
颗粒浓度对文蛤、硬壳蛤和菲律宾蛤仔保留效率的影响*
2022-09-05 01:41林志华
渔业科学进展 2022年4期
谢 彬 林志华 何 琳
颗粒浓度对文蛤、硬壳蛤和菲律宾蛤仔保留效率的影响*
谢 彬1,2林志华2,3何 琳2①
(1. 宁波大学海洋学院 浙江 宁波 315100;2. 浙江万里学院生物与环境学院 浙江省水产种质资源高效利用技术研究重点实验室 浙江 宁波 315100;3. 浙江万里学院 宁海海洋生物种业研究院 浙江 宁波 315100)
本研究通过在养殖水体中添加底泥或单胞藻调节悬浮颗粒浓度,研究了悬浮物数量浓度和质量浓度变化对文蛤()、硬壳蛤()和菲律宾蛤仔()保留效率的影响。结果显示,文蛤、硬壳蛤及菲律宾蛤仔的保留效率分别在粒径为8、6和6 μm时达到最大值,分别为51.1%、59.6%和62.6%。随着数量浓度的增加,文蛤在低(4.32× 107cells/L)、中(5.27×107cells/L)、高(6.65×107cells/L) 3个数量浓度下保留效率达到最大值时的最小粒径逐渐增大,分别为9、13和14 μm,保留效率最大值分别降至49.7%、33.4%和26.2%;与文蛤相似,菲律宾蛤仔保留效率达到最大值时的最小粒径也分别增大至9、12和14 μm,但最大保留值无明显变化;硬壳蛤保留效率的最大值保持不变,但达到最大值时的最小粒径略有增大,分别为8、9和10 μm。随质量浓度的增加,文蛤和菲律宾蛤仔在低(5.7 mg/L)、中(11.8 mg/L)、高(23.3 mg/L) 3个质量浓度下的保留效率最大值和达到最大值时的最小粒径均无明显变化;硬壳蛤保留效率最大值显著降低,分别为60.7%、27.6%和25.5%,但保留效率达到最大值时的最小粒径保持不变。研究表明,文蛤和菲律宾蛤仔的保留效率对食物颗粒数量浓度变化更敏感,而硬壳蛤的保留效率受质量浓度变化影响较大。
悬浮颗粒浓度;保留效率;文蛤;硬壳蛤;菲律宾蛤仔
滤食性贝类通过鳃丝和其上纤毛的组合运动将流过鳃部的水中的颗粒物质保留下来,被截留下来的颗粒与流过鳃部水中颗粒的比值称为保留效率(retention efficiency)。颗粒保留效率在研究滤食性贝类的营养需求和能量收支时是非常重要的,鳃输送的水量和颗粒保留效率决定了贝类可利用的食物量。近年来,贝类养殖产业发展迅速,但存在无序放苗、超容量养殖等问题(侯兴等, 2021)。养殖容量是保障贝类养殖绿色可持续发展的重要依据,颗粒保留效率是评估贝类养殖容量的重要参数之一。一些以能量转化为基础的贝类养殖容量估算模型(方建光等, 1996; Officer, 1982; Dame, 1997)需要测定滤水率(潘鲁青等, 2002),而实际中常以清滤率(董世鹏等, 2020; 林元烧等, 2003)来替代滤水率。只有当贝类对滤过水体中颗粒物的保留效率为100%时,清滤率才完全等于滤水率,否则清滤率总是小于滤水率。因此,可以认为颗粒保留效率决定了养殖容量估算模型参数的准确性,在养殖容量估算过程中至关重要。
保留效率并不是一成不变的,也可能受多种环境因子的影响,特别是与水体中悬浮颗粒物数量以及质量浓度之间的关系尚不完全清楚。数量浓度表示单位体积的水体中所含悬浮颗粒物的数量,而质量浓度是表示单位体积的水体中所含有机颗粒物的质量。Stenton-Dozey等(1992)研究发现,岩池蛤()保留效率受潮汐影响,在低潮期对5~9 μm颗粒的保留率最大,而在高潮期对8~13 μm颗粒的保留率最大,这与颗粒物有机含量变化规律相一致。Maurice (1985)研究报道,长牡蛎()对粒径>6 μm颗粒的保留效率达到100%。Barillé等(1992)研究认为,长牡蛎保留效率受悬浮颗粒数量浓度影响,而不受质量浓度的影响。相反,美洲牡蛎()和海湾扇贝()的保留效率只受颗粒食物质量浓度的影响(Palmer, 1980)。相比之下,悬浮颗粒物数量浓度和质量浓度均能影响栉孔扇贝()的保留效率(Zhang, 2010)。另有研究显示,食物颗粒数量浓度增大也会对欧洲食用牡蛎()的保留效率产生负面影响(Wilson, 1983)。同时,有研究认为,贝类的保留效率与食物颗粒浓度之间没有相关性(Davids, 1964; Hildreth, 1980)。Lucas等(1987)比较了生长在不同颗粒浓度环境中紫贻贝()的保留效率,结果显示,2个种群的保留效率相同。Zhang等(2010)研究也表明,食物颗粒质量浓度波动对紫贻贝的保留效率无明显影响。这些研究表明,滤食性贝类只能100%保留特定大小的颗粒,但不同贝类所能100%保留的颗粒略有不同,且对颗粒浓度变化的响应趋势存在很大差异,尚无一致的规律。
文蛤()、硬壳蛤()和菲律宾蛤仔()是我国沿海地区养殖规模和产量最大的沙质底埋栖型经济贝类。目前,关于它们的保留效率以及对颗粒食物浓度变化的响应情况尚未见报道。本研究运用流水法测定了文蛤、硬壳蛤和菲律宾蛤仔对自然海水中不同粒径颗粒物的保留效率,以及对颗粒质量浓度和数量浓度变化的短期响应趋势,为滤水率的计算提供更可靠的计算指标,从而可以更准确地评估养殖容量。
1 材料与方法
1.1 实验材料
实验生物(菲律宾蛤仔、文蛤和硬壳蛤)购自宁波路林水产品批发市场,平均壳长分别为(34.0±2.2),(40.5±1.3)和(42.8±2.3) mm。清除表面附着物后,将其放入25℃循环水养殖系统中暂养2 d,用于不同条件下保留效率的测定。
自然海水取自宁波市科技创新基地外,象山港水域,盐度为21.0~21.5,为避免抽水过程中带来的沉积再悬浮颗粒的干扰,经缓冲槽沉淀12 h后再注入蓄水槽。
1.2 实验方法
在贝类养殖区域,悬浮颗粒浓度通常会随着季节变化而改变,同时,也极易受到一些外界因素的影响(如暴雨、潮汐等)。为模拟悬浮颗粒浓度条件变化对贝类保留效率的影响,本实验分为3组,包括了自然海水(natural seawater, NSW),自然海水+单胞藻(NSW+algae),自然海水+表层底泥(NSW+silt)。在自然海水中,通过添加球等鞭金藻()调节颗粒的质量浓度,添加底泥(滩涂区域表层2 cm淤泥)调节颗粒的数量浓度(Barillé, 1992; Zhang, 2010),其中,金藻的直径为4.4~7.1 μm,底泥颗粒的直径为2~20 μm。根据单胞藻的添加量,质量浓度可分为低、中、高3个浓度级别,分别用low-algae、mid-algae和high-algae表示;根据底泥的添加量,数量浓度也分为低、中、高3个浓度级别,分别用low-silt、mid-silt和high-silt表示。流水装置和流水槽参照董世鹏等(2020)进行设计(图1)。每组设置15个流水槽(体积20 cm×5 cm×10 cm),流量设为1.3 L/h,其中10个实验组,每个水槽放置1只实验贝类;5个空白组,不放实验贝类。各实验组悬浮颗粒的质量浓度和数量详细情况见表1。质量浓度指标为叶绿素的浓度(Chl.a)和有机含量[悬浮有机颗粒物浓度(POM)和总悬浮颗粒物浓度(TPM)比,POM/TPM)],3个质量浓度级别颗粒的叶绿素浓度和有机含量分别为39.3、98.2和154.9 μg/L,25.11%、35.18%和59.15%;数量浓度指标为TPM和POM,3个数量浓度级别颗粒的TPM和POM分别为31.1、56.4和79.0 mg/L,7.4、8.3和9.5 mg/L。
为避免生物节律对实验结果的影响,实验时间一致,为07:00~11:00。实验开始时,选取摄食正常的个体放入流水槽,为了防止水流、朝向对其摄食行为的影响(Eckman, 1989),贝类放置的位置、朝向一致,并将流水槽设置在25℃恒温水浴中。实验生物放入水槽中适应1 h直到正常摄食,之后每隔1 h取50 mL流水槽出水处水样(水槽内的水体1 h可全部完成替换),共进行3次重复取样。使用便携式颗粒计数器(PAMAS S40,德国)测定颗粒物浓度,该仪器能检测水样中不同粒径颗粒的数量,以此计算不同粒径颗粒的保留效率。
Chl.a测定采用浮游植物分类荧光仪Phyto-PAM (Walz,德国)。TPM和POM的测定方法:将GF/C玻璃纤维滤纸(Whatman1822-055, 孔径为1.2 μm)在450℃条件下灼烧至恒重(6 h),使用电子天平(Sartorius,精度为0.1 mg,德国)称重,记为0(mg);取300 mL水样,用经450℃灼烧6 h的玻璃纤维滤纸进行抽滤,并用0.5 mol/L的甲酸铵漂洗(10 mL),然后,在60℃烘箱中烘干至恒重(48 h)后称重,记为60(mg);经450℃灼烧6 h后再次称重,记为450。TPM和POM计算公式为:
TPM=60–0
POM=450–60
保留效率(retention efficiency, RE)的计算公式(Cranford, 1992)为:
RE (%)=100%×[1–(2/1)]
式中,1和2分别为空白组和实验组出水处水体中某一粒径悬浮颗粒物的浓度(cells/mL)。
标准保留效率(standardizing retention efficiency, REs)为在一定的粒径范围内,将保留效率最大值视为贝类能够100%保留,某一粒径悬浮颗粒物的保留效率与最大值的比值称为标准保留效率(Eckman, 1989)。
清滤率(clearance rate, CR)的计算公式有2种,在流水中采用Walen (1972)的公式:
CR=×[1–(2/1)]
式中,为水槽中水的流速,1和2分别为水槽入水口和出水口处的饵料浓度。由此可得,清滤率的计算公式也可表示为:
CR=×RE
2 结果
自然海水、自然海水+单胞藻和自然海水+表层底泥各组的质量浓度和数量浓度的详细情况见表1。如表1所示,通过添加不同量的球等鞭金藻来调节悬浮颗粒的质量浓度,其悬浮有机颗粒物浓度和有机含量显著增加(<0.05),同时,数量浓度无显著变化(>0.05);通过添加不同量的底泥来调节悬浮颗粒的数量浓度,其数量浓度显著增加(<0.05),有机颗粒物浓度基本保持不变,但有机含量显著降低(<0.05)。各组别的悬浮颗粒分布情况如图2所示,粒径>20 μm的颗粒的数量非常少。在自然海水中,颗粒浓度一般随粒径增大而逐渐减小,但直径为4 μm颗粒的浓度高于粒径为3 μm的颗粒。不同数量浓度条件下的颗粒分布规律与自然海水中颗粒分布规律相似,添加底泥能够增加2~19 μm范围内所有颗粒的浓度;添加单胞藻调节食物颗粒质量浓度仅能改变4~8 μm颗粒的浓度,对其他直径颗粒的浓度几乎没有影响。
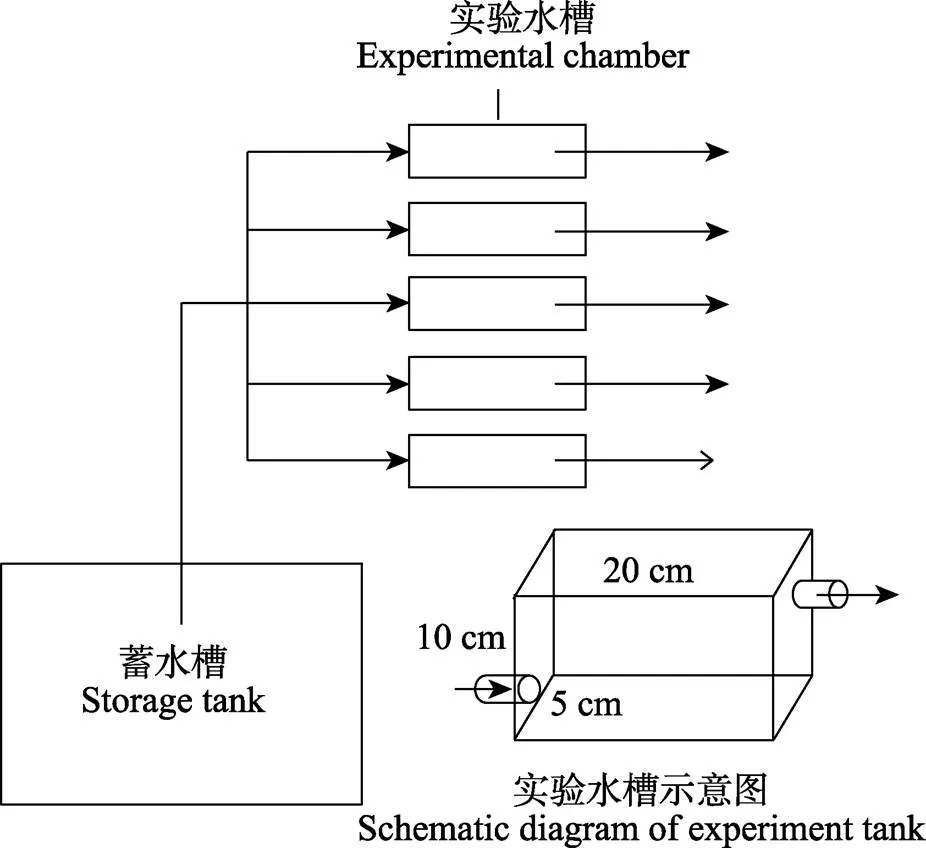
图1 流水系统模拟图
表1 各实验组的颗粒食物的质量浓度和数量浓度参数
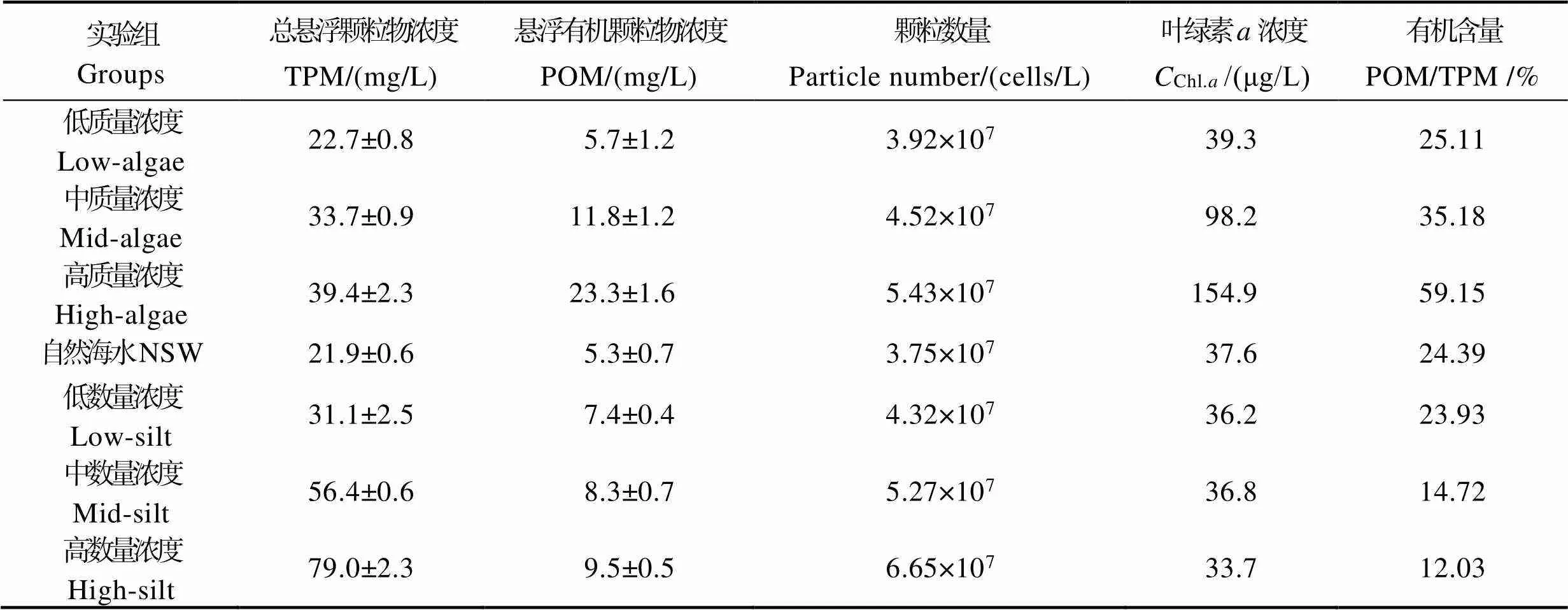
Tab.1 Summary of dietary characterstics during each measurment of retention efficiencies

图2 自然海水+单胞藻和自然海水+底泥各组别的粒径分布情况
A:自然海水+单胞藻;B:自然海水+底泥
A: NSW+algae; B: NSW+silt
2.1 自然海水中文蛤、硬壳蛤和菲律宾蛤仔的保留效率
在自然海水中,文蛤、硬壳蛤和菲律宾蛤仔的保留效率随颗粒粒径的增大逐渐增加,分别在粒径为8、6和6 μm时,保留效率达到最大值,最大保留效率分别为51.1%、59.6%和62.6%,且不再随颗粒的增大而增加。与其他2种贝类相比,菲律宾蛤仔对小颗粒悬浮物的保留效率较高,其对直径2 μm颗粒的保留效率为26.7%,相对应的标准保留效率(REs)为42.9%,文蛤和硬壳蛤对2 μm颗粒的保留效率分别为9.9%和14.6%,REs分别为18.8%和24.4%;对于粒径4 μm的颗粒,文蛤、硬壳蛤和菲律宾蛤仔对其保留率分别为29.5%、42.3%和49.4%,相对应的REs分别为55.9%、70.7%和79.4% (图3)。
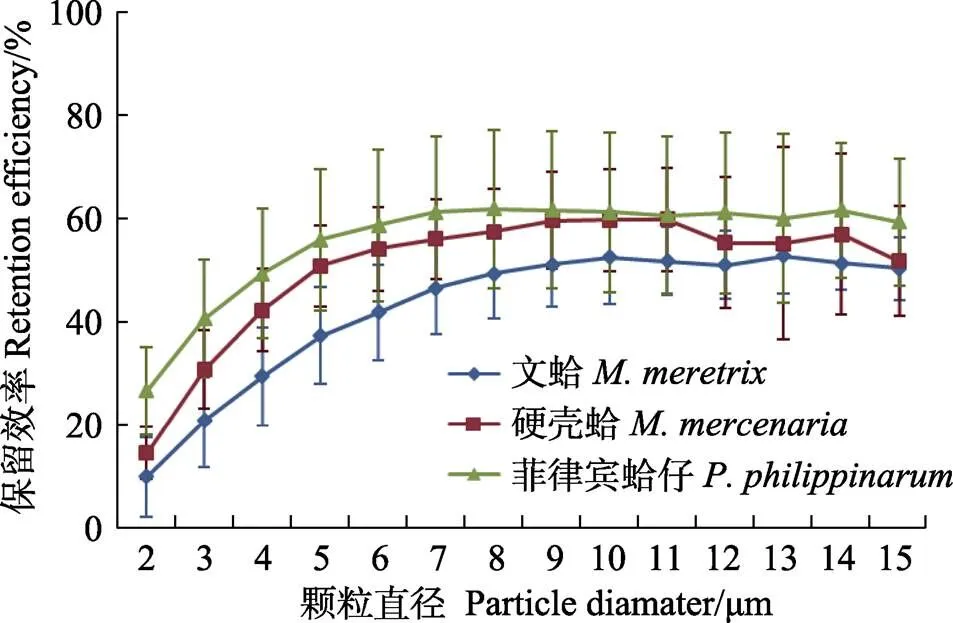
图3 文蛤、硬壳蛤和菲律宾蛤仔在自然海水中对不同粒径颗粒的保留效率
2.2 文蛤在不同颗粒食物条件下的保留效率
文蛤在不同数量浓度和质量浓度条件下的保留效率分别见图4和图5。结果显示,文蛤的保留效率因数量浓度和质量浓度的改变而发生相应的变化。随着数量浓度的增加,文蛤的保留效率逐渐减小,其中2 μm颗粒的保留效率由自然海水中的9.9%降至添加底泥组的–7.8%~–3.9%。自然海水+底泥实验组中low-silt、mid-silt及high-silt 3个数量浓度的最大保留效率由自然海水中的51.3%分别降至49.7%、33.8%和26.2%。同时,保留效率达到最大值时的颗粒粒径也由自然海水中的8 μm分别增至9、13和14 μm。
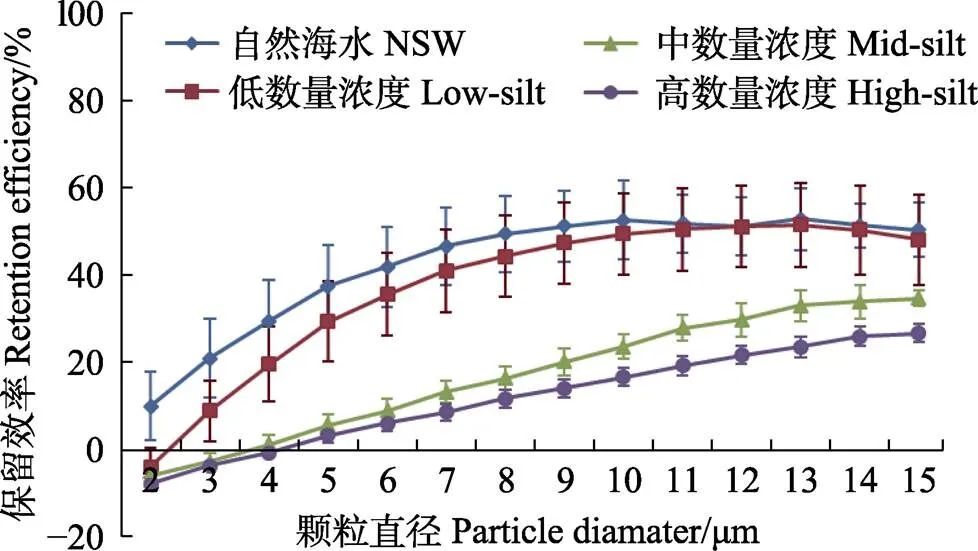
图4 文蛤在自然海水+底泥实验组3个浓度级别中对不同粒径颗粒的保留效率

图5 文蛤在自然海水+单胞藻实验组3个浓度级别中对不同粒径颗粒的保留效率
在自然海水+单胞藻实验组的low-algae条件下,悬浮颗粒的数量浓度和质量浓度变化较小,但保留效率变化明显。与自然海水组相比,文蛤的最大保留效率基本保持不变(由自然海水组的51.3%降为47.8%),达到保留效率最大值的颗粒粒径也由8 μm略微增大至9 μm;此外,在颗粒粒径为4 μm处有一小的低值,其标准保留效率为39.1%。
在mid-algae条件下,悬浮颗粒的质量浓度指标显著增加,Chl.a由自然海水中的37.6 μg/L增至98.6 μg/L,有机含量也由22.86%增至33.81%。文蛤保留效率的响应趋势与low-algae组相近,颗粒粒径为4 μm处仍有一低值,标准保留效率为38.2%;不同之处在于,保留效率最大值略有降低,由47.8%降至40%,保留效率达到最大值时的颗粒粒径由9 μm增至10 μm。
在high-algae条件下,悬浮颗粒的数量浓度也明显提高,数量浓度指标TPM由21.9 mg/L升至45.8 mg/L。文蛤对粒径4 μm颗粒的保留效率显著降低(<0.05),有非常明显的谷值(图4),保留效率为4%,相对应的标准保留效率仅为7.8%,达到保留效率最大值的颗粒粒径继续增大到11 μm,但保留效率最大值无显著变化(>0.05),最大保留率为50.6%。
2.3 硬壳蛤在不同颗粒食物条件下的保留效率
硬壳蛤在自然海水+底泥组和自然海水+单胞藻组的保留效率分别见图6和图7。在添加底泥条件下,硬壳蛤的保留效率随颗粒粒径的增大而逐渐增大,达到峰值后随颗粒的继续增大而逐渐减小,low-silt、mid-silt和high-silt 3个浓度下硬壳蛤保留效率的最大值无显著差异(>0.05),分别为57.4%、58.2%和56.4%,相对应的保留效率达到最大时的颗粒粒径分别为8、9和10 μm。硬壳蛤在不同质量浓度条件下的保留效率的差异在于小颗粒物的保留率随底泥添加量的增加而降低,例如,mid-silt实验组和high-silt实验组中粒径2 μm颗粒的保留效率由low-silt实验组的17.1%分别降至–7.7%和–11.2%。
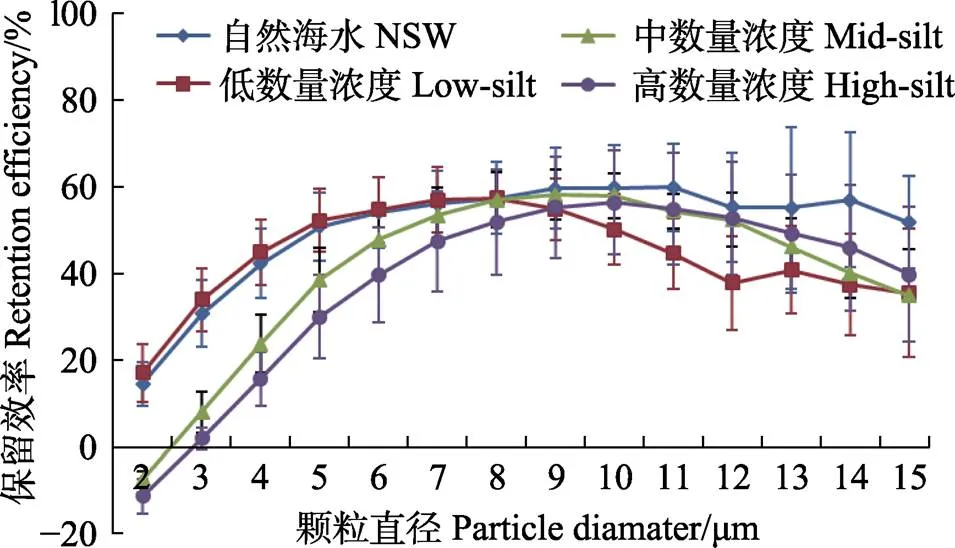
图6 硬壳蛤在自然海水+底泥实验组3个浓度级别中对不同粒径颗粒的保留效率
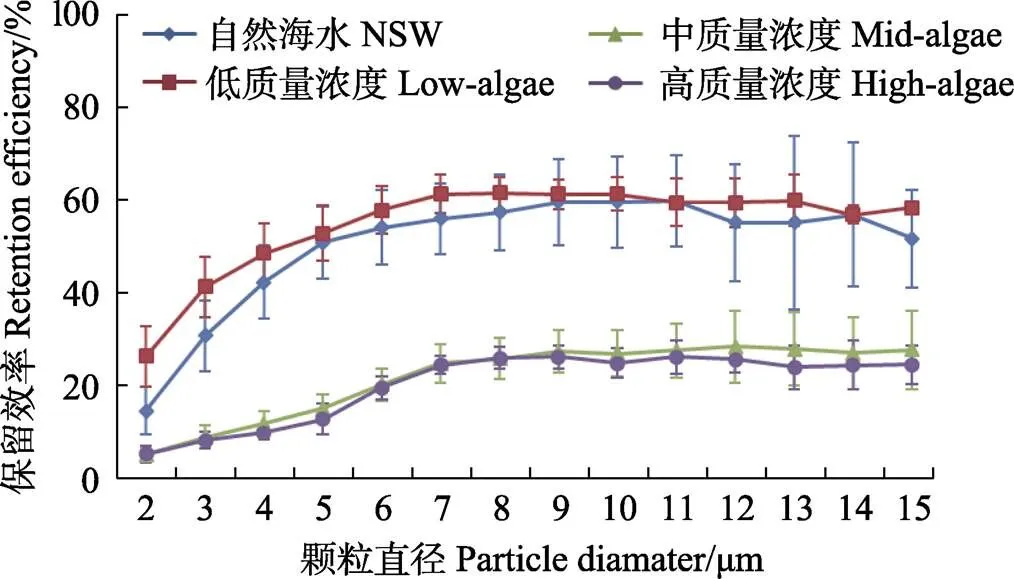
图7 硬壳蛤在自然海水+单胞藻实验组3个浓度级别中对不同粒径颗粒的保留效率
硬壳蛤的保留效率受质量浓度影响较大,随单胞藻添加量的增加,其保留效率最大值显著降低(<0.05)。low-algae实验组保留效率最大值为60.7%,而mid-algae实验组保留效率最大值骤降至27.6%,high-algae实验组保留效率的最大值与mid-algae实验组相近,为25.5%。但3个浓度级别下保留效率达到最大值时的颗粒粒径相同,均为7 μm。另外,在任何浓度条件下,硬壳蛤的保留效率在达到最大值之前都随颗粒增大而增大,不存在谷值。
2.4 菲律宾蛤仔在不同颗粒食物条件下的保留效率
菲律宾蛤仔的保留效率对数量浓度变化的响应趋势与文蛤相似,随数量浓度的增加,小颗粒物的保留效率逐渐减小,保留效率达到最大值时的颗粒粒径逐渐增大,low-silt、mid-silt和high-silt 3个实验组保留效率达到最大值时的颗粒粒径分别为9、12和14 μm。不同之处在于,菲律宾蛤仔保留效率的最大值不随数量浓度变化而改变,分别为56.4%、59.8%和55.0% (图8)。
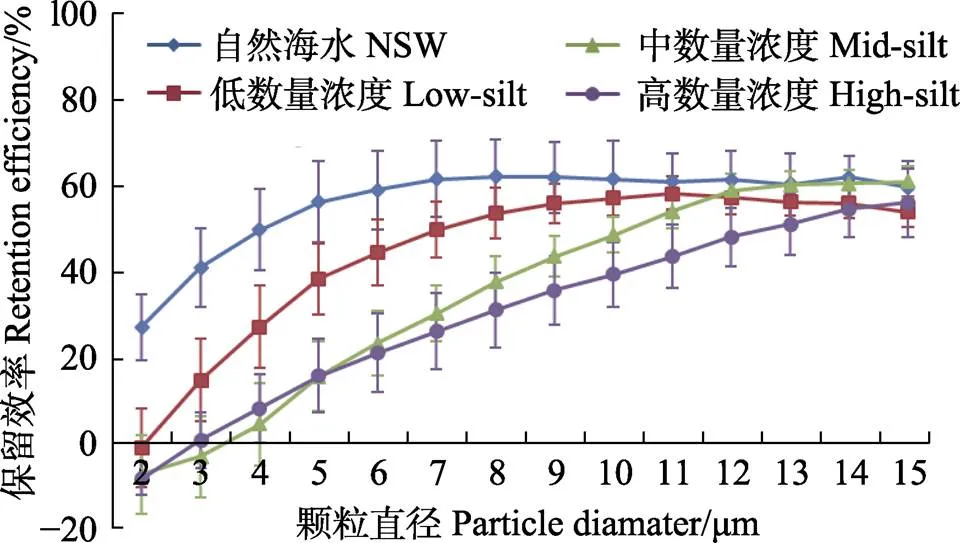
图8 菲律宾蛤仔在自然海水+底泥实验组3个浓度级别中对不同粒径颗粒的保留效率
菲律宾蛤仔的保留效率也因质量浓度的变化发生一定的改变,主要体现在对小颗粒物的保留效率方面。例如,随质量浓度的增加,粒径2~3 μm颗粒的保留效率不断增加,low-algae、mid-algae和high-algae实验组的保留效率分别为55%、60%和67%,其中high-algae实验组中粒径3 μm颗粒的保留效率已经接近保留效率最大值(REs为88%)。然而,保留效率并不随颗粒的增大而逐渐增大,在粒径4~5 μm处会出现一个谷值,这一点与文蛤非常相似,不同的是菲律宾蛤仔在high-algae实验组中的谷值点在5 μm处,当然粒径4 μm颗粒的保留效率也非常低,与5 μm颗粒的保留效率无显著差异(>0.05, 图9)。质量浓度的改变对菲律宾蛤仔保留效率达到最大值时的颗粒粒径影响并不明显,分别为7、7和8 μm。

图9 菲律宾蛤仔在自然海水+单胞藻实验组3个浓度级别中对不同粒径颗粒的保留效率
3 讨论
研究发现,滤食性贝类会因水体中悬浮颗粒物数量浓度和质量浓度(有机含量)的波动而改变他们的摄食行为。为适应水体中颗粒浓度的变化,不同物种对摄食过程有不同的调控机制,包括调节摄食的持续时间和速率、产生假粪、选择性摄食及改变保留效率等(Hawkins, 1990; Bayne, 1993)。
本研究结果显示,悬浮颗粒数量浓度变化能够显著地影响文蛤和菲律宾蛤仔的保留效率,而质量浓度变化对它们保留效率的影响较小,与长牡蛎、紫贻贝相似(Barillé, 1993; Riisgård, 1988)。随着数量浓度的增加,保留效率最大值逐渐降低,并且达到最大值时的颗粒粒径也增大。不仅如此,文蛤和菲律宾蛤仔对小颗粒的保留效率也逐渐减小,甚至为负值。宋强等(2006)研究表明,沉积再悬浮颗粒形成的高浓度浊液会对滤食性贝类的摄食行为产生一定的负面影响,其中包括了滤水率和摄食率等。Jørgensen (1974)研究也发现,贻贝对1.1~2.8 μm颗粒常出现负保留的现象,这是一种非正常状态,可能有几点原因:(1)在摄食过程中,纤毛摆动将大颗粒拍碎而产生了更多的小颗粒。(2)高浓度的悬浮颗粒刺激贝类产生大量的假粪,使测定的颗粒物中包含了假粪颗粒,影响了数据的测定和数值的计算。
另外,虽然悬浮颗粒质量浓度变化对文蛤和菲律宾蛤仔的保留效率的影响较小,但自然海水+单胞藻实验组中的4~5 μm颗粒的保留效率仍出现明显的谷值,同时,保留效率达到最大值的最小粒径也移动到更大的尺寸。保留效率出现谷值很可能是因为添加单胞藻导致粒径4~5 μm颗粒的数量大幅度上升,从而导致了保留效率的降低,也恰好证明了颗粒数量浓度增加将会导致保留效率降低。悬浮颗粒数量浓度增加导致贝类保留效率降低,意味着当养殖水体中某一微藻或某些粒径相近的颗粒食物的数量超过一定浓度时,贝类只能保留一部分颗粒,其余颗粒随水流排出无法被利用,浓度过高甚至会抑制贝类的摄食。生产中,可通过增加饵料丰度、循环利用含有高浓度食物颗粒的水体等方法来促进贝类生长、提高养殖效益。
相比之下,硬壳蛤对悬浮颗粒质量浓度变化更加敏感,不仅小颗粒的保留效率因颗粒浓度而改变,最大保留效率也随浓度变化而改变,但保留效率达到最大值时的颗粒粒径保持不变。因为一些双壳贝类可能通过调整摄食行为来适应环境中食物颗粒浓度的变化以满足生长需求(Bayne, 1987)。一方面,当悬浮颗粒浓度较低时,贝类会增强过滤能力以满足能量需求;另一方面,可能为了避免过量摄食,高浓度下贝类会减小滤水量或保留效率。为满足不同时期的能量需求,贝类可能通过调整鳃的形态结构来影响保留效率。Honkoop等(2003)研究发现,长牡蛎鳃的形态在较长的时间尺度内随时间而变化,而悬浮颗粒浓度也随时间而变化。Barillé等(2000)研究也发现,法国大西洋沿岸高浊度地区的牡蛎比低浊度地区的牡蛎有更少的纤毛和更大的唇瓣。可能正是这种鳃的形态可塑性使贝类具有通过改变保留效率来适应环境中悬浮颗粒浓度变化的能力。本研究中,硬壳蛤的保留效率对食物颗粒浓度变化做出的反应与文蛤和菲律宾蛤仔完全不同,也可能是三者鳃结构的差异所导致的。
在常用的滤水率测定方法中,必须考虑贝类对食物颗粒的保留效率,以便增加实验结果的可靠性。Williams (1982)研究证明,颗粒保留效率达到100%时的粒径是计算滤水率十分重要的参数,取不同粒径值,计算的滤水率有显著差异。本研究也发现,文蛤、硬壳蛤和菲律宾蛤仔3种贝类取不同粒径保留值计算得到的滤水率存在显著差异。例如,在自然海水中,文蛤以粒径为2 μm和大于8 μm颗粒(保留效率已达到最大值)的保留效率计算的滤水率分别为0.13和0.66 L/h,相差5倍;硬壳蛤和菲律宾蛤仔取不同粒径保留值计算的滤水率之间的差距也可达到2~4倍。因此,在测定滤水率时,不能以全部悬浮颗粒的浓度变化作为计算指标,应当以保留效率最大的颗粒的浓度变化作为计算指标,更能反映滤水率的真实情况。然而,实验贝类的保留效率达到最大值时的颗粒粒径因悬浮颗粒质量浓度或数量浓度的变化而改变,变化幅度在6~14 μm内,故悬浮颗粒浓度变化会对滤水率的计算产生较大的影响,应根据颗粒浓度变化选择合适的颗粒用于测定滤水率。
在基于滤水率的贝类养殖容量估算模型中,滤食性贝类滤水率的测定至关重要。若以清滤率直接替代滤水率或由于计算指标选择不合理导致的滤水率偏小,估算的养殖容量将会偏大。例如,李磊等(2014)利用方建光贝类养殖容量估算模型、营养动态模型和Tait沿岸海域能流分析模型估算了文蛤在滩涂的养殖容量,数据显示,方建光贝类养殖容量估算模型的结果显著高于其他2种模型,分别为34 623.90、15 528.45和25 949.10 kg/hm2,且由于浮游动物对初级生产力的消耗,方建光模型估算出来的贝类养殖容量数值偏大。当然,这也并不意味着其他2种模型估算的结果就十分准确。杨淑芳等(2016)利用营养动态模型、Tait沿岸海域能流分析模型估算的菲律宾蛤仔的养殖容量同样大于实际养殖容量。因此,在测定模型参数时,应充分考虑食物颗粒的保留效率对测定滤水率的影响,通过提高滤水率测定方法的客观性和准确性对养殖容量估算模型进行修正,提升养殖容量评估方法的准确性和适用性。
BARILLÉ L, HAURE J, COGNIE B,. Variations in pallial organs and eulatero-frontal cirri in response to high particulate matter concentrations in the oyster. Canadian Journal of Fisheries and Aquatic Sciences, 2000, 57(4): 837–843
BARILLÉ L, PROU J, HÉRAL M,. No influence of food quality, but ration-dependent retention efficiencies in the Japanese oyster. Journal of Experimental Marine Biology and Ecology, 1993, 171(1): 91–106
BARILLÉ L, PROU J, HÉRAL M,. Retention efficiency in oysterexposed to variable food quantity and quality. Ifremer, 1992, 8: 57–87
BAYNE B L, HAWKINS A J S, NAVARRO E. Feeding and digestion by the musselL. (Bivalvia: Mollusca) in mixtures of silt and algal cells at low concentrations. Journal of Experimental Marine Biology and Ecology, 1987, 111(1): 1–22
BAYNE B L, IGLESIAS J I P, HAWKINS A J S,. Feeding behaviour of the mussel,: Responses to variations in quantity and organic content of the seston. Journal of the Marine Biological Association of the United Kingdom, 1993, 73(4): 813–829
Cranford P J, Gordon Jr D C. The influence of dilute clay suspensions on sea scallop () feeding activity and tissue growth. Netherlands Journal of Sea Research, 1992, 30: 107–120
DAME R F, PRINS T C. Bivalve carrying capacity in coastal ecosystems. Aquatic Ecology, 1997, 31(4): 409–421
DAVIDS C. The influence of suspensions of microorganisms of different concentrations on the pumping and retention of food by the mussel (L.). Netherlands Journal of Sea Research, 1964, 2(2): 233–249
DONG S P, LIN F, JIANG W W,. Estimation of carrying capacity of Manila clam () in Jiaozhou Bay based on spatial and temporal distribution of chlorophyll. Progress in Fishery Sciences, 2020, 41(6): 100–107 [董世鹏, 蔺凡, 姜娓娓, 等. 基于叶绿素时空分布的胶州湾菲律宾蛤仔养殖容量评估. 渔业科学进展, 2020, 41(6): 100–107]
Eckman J E, Peterson C H, Cahalan J A. Effects of flow speed, turbulence, and orientation on growth of juvenile bay scallops(Say). Journal of Experimental Marine Biology and Ecology, 1989, 132(2): 123–140
FANG J G, KUANG S H, SUN H L,. Study on the carrying capacity of Sanggou Bay for the culture of scallop. Marine Fisheries Research, 1996, 17(2): 18–31 [方建光, 匡世焕, 孙慧玲, 等. 桑沟湾栉孔扇贝养殖容量的研究. 海洋水产研究, 1996, 17(2): 18–31]
HAWKINS A J S, NAVARRO E, IGLESIAS J I P. Comparative allometries of gut-passage time, gut content and metabolic faecal loss inand. Marine Biology, 1990, 105(2): 197–204
HILDRETH D I, MALLET A. The effect of suspension density on the retention of 5 µm diatoms by thegill. Biological Bulletin, 1980, 158(3): 316–323
HONKOOP P J C, BAYNE B L, DRENT J. Flexibility of size of gills and palps in the Sydney rock oyster(Gould, 1850) and the Pacific oyster(Thunberg, 1793). Journal of Experimental Marine Biology and Ecology, 2003, 282(1/2): 113–133
HOU X, GAO Y P, DU M R,. Temporal and spatial variation in phytoplankton community structure and their relationship with environmental factors in Sanggou Bay. Progress in Fishery Sciences, 2021, 42(2): 18–27 [侯兴, 高亚平, 杜美荣, 等. 桑沟湾浮游植物群落结构时空变化特征及影响因素. 渔业科学进展, 2021, 42(2): 18–27]
JØRGENSEN B C. On gill function in the musselL.. Ophelia, 1974, 13(1/2): 187–232
LI L, JIANG M, SHENG X Q,. Culture capacity ofin mud flats of Rudong, Jiangsu province. Marine Environmental Science, 2014, 33(5): 752–756 [李磊, 蒋玫, 沈新强, 等. 江苏如东滩涂文蛤养殖区养殖容量. 海洋环境科学, 2014, 33(5): 752–756]
LIN Y S, CAO W Q, LUO W X,. Experimental studies on filtration rates in four species of suspension feeding bivalves. Acta Oceanologica Sinica, 2003, 25(1): 86–92 [林元烧, 曹文清, 罗文新, 等. 几种主要养殖贝类滤水率的研究. 海洋学报, 2003, 25(1): 86–92]
LUCAS M I, NEWELL R C, SHUMWAY S E,. Particle clearance and yield in relation to bacterioplankton and suspended particulate availability in estuarine and open coast populations of the mussel. Marine Ecology Progress Series, 1987, 36(3): 215–224
MAURICE H. Evaluation of the carrying capacity of molluscan shellfish ecosystems. International seminar for research on planning and development aids to the shellfish industry. International Seminar in La Rochelle (France), 1985, 297–318
OFFICER C B, SMAYDA T J, MANN R. Benthic filter feeding: A natural eutrophication control. Marine Ecology Progress Series, 1982, 9(2): 203–210
PALMER R E, WILLIAMS L G. Effect of particle concentration on filtration efficiency of the bay scallopand the oyster. Ophelia, 1980, 19(2): 163–174
PAN L Q, FAN D P, MA S,. Influence of environmental factors on the filtration rate of. Journal of Fisheries of China, 2002, 26(3): 226–230 [潘鲁青, 范德朋, 马牲, 等. 环境因子对缢蛏滤水率的影响. 水产学报, 2002, 26(3): 226–230]
RIISGÅRD H. Efficiency of particle retention and filtration rate in 6 species of Northeast American bivalves. Marine Ecology Progress Series, 1988, 45(3): 217–223
SONG Q, FANG J G, LIU H,. Studies on the effects of suspended sediment on the feeding physiology of three suspension-feeding bivalves. Marine Fisheries Research, 2006, 27(4): 21–28 [宋强, 方建光, 刘慧, 等. 沉积再悬浮颗粒物对3种滤食性贝类摄食生理的影响. 海洋水产研究, 2006, 27(4): 21–28]
STENTON-DOZEY J M E, BROWN A C. Clearance and retention efficiency of natural suspended particles by the rock-pool bivalvein relation to tidal availability. Marine Ecology Progress Series, 1992, 82(2): 175–186
WALEN P R. The influence of current speed, body size and water temperature on the filtration rate of five species of bivalves. Journal of the Marine Biological Association of the United Kingdom, 1972, 52(2): 345–374
WILLIAMS L G. Mathematical analysis of the effects of particle retention efficiency on determination of filtration rate. Marine Biology, 1982, 66(2): 171–177
WILSON J H. Retention efficiency and pumping rate ofin suspensions of. Marine Ecology Progress Series, 1983, 12(1): 51–58
YANG S F, ZHANG L, YAN X Z. Estimation of carrying capacity of Manila clam () culture pond in Houhai reclamation area of Putian City, China. Environmental Science Survey, 2016, 35(6): 30–34 [杨淑芳, 张磊, 阎希柱. 莆田后海垦区菲律宾蛤仔养殖池养殖容量的估算. 环境科学导刊, 2016, 35(6): 30–34]
ZHANG J H, FANG J G, LIANG X M. Variations in retention efficiency of bivalves to different concentrations and organic content of suspended particles. Chinese Journal of Oceanology and Limnology, 2010, 28(1): 10–17
Effects of Particle Concentration on the Retention Efficiency of,and
XIE Bin1,2, LIN Zhihua2,3, HE Lin2①
(1. College of Marine Sciences, NingBo University, Ningbo, Zhejiang 315100, China; 2. Key Laboratory of Aquatic Germplasm Resource of Zhejiang Province, College of Biological and Environmental Sciences, Zhejiang Wanli University, Ningbo, Zhejiang 315100, China; 3. Ninghai Institute of Mariculture Seeds Industry, Zhejiang Wanli University, Ningbo, Zhejiang 315100, China)
In this study, the retention efficiency (RE) of,, andwere measured by adding sediment or unicellular algae to adjust the concentration of suspended particles. The results showed that in natural seawater, the RE of,, andreached maximum retention when the particle sizes were 8 μm, 6 μm, and 6 μm, which were 51.1%, 59.6%, and 62.6%, respectively. With an increase in particle number concentration, the minimum particle sizes for maximal retention inshifted to approximately 9 μm, 13 μm, and 14 μm in low (4.32×107cells/L), mid (5.27×107cells/L) and high (6.65×107cells/L) particle number concentration, respectively. The relative standardized RE decreased to 49.7%, 33.4%, and 26.2%. As in, the minimum particle sizes for maximal retention inshifted from 6 μm to 9 μm, 12 μm, and 14 μm with the change in particle number concentration, but the change in maximal retention was not significant. The change in maximal retention inwas not significant; however, the minimum particle sizes for maximal retention shifted to approximately 8 μm, 9 μm, and 10 μm, respectively. With the increase in particle concentration, the change in maximal retention and minimum particle size for maximal retention was not significant in three different particle concentration, low (5.7 mg/L), mid (11.8 mg/L) and high (23.3 mg/L), retention ofchanged significantly,60.7%, 27.6%, and 25.5%, respectively. The results indicate that the RE ofandwere more sensitive to particle concentration, while the RE ofwas more sensitive to organic content.
Suspended particle concentration; Retention efficiency;;;
HE Lin, E-mail: hlwithyou@qq.com
10.19663/j.issn2095-9869.20210310001
S968.31
A
2095-9869(2022)04-0190-09
*浙江省重点研发计划项目(2019C02054)、宁波市科技富民项目(2015C10008)和财政部和农业农村部: 国家现代农业产业技术体系共同资助[This work was supported by Focus on Research and Development Projects in Zhejiang Province (2019C02054), Ningbo Science and Technology and Enriching Project (2015C10008), China Agriculture Research System of MOF and MARA]. 谢 彬,E-mail: 1660654011@qq.com
何 琳,副教授,E-mail: hlwithyou@qq.com
2021-03-10,
2021-05-06
http://www.yykxjz.cn/
谢彬, 林志华, 何琳. 颗粒浓度对文蛤、硬壳蛤和菲律宾蛤仔保留效率的影响. 渔业科学进展, 2022, 43(4): 190–198
XIE B, LIN Z H, HE L. Effects of particle concentration on the retention efficiency of,and. Progress in Fishery Sciences, 2022, 43(4): 190–198
(编辑 马璀艳)
猜你喜欢
汽车实用技术(2022年15期)2022-08-19
西部交通科技(2021年1期)2021-11-09
意林·少年版(2021年18期)2021-09-29
红领巾·探索(2021年8期)2021-08-26
意林·少年版(2020年15期)2020-08-31
当代化工(2019年3期)2019-12-12
食品与生活(2019年8期)2019-10-30
智能建筑与工程机械(2019年1期)2019-09-10
食品与生活(2017年11期)2017-12-18
小学阅读指南·低年级版(2016年9期)2016-05-14
