
野生观赏植物黄金凤的繁殖生物学研究
2022-08-30 10:29曹孟会邵琳亚张颖刘婕左鹏飞汪琼黄海泉黄美娟
山东农业科学 2022年7期
曹孟会,邵琳亚,张颖,刘婕,左鹏飞,汪琼,黄海泉,黄美娟
(西南林业大学园林园艺学院/国家林业和草原局西南风景园林工程技术研究中心/云南省功能性花卉资源及产业化技术工程研究中心/西南林业大学园林园艺花卉研发中心,云南 昆明 650224)
野生观赏植物是处于野生或半野生状态、未在园林中普遍应用的一类具有观赏价值的植物,作为自然宝库中的精华,其潜在的种质资源利用价值难以估量。 生境多样化的野生观赏植物作为地球生态系统的重要组成部分,是能够反映区域环境演变和生物多样性层次的重要群体[1]。 我国是世界观赏植物原产地8 大起源分布中心,在世界生物多样性领域拥有突出的话语权,其中野生观赏植物约有113 科523 属,因此被亲切地赞誉为“世界园林之母”[2]。 野生观赏植物不断被利用于绿化和园林领域,在经济、社会、生态和文化等多方面产生了综合效益。 随着对野生观赏植物重要性的认知逐渐加强,人们对其相关研究也在不断深入。
凤仙花属(Impatiens)和水角属(Hydrocera)共同构成了凤仙花科。 1753年建立的凤仙花属作为被子植物中少数几个物种量超过1000 的世界性大属[3],主要分布于欧亚大陆、非洲的热带与亚热带高海拔(>500 m)高原及山地地区,少数生活于欧洲、北美和东亚的温带地区。 中国是世界凤仙花5 大起源分布中心之一,约有280 种凤仙花属植物,除I. balsamina、I. walleriana、I. usambarensis外均为野生种[4];其凤仙花属植物主体分布于第一阶梯的青藏高原,而位于第三阶梯的广大平原地区种类相对较少。 因此,中国凤仙花属植物在地理上的分布具有明显的区域性,即各个省、直辖市、自治区均有分布,但大多数是西南狭域分布的特有种,其中云南省的种类最多;石灰岩地区的特有性则更为突出,堪称“一山一种、一洞一种、一弄一种”[5]。 中国特有野生观赏植物黄金凤(Impatiens siculifer)又被称为广西水指甲,具有两侧对称状黄色花冠,花形奇特、花色明亮、花香淡雅,广泛分布于云南、广西、贵州等省、自治区,主要生长在海拔800 ~2500 m 的草丛、山谷及潮湿密林中[6],具有良好的开发和利用价值。
植物繁殖生物学是以植物生殖这一生命过程为核心,将生态系统中植物和环境之间的相互作用与相互适应关系有机地结合在一起[7],深刻揭示了生态系统中能量、基因的信息传递规律,是一门研究植物繁殖行为特征和过程以及探索自然规律与植物相互关系的学科,其中,开花物候学、繁育系统、传粉生物学、植物孢粉学、传粉策略5 个方面[8]从不同角度解释了繁殖生物学的具体内涵意义。 近年来关于凤仙花属植物繁育系统、传粉策略等相关繁殖生物学的研究逐渐增加。 田建平[9]于2004年首次提出花蜜抢窃者的作用:可能与植物居群花期的提早结束有关;首次探究匍匐凤仙花与传粉昆虫相互适应和协同进化问题;同时认为对特殊生存环境的选择、种子萌发率低及生态位缺失等因素导致了匍匐凤仙花只在一定区域存在。 2009年肖乐希[10]研究了华凤仙、瑶山凤仙花、山地凤仙花和湖南凤仙花的花部特征与繁育系统的关系,证实花部结构在繁殖中与其长短和相对位置有关,对繁殖成功的影响存在区别,肯定了翼瓣对于繁殖成功的重要性。 2011年毛志斌等[11]研究指出侧穗凤仙花能通过特别的花部器官结构来避免自交。 2014年钟云芳[12]提出人工授粉能明显提高结实率,自然结实率在高海拔地区相对较低,存在较强的生殖限制;同年,还通过调查发现海南凤仙花的授粉昆虫主要为黄黑无垫蜂和绿条无垫蜂[13]。 2014年李艳红等[14]研究发现盛花期红雉凤仙花大量自交导致的花粉质量限制而不是数量限制可能是其结籽率低的主要原因之一。 2021年钟楠蝶等[15]试验发现异色纱网袋套蜜距的花朵被传粉者访问的频率差异显著,纱网袋颜色对传粉者的停留时间无显著影响,观测时间段对目标花朵被访问频率和单花停留时间的影响均显著,为隔离盗蜜者以及研究盗蜜对植物与传粉者互利共生关系的影响提供了重要的方法参考。
2004年14 种凤仙花属植物作为珍惜濒危植物被列入IUCN 世界自然保护联盟濒危物种红色名录,2020年凤仙花属植物被新增选为第七批中华人民共和国植物新品种保护名录(林草部分),凤仙花属植物的种质资源保护工作越来越受到人们的关注。 开展黄金凤的繁殖生物学研究,对于探究其有性繁殖过程和系统进化的演化趋势具有积极意义,同时可为凤仙花属野生观赏种黄金凤种质资源的育种扩繁、花期调控、引种驯化等保护与开发工作提供理论指导与科学依据。
1 材料与方法
1.1 试验材料和地点
试验材料为野外采集的黄金凤种子栽培养护后形成的成年植株,播种时间为2020年6月21日,种子采集地为云南省红河州屏边县大围山(103°41′47″E,22°56′37″N)。 观测地点位于昆明市西南林业大学树木园(102°46′5″E,25°3′58″N)。
1.2 研究方法
1.2.1 开花物候学 于花芽萌动期从不同黄金凤植株上选取30 朵花芽进行标记,每0.50 d 统计1 次发育状况,记录不同发育阶段的持续时间和单花发育进程。
1.2.2 植物孢粉学 (1)花粉生活力和柱头可授性:采用TTC 染色法[16]测定花粉生活力。 采用联苯胺-过氧化氢法[17]测定柱头可授性。
(2)花蜜量和含糖量:使用微量进样器(10 μL)深入花距中(图1)抽取花蜜,记录花蜜量。将花蜜注入1.50 mL EP 管中,使用手持折光仪(0~80%)[18]测量含糖量。
1.2.3 繁育系统 (1)花粉-胚珠比和杂交指数:在室内用镊子夹取花药,置于1.50 mL 离心管中,用50%酒精制成花粉悬浮液;用滴管吸打混匀后,利用血球计数板统计花粉粒数目。 解剖子房统计胚珠数,计算单花花粉数与胚珠数之比,即为P/O 值。 按照Cruden[19]和Dafni 等[20]的方法测定杂交指数,即为OCI。
(2)繁育类型:共分为5 种[19,20],分别为闭花受精型(P/O =2.70 ~5.40,OCI =0)、专性自交型(P/O=18.10 ~39.00,OCI =1)、兼性自交型(P/O =31.90~396.00,OCI =2)、兼性异交型(P/O =244.70~588.70,OCI =3)、专性异交型(P/O =608.00~19525.00,OCI≥4)。
1.2.4 传粉生物学 (1)授粉试验:在花芽萌动期标定30 朵花芽,于现蕾期对其进行6 种授粉处理[20],统计结实率。
(2)传粉通道:使用游标卡尺(mm)分别测量雄蕊到翼瓣凸起的距离(通道长度)、2 朵翼瓣中段直线距离(通道宽度)以及翼瓣到唇瓣檐部与距部接壤处的距离(通道高度),见图1。

图1 黄金凤的花部示意图和传粉通道
1.2.5 传粉策略 在盛花期标定30 朵花冠,于9—18 时(单位时间为1 h)观察并统计传粉虫媒种类及数量,记录其访花时间、访花方式与访花频率,持续进行7 d。
1.3 数据统计分析
利用SPSS 22 软件采用单因素方差分析法进行数据统计分析。
2 结果与分析
2.1 开花物候学
通过对黄金凤物候期的观察发现,其花期在7—9月,开花进程分为花芽萌发期、花蕾期、初花期、盛花期、末花期、结实期6 个时段,共持续37.00~55.00 d;单花期持续5.00~10.50 d(表1 和图2),规定开花进程各时段的发育程度为0.25 ~10.00。在花芽萌发期,黄金凤的花芽由原来的近似三角形逐渐膨大、弯曲并最终演变为花蕾;花蕾期花蕾继续膨大,花萼微张,花距弯曲程度加剧;初花期花药未发生开裂;盛花期花药开裂并向外散播花粉;末花期雄蕊群脱落,柱头开始出现;结实期果实形成,体积逐渐膨大,最终爆裂散播种子。
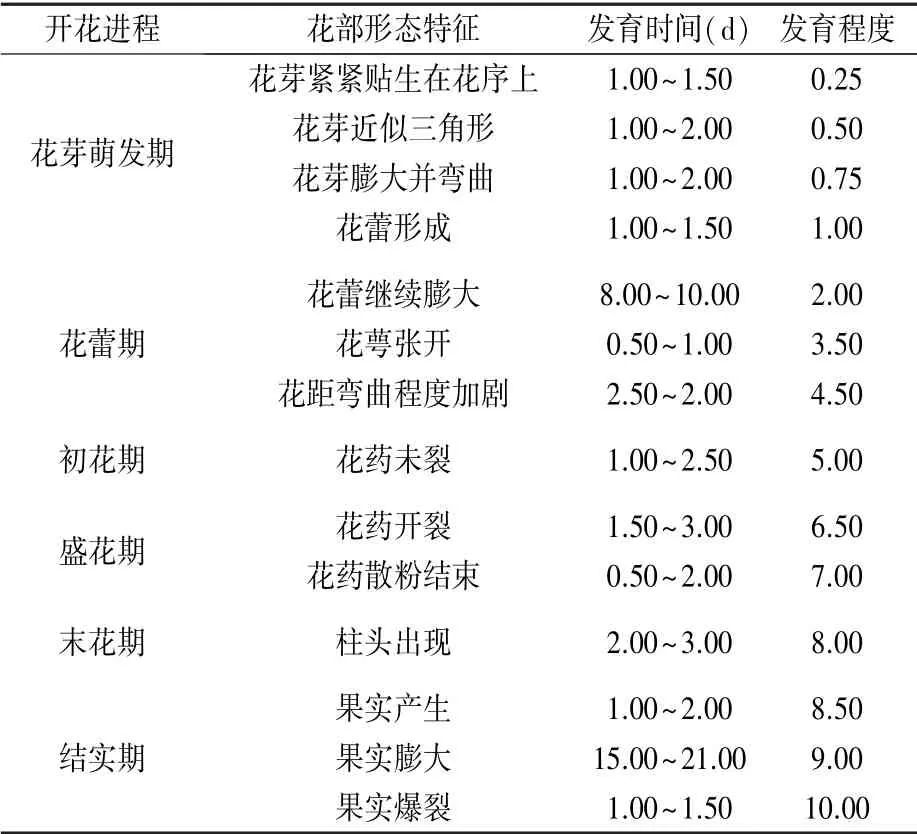
表1 黄金凤的开花物候学

图2 黄金凤的开花进程
2.2 植物孢粉学
2.2.1 花粉生活力和柱头可授性 花蕾期黄金凤的花粉已具备较高生活力,柱头具备较弱可授性;初花期花粉生活力最高(>90%),柱头具备较强可授性,部分花朵的柱头已经开始响应成花诱导信号,在后续开放过程中花粉生活力随花期推进递减;盛花期柱头进入最强可授期;末花期花粉生活力最低(<5%),柱头失去可授性(表2)。 花粉生活力与柱头可授性的结果总体均呈现倒“U”型态势,但最高值所处时期不同,可据此推断黄金凤属于雄蕊先熟植物。

表2 黄金凤的花粉生活力和柱头可授性
2.2.2 花蜜量和含糖量 黄金凤的单花花蜜量为(0.52±0.08) μL,花蜜含糖量为33.50%。 其唇瓣囊部深度相比其他种类凤仙花较长,在符合自身繁殖策略的同时可满足昆虫的传粉动机。
2.3 繁育系统
2.3.1 花粉-胚珠比和杂交指数 于盛花期对30朵黄金凤进行测定,测得单花花粉量为5189.60粒,单花胚珠数为7.50,由此可知单花花粉-胚珠比为691.95。
2.3.2 繁育类型 黄金凤花朵直径平均值为25.17 mm,记为3;柱头在开花前一天就具有可授性,记为1;花药合生在子房上部,记为0。 综上可知黄金凤的P/O 为691.95、OCI 为4,表明其为专性异交型,特征为部分自交亲和,需要传粉者。
2.4 传粉生物学
2.4.1 授粉试验 黄金凤的蒴果为条形,顶端钝。 自然状态下的结实率小于人工授粉的结实率,且授粉方式不同其结实率差异较大(表3),表明黄金凤在自然状态下的有性繁殖可能受到花粉限制以及缺乏有效的传粉虫媒,其繁育系统以异交型为主,不存在无融合生殖。

表3 黄金凤人工授粉试验结果
2.4.2 传粉通道 黄金凤作为雌雄异熟的两性花,雄蕊脱落后柱头才会外显,其通道长度为(13.10±0.12) mm、宽度为(6.82±0.25) mm、高度为(4.88±0.36) mm,适宜中小体型昆虫顺利通行。
2.5 传粉策略
黄金凤的传粉虫媒主要有6 类:膜翅目蜂类、膜翅目蚁类、鳞翅目蝶类、鳞翅目蛾类、双翅目蝇类、鞘翅目甲虫类,其中,黄熊蜂(Bombus flavescens)访花次数最多,三条熊蜂(Bombus trifasciatus)次之,访花时间主要分布在9—18 时(图3)。膜翅目蜂类访花时停靠在旗瓣上,然后伸出口器来吸取花蜜,具有访花速度快、访花频率高等特性,停留时间为2~6 s,使用手持放大镜可看到其腹部的绒毛均匀分布着黄金凤的花粉。 膜翅目蚁类、鳞翅目蝶类和蛾类经常在不同种类花朵间游走,停留时间约10 ~20 s。 双翅目蝇类借助长吻深入唇瓣内部吸食花蜜,对黄金凤的传粉作用较小,停留不超过8 s。 体型较大的鞘翅目甲虫类同样通过游走来推动传粉,一般停留5~7 s。
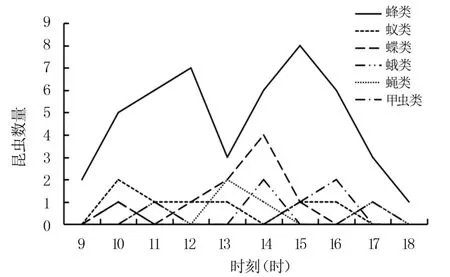
图3 传粉昆虫的访花情况
3 讨论与结论
3.1 花部构造对传粉策略的影响
植物凭借整体花器官构造和开放式样吸引虫媒[21]进行花粉扩散来主导生殖场景,为虫媒传粉途径的产生创造了有力条件。 黄金凤为总状花序,整体美化效果优,增强了对传粉虫媒的吸引力。 虫媒花的花冠通常呈左右对称状、花色明亮[22]。 黄金凤具有左右对称状黄色花,符合虫媒花标准,其传粉通道长度为(13.10±0.12) mm、宽度为(6.82±0.25) mm、高度为(4.88±0.36) mm,适宜中小体型昆虫顺利通行。 黄金凤的有效传粉昆虫为膜翅目蜂类的黄熊蜂和三条熊蜂,其在进入花朵内部后挤压内壁所产生的反作用力增强了与花药接触的成功率,有利于进行异花传粉。 黄金凤的花距较长,可能会增大虫媒获取花蜜的困难程度,从而形成一种特殊协同进化关系,推动黄金凤的花部构造不断演化,以与虫媒的传粉方式相协调。
3.2 黄金凤的繁殖策略
黄金凤的花果期在7 ~9月,分为花芽萌发期、花蕾期、初花期、盛花期、末花期、结实期6 个时期,初花期花粉生活力最高(92.21%),盛花期柱头进入最强可授期,为持续开花式样,同时其单花花蜜量为(0.52±0.08) μL,含糖量为33.50%,符合自身繁殖策略,满足昆虫的传粉动机。 雄蕊先熟是促进异交的重要繁殖机制。 黄金凤作为雄蕊先熟植物,自然授粉途径为异花授粉,两者结合共同增强其子代育性及种群的传播范围,提高了繁育成功率,有利于系统演化的不断发展。 人工授粉试验中黄金凤没有产生无融合生殖现象,其繁育系统特征为部分自交亲和,需要传粉者;而同属植物侧穗凤仙花则通过避免自花授粉的机制加大虫媒授粉的概率[9],水金凤繁育类型则为兼性异交[23]。 说明自交与异交都是凤仙花属植物的有效繁殖机制。
实际观测发现黄金凤的野外种群规模较小,推测可能由以下几种因素所导致:易受到花粉限制;缺乏有效的传粉媒介;种子较小,种荚在受到外部作用力时会迅速爆炸裂开,进行远距离传播比较困难。 因此,黄金凤的保护与开发工作要着重加强对传粉者的保护,解除传粉者限制,探明生殖发育规律,并将分株、扦插、组织培养等无性繁殖方式作为有性繁殖的补偿机制,制定出一套繁殖对策以更好地促进黄金凤遗传多样性与有效种群面积的扩展。
猜你喜欢
安徽农业科学(2022年18期)2022-10-13
农产品市场周刊(2022年4期)2022-05-21
农产品市场周刊(2022年4期)2022-03-23
科学之友(2021年12期)2021-12-23
湖北农业科学(2019年20期)2019-12-11
安徽农业科学(2018年27期)2018-05-14
大科技·百科新说(2018年3期)2018-04-11
科学种养(2017年11期)2017-11-28
山西果树(2015年3期)2015-05-29
