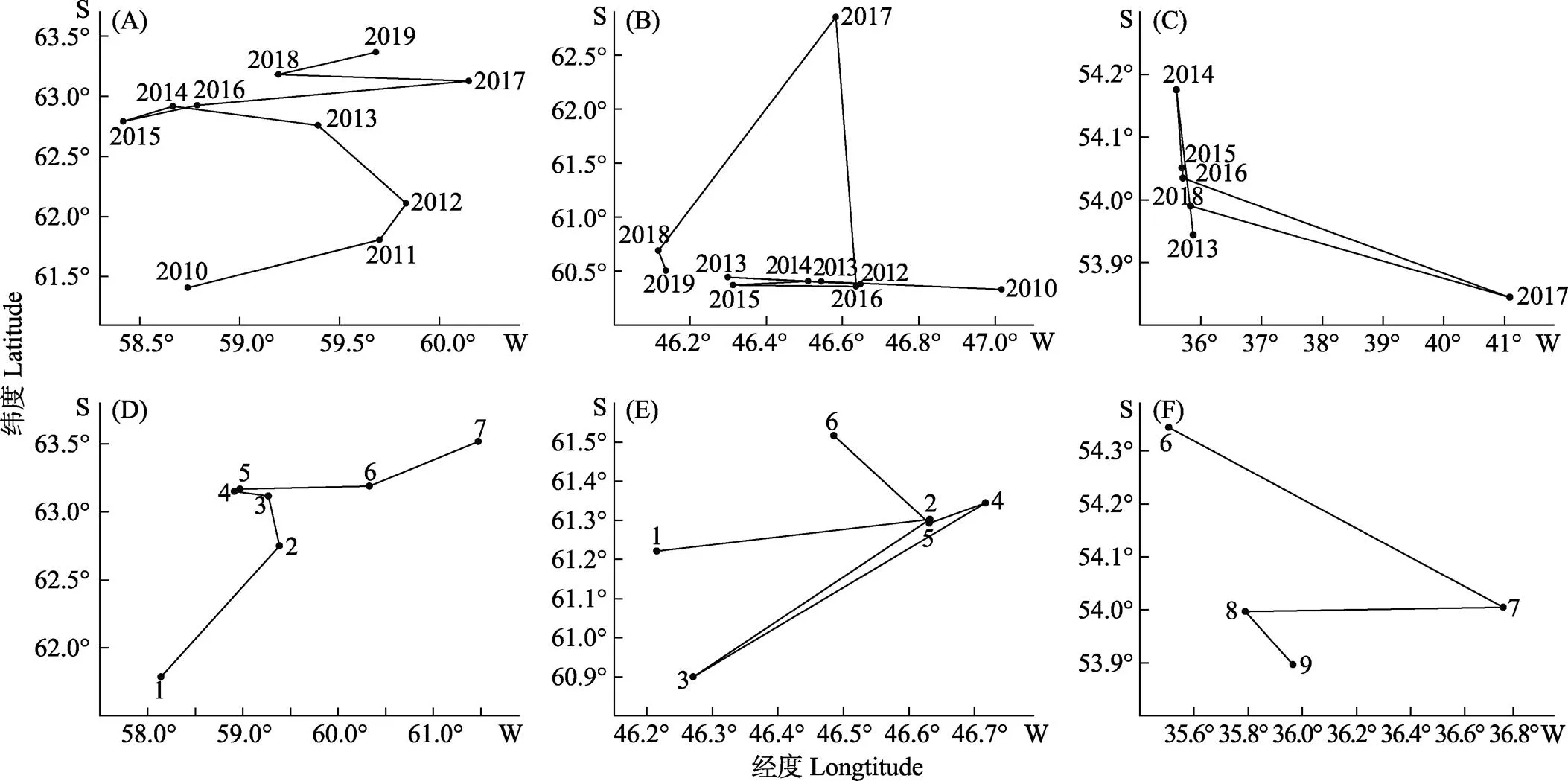
图5 48区磷虾历年产量重心年间(A、B和C)和月间(D、E和F)变化
A、B和C分别表示48.1、48.2和48.3亚区渔场重心的年间变动,D、E和F分别表示48.1、48.2和48.3亚区渔场重心的月间变动。
A, B and C represent the annual variation of fishing ground gravity center in sub region 48.1, 48.2 and 48.3 respectively; D, E and F represent the monthly variation of fishing ground gravity center in sub region 48.1, 48.2 and 48.3 respectively.
从月间变化来看,48.1亚区产量重心总体上逐渐向西南方向移动(图5D);48.2亚区的产量重心随月份无明显的变动规律,产量重心集中在46.20°W~46.75°W、60.90°S~61.55°S范围内,变动范围较小(图5E);48.3亚区仅在6—9月有产量存在,产量重心总体上逐渐向西北移动(图5F)。
2.3 磷虾渔场变动方向和分布格局
为进一步揭示48区磷虾渔场的变动方向和分布格局,采用SDE法对磷虾的重心迁移轨迹进行了互补分析。如图6A所示,对2010—2019年10年的数据进行汇总分析可得,从分布方向上看,磷虾资源在48.1亚区基本上沿布兰斯菲尔德海峡分布,在48.2亚区则分布在南奥克尼群岛偏东方向,在48.3亚区分布在南乔治亚群岛东北侧。另外,通过椭圆的扁率、面积、长轴和短轴可知,磷虾资源在48.1亚区的分布范围最广,离散程度最大;在48.3亚区的方向性最强,向心力最明显。
图6B、6C、6D和表1分别展示了48.1、48.2和48.3亚区2010—2019年每年磷虾渔场的变动情况、产量分布SDE形状的相关参数。48.1亚区磷虾的渔场重心逐渐从西北往东南迁移到布兰斯菲尔德海峡内,年间扩张海域面积差距较大,在2017年达到最大值。除2011年外,渔场分布方位角均在54.05°~87.46°之间变动,表明磷虾渔场分布呈东南–西北格局,2015、2017年的扁率较大,表明2015年和2017年的方向性较强。
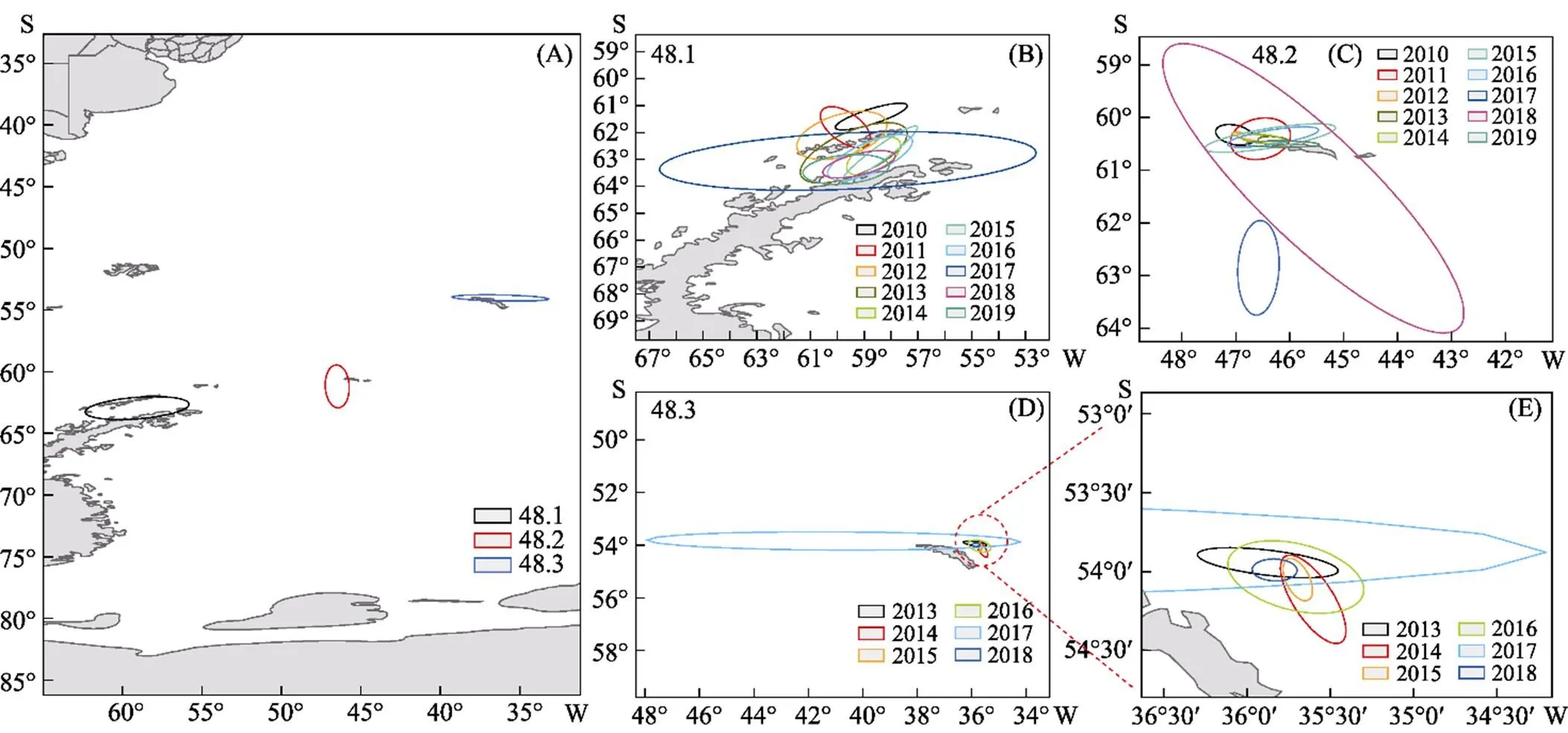
图6 2010—2019年磷虾产量分布标准差椭圆
表1 2010—2019年磷虾产量分布标准差椭圆形状参数
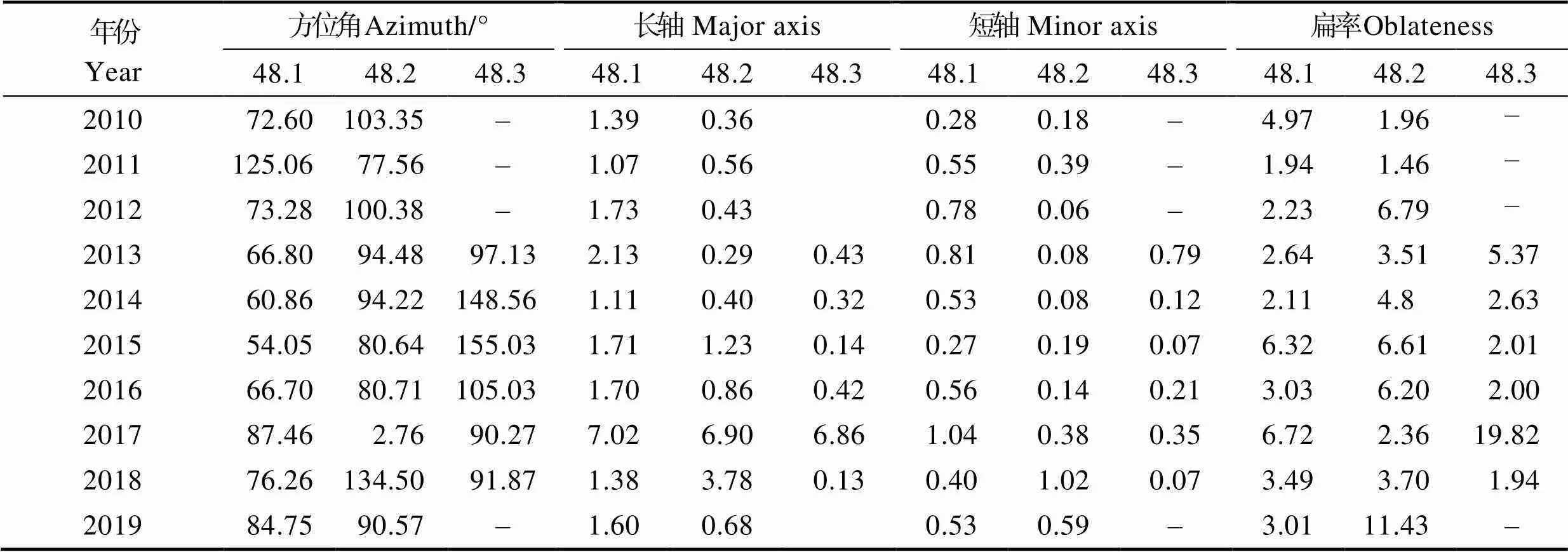
Tab.1 Variation of parameters of standard deviational ellipse of production distribution of E. superba from 2010 to 2019
注:作图时使用经纬度作为度量单位,因此,短轴和长轴只用来提供距离比例和计算扁率。
Note: Latitude and longitude are used as the unit of measurement in drawing, so the minor axis and major axis only provide distance proportion and calculation of flatness.
48.2亚区除2011年和2018年外,渔场重心及扩张海域均在南奥克尼群岛偏东方向,且结合图6A可以得出,48.2亚区的磷虾渔场扩张海域和方向基本受2011年和2017年支配,表明这2年的产量较其他年份占比大。渔场分布方位角的变动范围为2.76°~ 103.35°,年间变动较大,不具有规律,2012、2015年的扁率较大,方向性较强。
48.3亚区除2017年外,其他年份渔场重心及扩张海域均在南乔治亚群岛东北方向,且较为集中,在53°30ʹS~54°30ʹS、35°20ʹW~36°20ʹW之间(图6E),2017年的扩张海域最大,且渔场重心与其他年份均有较大差距,结合图6A可知,渔场重心和扩张海域基本不受2017年影响,因此,2017年的产量占比较小。渔场分布方位角在90.27°~155.03°之间变动,表明渔场基本分布在东西方向,2017年的扁率最大,方向性最强。
2.4 产量重心聚类分析
2010—2019年48.1区不同年份产量重心分布差异较小,由欧式距离可得,年间产量重心的空间距离范围在0.122 (2014年/2016年)~2.223 (2010年/2017年)之间,且欧氏距离超过2的仅有2010年/2017年和2010年/2019年,其他年份之间均小于2(表2)。经聚类分析可得,若以空间距离2为阈值,则可将产量重心分为4类,即2010年为一组,2011年和2012年为一组,2013年、2014年、2015年、2016年和2018年为一组,2017年和2019年为一组(图7A)。48.2亚区2017年产量重心分布与其他年份间差异较大,2017年与其他年份之间的欧式距离均>2,其他年份之间的欧式距离均<1,表明除了2017外,其他年份渔场重心较为集中(表3)。经聚类分析可得,若以空间距离2为阈值,可以将年产量重心分为2类,即2017年为一组,其他年份为一组(图7B)。48.3亚区2017年产量重心分布与其他年份差异较大,均>5,其他年份之间重心的欧式距离均<1,表明除2017年外,渔场重心较为集中(表4)。经聚类分析可得,以空间距离2为阈值,可将年产量重心分为2类,即2017年为一组,其他年份为一组(图7C)。
表2 48.1渔区磷虾各年产量重心的空间距离
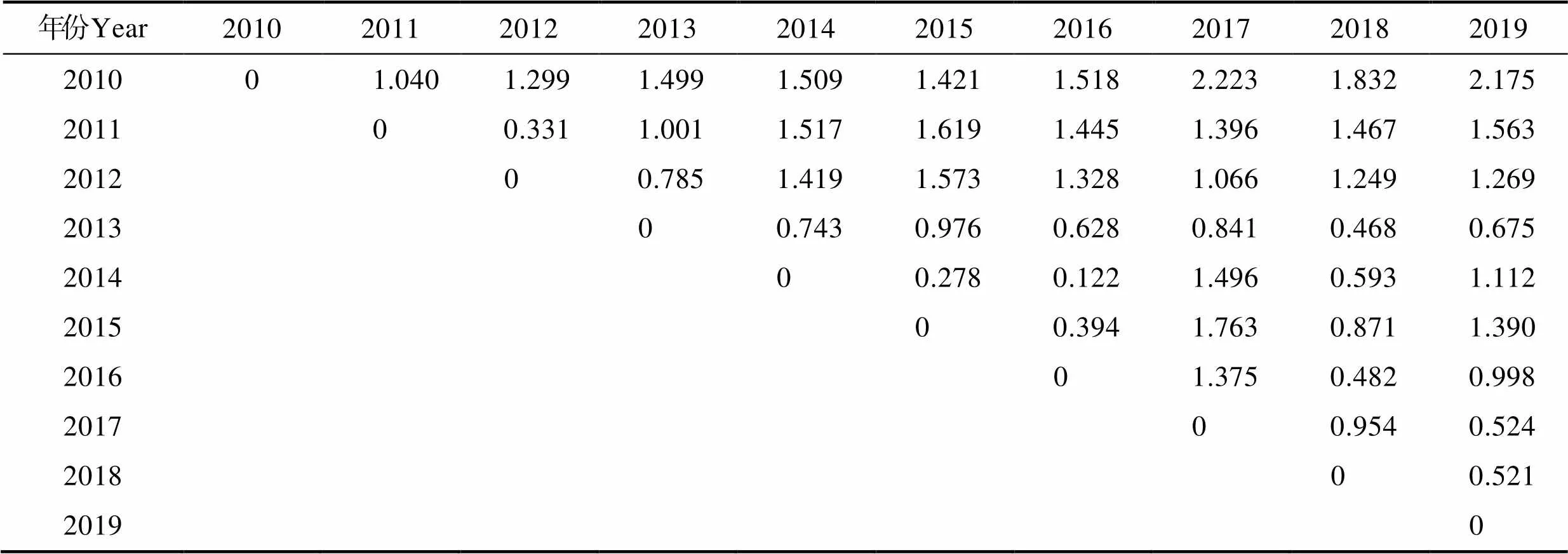
Tab.2 Distribution of annual catch gravity position of E. superba in the 48.1 fishing area
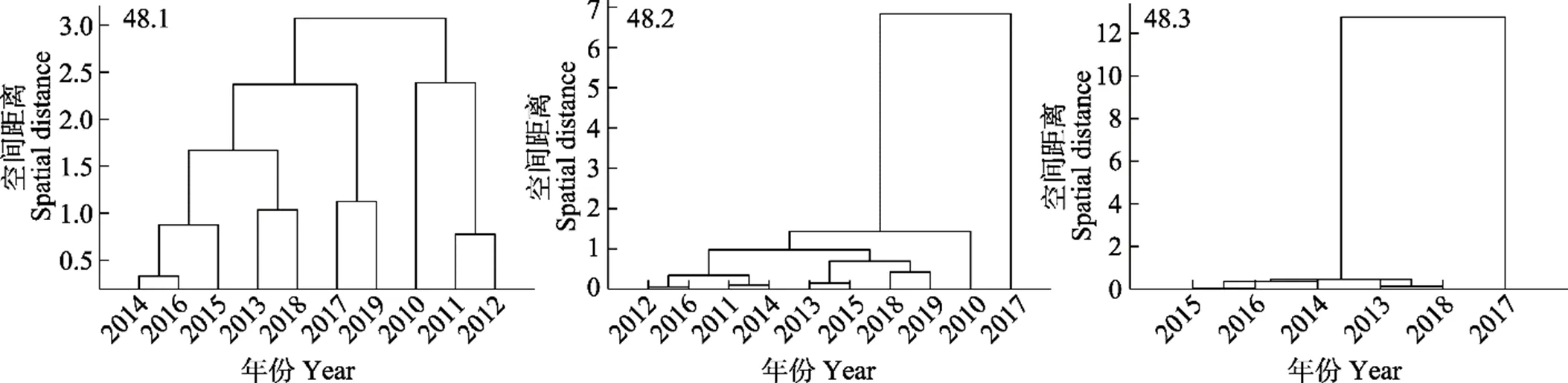
图7 各渔区磷虾年产量聚类结果
表3 48.2渔区磷虾各年产量重心的空间距离

Tab.3 Distribution of annual catch gravity position of E. superba in the 48.2 fishing area
表4 48.3渔区磷虾各年产量重心的空间距离

Tab.4 Distribution of annual catch gravity position of E. superba in the 48.3 fishing area
3 讨论
3.1 磷虾渔场时空变化特征
自20世纪70年代磷虾渔业商业性开发以来,发展迅速,磷虾渔场的时空分布和资源量变动研究等越来越受重视。然而,有关磷虾渔场的年间和季节性分布、资源量变化的研究结果多有分歧(Siegel, 2005)。磷虾渔业活动一直处在变化中,渔场和渔季也随之发生变化。20世纪70年代,磷虾渔业遍布整个南极大陆周围海域,80年代末,印度洋成为磷虾捕捞的主要渔场,90年代开始逐渐向南大西洋转移,自1996年以来,磷虾渔业几乎完全集中在南设得兰群岛、南乔治亚岛和南奥克尼群岛(图8) (Krafft, 2015; Kawaguchi, 2007; CCAMLER, 2020)。自1973年以来,磷虾渔场范围逐渐缩小,捕捞量表现出较强的波动性。总体来看,20世纪90年代之前磷虾的年均总产量要远大于90年代以后的年均产量,但90年代以后,其产量呈现逐年增加的趋势(赵国庆, 2018)。磷虾捕捞量的季节性变化也存在较大波动,20世纪80年代磷虾捕捞产量主要集中在夏季,但自90年代以后,秋冬季磷虾产量比例逐渐增大并占据优势(Kawaguchi, 2006)。赵国庆(2018)通过对以往相关文献和资料总结分析后发现,磷虾全年均可生产。目前,主要捕捞区域位于48区,其中,48.1区作业时间最长,1—6月产量较高;48.2区主要作业时间为3—8月;48.3区主要作业时间为5—10月,其中,5—8月产量较高。李显森等(2015)对2012/2013渔季磷虾的捕捞量进行统计分析后认为,各渔区中CPUE均值以48.1区最高,48.3区最低;月均CPUE值在1—6月较为稳定,7—9月逐渐下降。张瑛瑛等(2020)对中国2010—2015年磷虾捕捞数据分析后发现,磷虾渔场年际重心由南设德兰群岛周边海域向东北方的南奥克尼群岛、南乔治亚群岛周边海域推移,且大多数捕捞作业频次分布于南设德兰群岛夏季时段(1—3月)。Francisco等(2018)通过对各个时空尺度上的捕捞渔船活动情况和磷虾产量分析后发现,历史上的渔场和渔获量的空间分布已经发生了很大变化,并确定了布兰斯菲尔德海峡为主要磷虾渔场。
据南极海洋生物资源养护委员会(Commission for the Conservation of Antarctic Marine Living Resources, CCAMLR)统计,自1996年以来,磷虾的捕捞作业几乎全部集中在48.1、48.2和48.3亚区(CCAMLR, 2020; CCAMLR Secretariat, 2020),因此,本文对该海域的磷虾进行时空分布的研究具有较大的现实意义。磷虾广泛分布于南极大陆周围各个海域,栖息地范围为19×106km2(Atkinson, 2009),密集区基本上出现在大陆架边缘、冰架边缘、海峡以及岛屿周围(Siegel, 2005; Atkinson, 2009)。Patricio等(2020)对智利2011—2016年磷虾捕捞数据分析后发现,48.1区作业范围逐年扩大;48.区的捕捞活动主要发生在南奥克尼群岛西北部,捕捞频次逐年减少;48.3区的捕捞活动主要集中在南乔治亚岛东北部,捕捞范围逐年缩小。这与本文所得出的结论相吻合,即磷虾渔场重心在48.1亚区基本上沿布兰斯菲尔德海峡分布,在48.2亚区则分布在南奥克尼群岛东侧,在48.3亚区分布在南乔治亚群岛东北侧,表明中国与智利的磷虾捕捞区域有一定的重合。在此基础上,本研究运用重心迁移轨迹模型和SDE模型,从更深层次挖掘磷虾渔场重心在各个亚区的移动轨迹和分布格局。2010—2019年磷虾产量和捕捞网次主要集中在48.1亚区,且斯菲尔德海峡的磷虾产量大于48.2亚区和48.3亚区,基本与CCAMLR的统计结果相吻合(CCAMLR, 2020; CCAMLR Secretariat, 2020)。从本研究还可以看出,48.1亚区磷虾渔场的分布范围要远远大于48.2亚区和48.3亚区。
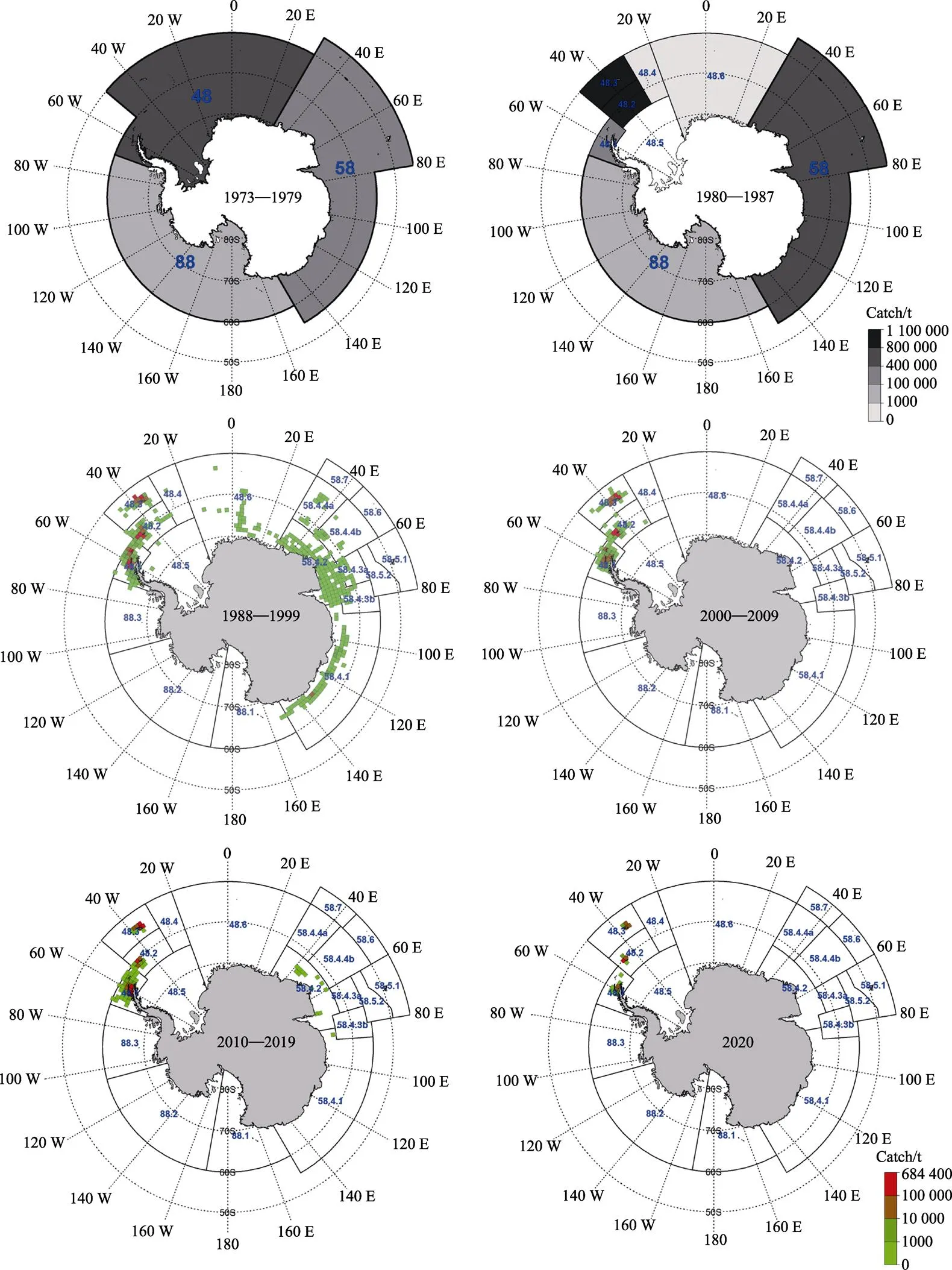
图8 南极海洋生物资源养护委员会统计的磷虾渔获量空间分布(CCAMLR Secretariat, 2020)
3.2 磷虾渔场变动的影响因素
磷虾的商业性捕捞已近50年,磷虾渔业多次被强调为世界上最后未被开发的渔业之一(Garcia, 2011),具有很大的发展潜力,几十年来,磷虾渔场的主要海域、渔季都发生了巨大变化。
磷虾渔业往往跟随磷虾群进行捕捞作业,因此磷虾渔场的选择与其集群行为具有很大关系。集群是磷虾的主要生态行为,在它们的整个生命史中都发挥着重要作用,它们通过集群在某些区域高度集中,但是在某些区域几乎完全没有分布(Murphy, 1988),有时磷虾集合成群时面积可达100万km,内含200万t磷虾(Siegel, 2005)。磷虾的集群和分布是多种因素互相作用的结果,很少有研究能够证明单一因素诸如温度、盐度、溶解氧浓度、溶解有机质、海水理化性质、洋流、锋面系统和经纬度等与磷虾密度或分布具有明显的可预测关系(Siegel, 2016)。磷虾往往因为洋流和水深的共同影响集中分布于陆架边缘和峡谷,可能会密集聚集形成热点区域(Bernard, 2017)。本文与相关研究(Patricio, 2020)发现,近年来,磷虾捕捞多集中于岛屿周围(南奥克尼群岛和南舍得兰群岛)和海峡(布兰斯菲尔德海峡)内。据研究,磷虾生物量的聚集也依赖于潮汐(Bernard, 2017)、风向(Krafft, 2015)的情况,在昼夜潮汐期间,安弗斯岛附近水域的生物量显著更高,当西风为主和局地混合潮处于日变化状态时,大密度聚集更频繁发生。但不得不提的是,磷虾的集群分布难以预测,到目前为止,仍无法准确掌握磷虾的集群机制(Krafft, 2015)。
磷虾渔业是一种利益驱动的经济性活动,渔船通常以利润最大化为目标(Krafft, 2015),磷虾渔场的变迁、渔获量的增减除了受各种物理环境因素的影响外,还与磷虾的商业用途、市场的需求、捕捞渔船的增减和磷虾管理政策等有很大关系(Nicol, 2012)。影响渔船收入的因素有很多,但磷虾集群的密度具有预测性。Krafft等(2015)指出,声学得到磷虾集群密度最高的深度与实际捕捞深度一致,并表示渔船更倾向于在垂直方向上瞄准高密度磷虾群。但有时磷虾渔场的变动可能并不完全由磷虾集群的密度决定。Hewitt等(2004)分析了南设德兰群岛周围海域磷虾捕捞随时间的变化模式后指出,该海域磷虾密度较高的区域主要有象岛东端、象岛和乔治国王岛屿之间和希里夫角附近,但前二者均不是磷虾的主要渔场。磷虾捕捞量的时空波动与船队组成也有一定关联。前苏联自1972年开始在南极水域进行商业捕捞以来一直占据主导地位,年捕捞量超过20万t,主要的捕捞地为南极洲东部海域和南大西洋(Nicol, 2011)。20世纪90年代初期前苏联解体后,磷虾产量大幅下降,主要来自于如本和韩国,后来随着挪威、中国、韩国等国家相继加入磷虾的捕捞,其产量有较大提升,挪威逐渐占据主导地位,磷虾渔场逐渐向南大西洋迁移(陈雪忠等, 2009; 赵宪勇等, 2016; CCAMLR Secretariat, 2020)。
因为磷虾渔船的捕捞活动只发生在无冰区,海冰是影响捕捞渔船作业的主要因素,捕捞热点区域位置与海冰覆盖度关系密切,这也导致磷虾渔场季节性明显(Kawaguchi, 2010b)。另外,海冰为磷虾提供了优良的栖息环境,为越冬的磷虾群提供了优良的索饵环境,也减少了被捕捞和猎食的风险(Brierley, 2002)。据研究,48渔区磷虾夏季的磷虾丰度与上一季度冬季海冰的面积呈正比(Hewitt, 2004),而磷虾渔业中,CPUE与海冰总面积年间变化呈显著负相关关系(戴立峰等, 2012)。近年来,全球气候变暖导致海冰形成延迟、无冰区域扩大,进而使磷虾捕捞的时间跨度变得越来越长,曾只在夏季的捕捞活动现已持续到秋季(Kawaguchi, 2006)。而冰川融化对磷虾的生存具有不容忽视的威胁,据研究,气候引起的冰川融水导致海洋中悬浮颗粒增加,大量的岩性颗粒影响了磷虾的摄食、吸收能力和交配行为,这可能是南极洲波特湾磷虾大量死亡的原因(Fuentes, 2016)。随着全球气候变暖,在过去的90年里,西南大西洋磷虾的分布逐渐向南收缩,在分布的北部极限区域密度急剧下降,密度重心越来越靠近南极冰架(Atkinson, 2019),这可能也是导致48.1亚区磷虾渔场重心逐渐向西南方向移动的主要原因。据研究,在未来几十年里南大洋将会持续变暖(Michael, 2021),而这可能进一步对磷虾的栖息地、分布等产生重要影响。Veytia等(2020)模拟了磷虾栖息地质量的季节性变化,指出未来80年里海洋温度和初级生产力的改变可能会导致磷虾栖息地萎缩,渔场不断向高纬度地区转移。
随着磷虾渔业的发展,磷虾渔场的掌控能力和捕捞技术都有了长足的发展。自2007年以来,挪威将连续泵吸技术应用到磷虾捕捞,磷虾渔获产量持续增加,已达到季节性配额,导致渔场提前关闭(Francisco, 2018)。目前,磷虾渔业多集中在西南大西洋(48区),CCAMLR对48.1、48.2和48.3亚区的捕捞出发限额分别为15.5万t、27.9万t和27.9万t (Nicol, 2012)。48.1亚区作为捕捞热点区域,已多次达到触发水平,对磷虾捕捞时长产生了一定影响(CCAMLR Secretariat, 2020)。目前,CCAMLR以亚区为单位进行捕捞限额的制定,而并未考虑磷虾的聚集特征。南极半岛周边水域,特别是布兰斯菲尔德海峡,作为磷虾重要的产卵场、索饵场和越冬场,大量磷虾聚集在此,捕捞区域和捕捞季节与企鹅和鲸鱼的捕食活动有较大的重合(Weinstein, 2017)。Francisco等(2018)指出,应在较小的空间单元中重新分配预防性捕捞限制,以尽量减少对依赖磷虾的捕食者的不良影响。大约20年前,CCAMLR探索过这种依据小单元进行管理的方法,目前仍有待实施(Hewitt, 2004)。在局部渔获产量不断提升的前提下,这种管理方法很可能会得到采用,而这将进一步对磷虾渔场的时空分布产生影响。
磷虾渔场的选择需要多方面考虑,在进行安全捕捞的前提下,既要严格遵守CCAMLR的管理措施,又需要实现利益最大化,特别是目前气候变化可能已经对南极产生了不可逆的影响,这就要求我们要实时掌握磷虾资源和其生存环境的相关信息,并做出迅捷反应,掌握更先进的泵吸捕捞技术,提升渔场的掌控能力,在世界磷虾渔业中占据有利地位。
ATKINSON A, SIEGEL V, PAKHOMOV E A,. A re-appraisal of the total biomass and annual production of Antarctic krill. Deep-Sea Research Part Ⅰ, 2009, 56(5): 727–740
ATKINSON A, NICOL S, KAWAGUCHI S,. Fittinginto southern ocean foodweb models: A review of data sources and their limitations. CCAMLR Science, 2012, 19: 219–245
ATKINSON A, HILL S L, PAKHOMOV E A,. Krill () distribution contracts southward during rapid regional warming. Nature Climate Change, 2019, 142–149
BERNARD K S, CIMINO M, FRASER W,. Factors that affect the nearshore aggregations of Antarctic krill in a biological hotspot. Deep Sea Research Part Ⅰ: Oceanographic Research Papers, 2017, 126: 139–147
BRIERLEY A S, FERNANDES P G, BRANDON M A,. Antarctic krill under sea ice: Elevated abundance in a narrow band just south of ice edge. Science, 2002, 295(5561): 1890–1890
CARLO R. From the Euclidean distance to compositional dissimilarity: What is gained and what is lost. Acta Oecologica, 2021, 111: 1–3
CCAMLR. CCAMLR statistical bulletin volume 32. Tasmania, Australia: Commission for the conservation of Antarctic marine living resources, 2020
CCAMLR Secretariat. Fishery report 2020:in Area 48. Tasmania, Australia: Commission for the conservation of Antarctic marine living resources, 2020
CHEN M F, LIU F, ZHAO Y L,. The expansion and gradient evolution of impervious surface within the Guangdong-Hong Kong-Macao Greater Bay Area from 1987 to 2017. Journal of Geo-Information Science, 2021, 23(3): 443–455 [陈明发, 刘帆, 赵耀龙, 等. 1987—2017年粤港澳大湾区城市群的不透水地表扩张与梯度演化. 地球信息科学学报, 2021, 23(3): 443–455]
CHEN X Z, XU Z L, HUANG H L,. Development strategy on Antartic krill resource utilization in China. Journal of Fishery Sciences of China, 2009, 16(3): 451–458 [陈雪忠, 徐兆礼, 黄洪亮, 等. 南极磷虾资源利用现状与中国的开发策略分析. 中国水产科学, 2009, 16(3): 451–458]
DAI L F, ZHANG S M, FAN W. The abundance of Antarctic krill and its relationship to sea ice and sea surface temperature. Chinese Journal of Polar Research, 2012, 24(4): 352–360 [戴立峰, 张胜茂, 樊伟. 南极磷虾资源丰度变化与海冰和表温的关系. 极地研究, 2012, 24(4): 352–360]
ERICSON J A, HELLESSEY N, NICHOLS P D,. Seasonal and interannual variations in the fatty acid composition of adultDana, 1850 () samples derived from the Scotia Sea krill fishery. Journal of Crustacean Biology, 2018, 38(6): 1–11
FRANCISCO S C, BILLY E, ARATA J A,. Spatial and temporal dynamics of the Antarctic krill fishery in fishing hotspots in the Bransfield Strait and South Shetland Islands. Fisheries Research, 2018, 208: 157–166
FUENTES V, ALURRALDE G, MEYER B,. Glacial melting: An overlooked threat to Antarctic krill. Scientific Reports, 2016, 1–6
GARCIA S M, ROSENBERG A A. Food security and marine capture fisheries: Characteristics, trends, drivers and future perspectives. Philosophical Transactions of the Royal Society B, 2010, 365(1554): 2869–2880
HEWITT R P, WATKINS J, NAGANOBU M,. Biomass of Antarctic krill in the Scotia Sea in January/February 2000 and its use in revising an estimate of precautionary yield. Deep-Sea ResearchⅡ, 2004, 51: 1215–1236
KAWAGUCHI S, NICOL S, TAKI K,. Fishing ground selection in the Antarctic krill fishery: Trends in patterns across years, seasons and nations. CCAMLR Science, 2006, 13: 117–141
KAWAGUCHI S, NICOL S. Learning about Antartic krill from the fishery. Antarctic Science, 2007, 19(2): 219–230
KAWAGUCHI S, CANDY S G. Quantifying movement behaviour of vessels in the Antarctic krill fishery. CCAMLR Science, 2009, 16: 131–148
KAWAGUCHI S, KING R, MEIJERS R,. An experimental aquarium for observing the schooling behaviour of Antarctic krill (). Deep-Sea Research PartⅡ, 2010a, 57(7–8): 683–692
KAWAGUCHI S, NICOL S, PRESS A J. Direct effects of climate change on the Antarctic krill fishery. Fisheries Management and Ecology, 2010b, 16: 424–427
KRAFFT B A, SKARET G, KNUTSEN T. An Antarctic krill () hotspot: Population characteristics, abundance and vertical structure explored from a krill fishing vessel. Polar Biology, 2015, 38(10): 1687–1700
LEFEVER D W. Measuring geographic concentration by means of the standard deviational ellipse. American Journal of Sociology, 1926, 32(1): 88–94
LEHODEY P, BERTIGNAC, HAMPTON J,. El Nino Southern Oscillation and tuna in the western Pacific. Nature, 1997, 389(6652): 715–718
LI X S, ZHANG J C, ZHAO X Y,. Spatiotemporal distribution ofin CCAMLR Area 48 during 2012/2013 fishing season. Progress in Fishery Sciences, 2015, 36(4): 1–9 [李显森, 张吉昌, 赵宪勇, 等. 2012/2013渔季CCAMLR 48区南极磷虾()资源时空分布. 渔业科学进展, 2015, 36(4): 1–9]
MARRARI M, DALY K L, HU C. Spatial and temporal variability of SeaWiFS chlorophylldistributions west of the Antarctic Peninsula: Implications for krill production. Deep-Sea Research PartⅡ, 2008(55): 377–392
MICHAEL K, SUBERG LA, WESSELS W,. Facing Southern Ocean warming: Temperature effects on whole animal performance of Antarctic krill (). Zoology, 2021, 146: 1–31
MURPHY E J, MORRIS D J, WATKINS J L,. Scales of interaction between Antarctic krill and the environment. Springer Berlin Heidelberg, 1988
NICOL S, FOSTER J. Recent trends in the fishery for Antarctic krill. Aquatic Living Resources, 2003, 16(1): 42–45
NICOL S, FOSTER J, KAWAGUCHI S. The fishery for Antarctic krill: Recent developments. Fish and Fisheries, 2011, 13(1): 30–40
PATRICIO M A, RENZO R, ÁLVARO D C. Chilean Antarctic krill fishery (2011–2016). Latin American Journal of Aquatic Research, 2020, 48(2): 179–196
Polar Special Office of State Oceanic Administration. Investigation and assessment of krill and other biological resources in the waters around Antarctica. Beijing: Ocean Press, 2016, 3 [国家海洋局极地专项办公室. 南极周边海域磷虾等生物资源考察与评估. 北京: 海洋出报社, 2016, 3]
SAENZ B T, AINLEY D G, DALY K L,. Drivers of concentrated predation in an Antarctic marginal-ice-zone food web. Scientific Reports, 2020, 10(1): 1–17
SEOK G C, EUN A Y, DOO H A,. Characterization of frequency and aggregation of the Antarctic krill () using acoustics. Ocean Science Journal, 2018, 53(4): 1–11
SIEGEL V. Distribution and population dynamics of: Summary of recent findings. Polar Biology, 2005, 29(1): 1–22
SIEGEL V. Biology and ecology of Antarctic krill. Advances in Polar Ecology. Springer International Publishing, 2016, 22– 28
STOWASSER G, ATKINSON A, MCGILL R,. Food web dynamics in the Scotia Sea in summer: A stable isotope study. Deep Sea Research PartⅡ, 2012, 59(1): 208–221
VEYTIA D, CORNEY S, MEINERS K M,. Circumpolar projections of Antarctic krill growth potential. Nature Climate Change, 2020, 10(6): 1–8
WANG L Z, XUE C H, WANG Y M,. Extraction of proteins with low fluoride level from Antarctic krill () and their composition analysis. Journal of Agricultural and Food Chemistry, 2011, 59(11): 6108–6112
WEINSTEIN B G, DOUBLE M, GALES N,. Identifying overlap between humpback whale foraging grounds and the Antarctic krill fishery. Biological Conservation, 2017, 210: 184–191
ZHANG J C, ZHAO X Y, WANG X L,. Numericalization of pictorial image of Antarctic krill echogram taken from a commercial echo-sounder. Progress in Fishery Sciences, 2012, 33(4): 64–71 [张吉昌, 赵宪勇, 王新良, 等. 商用探鱼仪南极磷虾声学图像的数值化处理. 渔业科学进展, 2012, 33(4): 64–71]
ZHANG H, ZHANG S M, CUI X S,. Spatiotemporal dynamics in the location of the fishing grounds and catch per unit effort (CPUE) for Chilean jack mackerel (Nichols, 1920) from Chinese trawl fleets on the high seas of the Southeast Pacific Ocean, 2001–2010. Journal of Applied Ichthyology, 2015, 31(4): 646–656
ZHANG Y Y, XU B, ZHANG H,. Interannual and monthly variations of catch per unit effort and the relation with sea surface temperature and chlorophyll concentration in fishing grounds (Fishing Area 48) of Antarctic krill. Chinese Journal of Ecology, 2020, 39(5): 1685–1694 [张瑛瑛, 徐博, 张衡, 等. 南极磷虾渔场(48渔区)CPUE的年、月变化及其与海表温度、叶绿素浓度的关系. 生态学杂志, 2020, 39(5): 1685–1694]
ZHAO G Q. Study on the spatial-temporal distribution of Antarctic krill () fishing ground and its relationship with environmental factors. Master´s Thesis of Yantai University, 2018, 6 [赵国庆. 南极磷虾在渔场中的分布及与环境因子之间关系研究. 烟台大学硕士研究生学位论文, 2018, 6]
ZHAO X Y, ZUO T, LENG K L,. Engineering science and technology challenges in the Antarctic krill fishery. Engineering Sciences, 2016, 18(2): 85–90 [赵宪勇, 左涛, 冷凯良, 等. 南极磷虾渔业发展的工程科技需求. 中国工程科学, 2016, 18(2): 85–90]
Temporal and Spatial Distribution of Antarctic Krill in 48 Fishing Areas Based on Fishery Data
ZHAO Guoqing1,2, LUO Junrong3, TANG Fenghua2, FAN Wei2, SONG Xuefeng2, YANG Chao1,2, ZHANG Heng2①
(1. College of Marine Sciences, Shanghai Ocean University, Shanghai 201306, China; 2. East China Sea Fisheries Research Institute, Chinese Academy of Fishery Science, Shanghai 200090, China;3. Fujian Zhengguan Fishery Development Coperation, Fuzhou, Fujian 350500, China)
Antarctic krill () is a vital part of the food web, supporting substantial fishery in the Southern Ocean. Krill represent the world´s largest animal protein pool, rich in lipids, nutritional long-chain polyunsaturated fatty acids, mineral elements, and carotene, with great potential for development. It is of great significance to understand the changes in fishing grounds, as they profoundly affect the krill population. The distribution characteristics of Antarctic krill fishing grounds in 48 areas were analyzed based on the production statistics of Antarctic krill fishing vessels in China from 2010 to 2019. The gravity center migration trajectory model and standard deviational ellipse (SDE) model were used to analyze the trends and characteristics of Antarctic krill fishing grounds. The results showed that the production of Antarctic krill was mainly concentrated in sub-region 48.1, accounting for 70.30% of total production. The yields of the 48.2 and 48.3 sub-regions showed little difference, accounting for 14.28% and 15.42%, respectively. The annual catch per unit effort (CPUE) curve increased over time, with a minimum value in 2012 and a maximum value in 2019. The monthly CPUE first increased and then decreased, with the minimum value in January and the maximum value in June. The annual and monthly gravity center of Antarctic krill in sub-region 48.1 moved southwest. The annual gravity center of Antarctic krill in sub-region 48.2 moved to the east, but the range of movement was small, and the monthly trends were weak. The annual gravity center of Antarctic krill in sub-region 48.3 moved to the south, and the monthly gravity center moved to the northwest. According to SDE analysis, krill fishing grounds in sub-region 48.1 had the widest distribution and the greatest dispersion, with the strongest directivity and the most obvious centripetal force in sub-region 48.3. The center of gravity of the fishing ground was distributed along the Bransfield Strait in sub-region 48.1, east of the South Orkney Islands in sub-region 48.2, and northeast of the South Georgia Islands in sub-region 48.3. Clustering results showed that the annual fishing ground gravity centers were concentrated in sub-region 48.1. The annual fishing ground gravity centers of sub-regions 48.2 and 48.3 were relatively concentrated in all years, except in 2017.
Antarctic krill; Fishing ground gravity center; Standard deviation ellipse analysis; Cluster analysis; Spatial and temporal distribution; 48 fishing areas
ZHANG Heng, E-mail: zhangziqian0601@163.com
10.19663/j.issn2095-9869.20210407004
S931
A
2095-9869(2022)04-0081-12
*国家重点研发计划(2018YFC1406802)资助 [This work was supported by National Key Research and Development (2018YFC1406802)].赵国庆,E-mail: zgq617717@163.com
张 衡,副研究员,E-mail: zhangziqian0601@163.com
2021-04-07,
2021-06-09
http://www.yykxjz.cn/
赵国庆, 罗俊荣, 唐峰华, 樊伟, 宋学锋, 杨超, 张衡. 基于渔业数据的南极磷虾48渔区渔场时空分布. 渔业科学进展, 2022, 43(4): 81–92
ZHAO G Q, LUO J R, TANG F H, FAN W, SONG X F, YANG C, ZHANG H. Temporal and spatial distribution of Antarctic krill in 48 fishing areas based on fishery data. Progress in Fishery Sciences, 2022, 43(4): 81–92
(编辑 冯小花)
猜你喜欢
当代水产(2022年7期)2022-09-20
今日农业(2022年14期)2022-09-15
小哥白尼·野生动物画报(2022年4期)2022-05-31
意林(2021年2期)2021-02-08
大自然探索(2021年12期)2021-02-07
当代贵州(2020年26期)2020-08-21
传奇故事(上旬)(2019年10期)2019-11-16
电脑知识与技术(2018年20期)2018-11-15
金山(2018年8期)2018-11-09
考试周刊(2016年37期)2016-05-30
