
毛蕊花糖苷抑制2型猪链球菌的溶血素蛋白活性而降低其小鼠致病性
2022-08-27 05:12詹佳飞夏介英刘洋露袁明铭王永金鄢良春
浙江农业学报 2022年8期
詹佳飞, 徐 魁, 张 磊, 夏介英, 洪 杨, 董 涵, 刘洋露, 周 静, 袁明铭, 王永金, 鄢良春,*
(1.四川省中医药科学院 实验动物中心,四川 成都 610041; 2.四川农业大学 动物医学院,四川 成都 611130)
猪链球菌(,)是一种呈世界范围分布的革兰氏阳性菌,人和牲畜(尤其是猪)感染后通常会引发脑膜炎、败血症及关节炎等临床症状,给人畜的生命健康带来严重威胁,同时给养猪业造成了巨大的经济损失。根据荚膜抗原的差异,猪链球菌可被分为35种血清型(1~34及1/2),其中属2型猪链球菌(serotype 2, SS2)的毒力最强,且流行范围最广。目前由于长期使用青霉素、阿莫西林、氨苄西林等抗生素,猪链球菌的耐药性日益加重,因此,寻求更加安全有效的办法抵抗猪链球菌感染已是当务之急。
猪链球菌在感染宿主过程中可分泌多种毒力因子,研究表明,毒力因子对猪链球菌的生长虽无必要,但却与猪链球菌的致病性强弱密切相关。溶血素(suilysin, SLY)是猪链球菌一种重要的毒力相关因子,被认为是猪链球菌的毒力标志物。SLY与细菌的溶血活性有重要关系,对人、猪、鸡及鼠等动物的红细胞均表现出很强的溶血活性,可溶解红细胞释放血红蛋白。SLY属于胆固醇依赖性细胞裂解素家族(cholesterol dependent cytolysins, CDCs),由4个结构域组成。当SLY被细菌释放后,SLY的第4结构域最先特异性识别并结合宿主细胞膜上的胆固醇。与细胞膜结合后的SLY单体通过构象改变发生寡聚形成成孔前体,随后第3结构域的两个β-发夹结构(TMH1、TMH2)插入细胞膜形成β-桶样跨膜结构,从而在细胞膜上形成亲水性孔道,最终引起细胞裂解直至死亡。Takeuchi等认为在感染过程中SLY高分泌的猪链球菌菌株比SLY低分泌的菌株具有更强的毒性及侵袭力。将基因缺失后,缺失株的毒力显著下降,几乎不造成感染小鼠死亡。此外,在转录水平通过上调基因可增强猪链球菌的毒力。因此,靶向溶血素SLY的抗毒力因子策略为治疗猪链球菌感染提供了一种新的思路。
毛蕊花糖苷(verbascoside,VBS)又被称为类叶升麻苷,是一种苯乙醇苷类化合物,主要存在于连翘、肉苁蓉、洋丁香、车前草及绿豆等多种植物中,单体经机体摄入后能快速吸收发挥生理作用,例如抗炎镇痛、抗氧化等。目前已有研究表明,连翘中的VBS可对金黄色葡萄球菌(,)产生中等强度的抑制作用。因此,本研究拟将毛蕊花糖苷作用于2型猪链球菌,探讨该药物对2型猪链球菌致病力的影响及其机制研究,为临床上治疗2型猪链球菌病提供一个新的研究方向。
1 材料与方法
1.1 材料
试验所需2型猪链球菌菌株SC21由四川省中医药科学院实验动物中心提供;胰酪大豆胨液体培养基(TSB)、小牛血清、二甲基亚砜(DMSO)及鸡红细胞均购自索莱宝(北京)有限公司;鼠源GFP抗体购自武汉爱博泰克生物科技有限公司;HRP标记的羊抗鼠IgG抗体购自武汉博士德有限公司;毛蕊花糖苷(纯度>98%)购自成都普思生物科技股份有限公司,利用DMSO将毛蕊花糖苷稀释为20 480 μg·mL,于-20 ℃环境下避光储存备用;雌性C57 BL/6J小鼠(7周龄)购自北京维通利华实验动物技术有限公司。
1.2 方法
1.2.1 测定最低抑菌浓度(MIC)试验
将冻存的2型猪链球菌菌株SC21按1∶50接种于新鲜的TSB培养基(添加5%小牛血清)中,在摇床内37 ℃振荡(220 r·min)培养10 h,于4 ℃环境下保存备用。
毛蕊花糖苷对2型猪链球菌MIC的测定以“美国临床实验室标准化委员会(NCCLS)抗菌药物敏感性实验执行标准”为依据。具体方法如下,将毛蕊花糖苷储存液按0.5~2 048 μg·mL的浓度梯度稀释,并依次加到等量分装的2型猪链球菌中使菌液浓度均为5×10CFU·mL。另外分别设置阳性对照和阴性对照,其中阳性对照中不加入药物只加等量细菌重悬液,阴性对照中只加入高压灭菌的TSB培养液(添加5%小牛血清)。将处理好的培养物放入37 ℃恒温培养箱内,每隔12 h观察一次细菌生长情况。以培养物呈现透明状态,没有肉眼可见细菌生长为标准,该药物浓度即为最低抑菌浓度。
1.2.2 2型猪链球菌生长曲线的绘制
将2型猪链球菌菌液接种到TSB培养液(添加5%小牛血清)中振荡培养至菌液为0.3。将菌液等量分装并依次加入不同剂量的毛蕊花糖苷储存液,使培养物中药物终浓度分别达到0、0.5、1、2、4、8、16、32及64 μg·mL。将培养物放入摇床内继续振荡6 h并每隔30 min测一次菌液吸光度。
1.2.3 毛蕊花糖苷对2型猪链球菌溶血活性的测定
在约为0.3的2型链球菌菌液中加入不同剂量的毛蕊花糖苷使药物浓度梯度稀释(0、0.5、1、2、4、8、16、32、64 μg·mL)。将培养物振荡培养12 h后离心(12 000×,2 min)并收集培养物上清。将25 μL鸡红细胞(4%)、100 μL上述培养物上清及875 μL PBS缓冲液轻柔混匀并放入37 ℃培养箱内孵育1 h。将孵育后的混合物低速离心(3 000×,5 min)获取混合物上清并测定其吸光度。另外利用1% Triton X-100和PBS缓冲液分别作为阳性对照和阴性对照。
1.2.4 免疫印迹试验
参照1.2.3节试验步骤,收集2型猪链球菌与毛蕊花糖苷(0~64 μg·mL)共同培养体系的上清。参照之前研究的操作步骤,在收集的上清中分别加入等量纯化的GFP蛋白(本实验室前期已制备)作为内参对照。在20 μL上清中加入4 μL 6×上样缓冲液混匀并加热煮沸8 min,离心(12 000×,2 min)取其上清进行12% SDS-PAGE凝胶电泳。将凝胶转移至硝酸纤维素膜上,5%脱脂牛奶封闭2 h后,分别加入1∶1 000的鼠源SLY一抗(本实验室前期已制备)及1∶2 000的鼠源GFP一抗4 ℃孵育过夜。经洗涤后,加入1∶2 000以HRP标记的羊抗鼠IgG作为二抗常温孵育2 h,最后使用化学发光成像分析系统进行显色观察。
1.2.5 分子模拟对接
在PDB在线软件() 中下载SLY蛋白(代码: 3HVN)三维结构,PubChem在线软件(https://pubchem.ncbi.nlm.nih.gov/)可下载毛蕊花糖苷小分子(代码: 5281800)的二维和三维结构。利用AutoDock Vina 1.5.6软件对SLY及VBS进行半柔性对接。在对接过程中毛蕊花糖苷所有扭转键均可自由旋转,SLY蛋白则保持刚性结构。
1.2.6 SLY蛋白的表达及纯化
以除去信号肽及终止密码子的基因序列(GenBank: AY341263.1)为模板设计引物(上游5′-GATTCCAAACAAGATATTAATCA-3′,酶切位点为H Ⅰ;下游5′-TACTCTATCACCTCATCCGCAT-3′,酶切位点为d Ⅲ)。从cDNA中扩增出序列,将扩增产物连接到pCold-I质粒上并转入大肠埃希菌BL21中,37 ℃条件下利用1 mmol·LIPTG诱导表达8 h,离心(12 000×,2 min)收集菌体沉淀,用超声破碎仪超声裂解重组菌。利用Ni亲和层析的方法对重组蛋白进行纯化,且纯化后的蛋白质浓度用BCA蛋白试剂盒进行测定。
1.2.7 毛蕊花糖苷对SLY蛋白溶血活性的测定
将100 μL纯化后的SLY重组蛋白与毛蕊花糖苷均匀混合使药物终浓度梯度稀释(0、0.5、1、2、4、8、16、32、64 μg·mL),将培养物置于37 ℃培养箱内孵育30 min。之后将25 μL的鸡红细胞(4%)、100 μL上述培养物及875 μL PBS缓冲液混匀继续在37 ℃培养箱孵育1 h后,低速离心(3 000×,5 min)获取上清并测定其。此外利用1% Triton X-100和PBS缓冲液分别作为阳性对照和阴性对照。
1.2.8 小鼠体内感染试验
将2型猪链球菌菌液培养至约为0.8时低速离心(3 000×,5 min)收集菌体,利用无菌PBS反复清洗菌体3次并重悬菌体使其浓度保持于5.6×10CFU·mL。将雌性C57 BL/6J 小鼠随机分为3组,每组10只,分别为SC21+VBS组、SC21+DMSO组、PBS+VBS组。对实验组小鼠腹腔注射500 μL上述细菌重悬液,对照组小鼠腹腔注射500 μL PBS。猪链球菌感染小鼠2 h后,皮下注射100 mg·kg毛蕊花糖苷,并隔8 h再次给药,连续观察72 h并每隔12 h统计一次小鼠死亡率。
1.3 数据分析
所有试验均进行了3次重复。软件GraphPad Prism 8.0被用来进行实验数据分析,平均值±标准差表示最终结果,Student’s-test 方法分析组间数值的统计学差异。* 表示差异显著(<0.05),**表示差异极显著(<0.01)。
2 结果与分析
2.1 毛蕊花糖苷对2型猪链球菌生长活性的影响
试验结果显示,毛蕊花糖苷对2型猪链球菌的最低抑菌浓度高达1 024 μg·mL。此外,当不同浓度的毛蕊花糖苷(0~64 μg·mL)作用2型猪链球菌6 h后,菌液近乎相同,由此可见在最低抑菌浓度范围内毛蕊花糖苷并不会抑制2型猪链球菌的生长活性(图1)。
2.2 毛蕊花糖苷对2型猪链球菌溶血活性的抑制作用
如图2所示,当没有毛蕊花糖苷作用时猪链球菌菌液上清的溶血活性为77.97%。随着药物浓度的增加,上清溶血活性呈现逐渐下降的趋势,其中当药物浓度为64 μg·mL时,上清溶血活性已降至5.24%。试验结果表明,较低浓度的毛蕊花糖苷即可抑制2型猪链球菌的溶血活性,且其溶血活性与药物的作用浓度呈负相关关系。
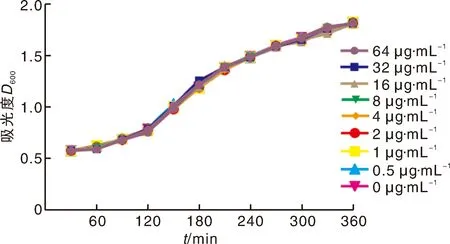
图1 在不同浓度毛蕊花糖苷作用下2型猪链球菌的生长曲线Fig.1 Growth curves of SS2 treated with different concentrations of VBS
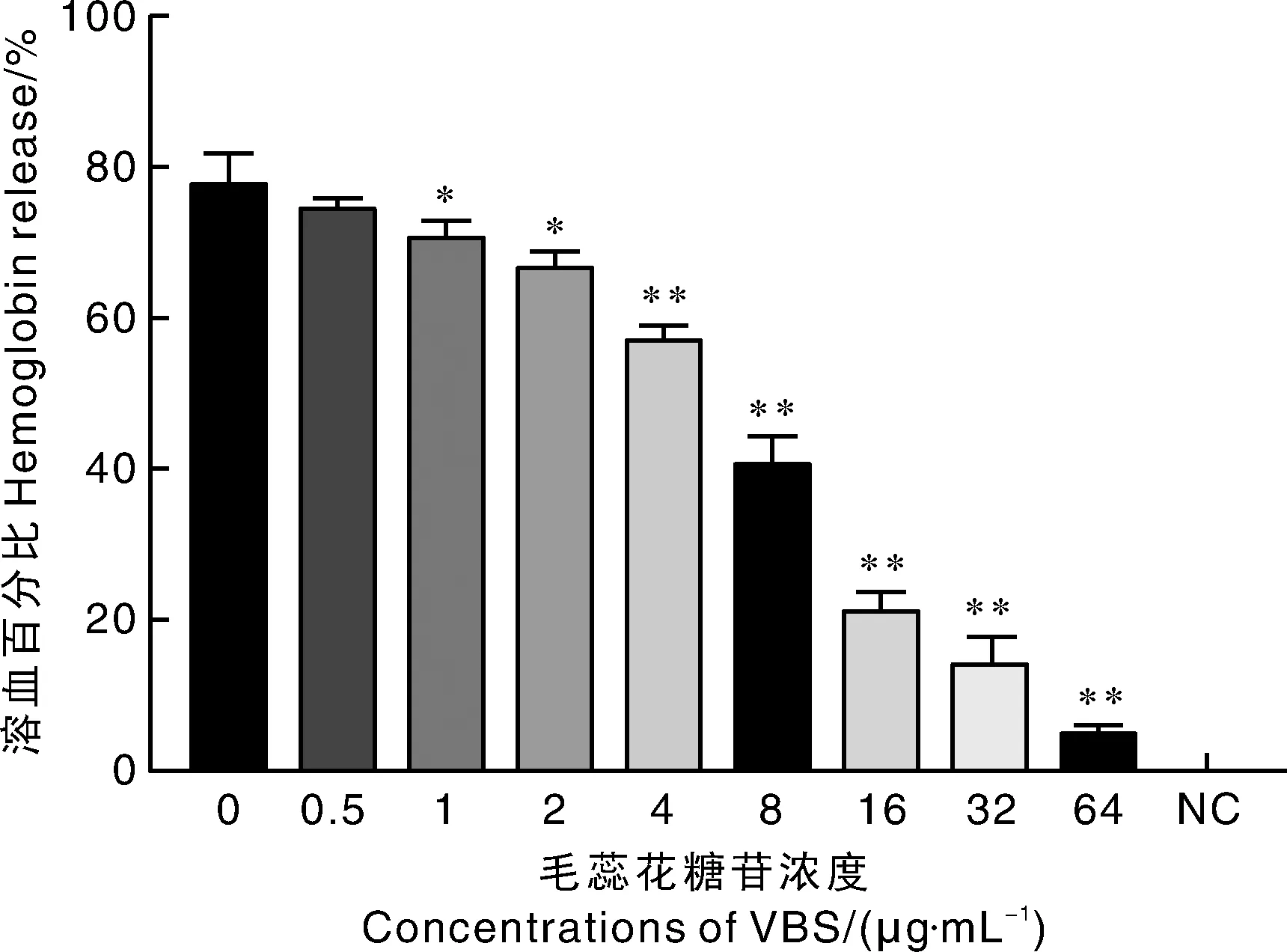
NC, 阴性对照。*表示与未添加毛蕊花糖苷组差异显著 (P<0.05),**表示与未添加毛蕊花糖苷组差异极显著 (P<0.01)。下同。NC, Negative control. * meaned significant difference compared with VBS-free group (P<0.05), ** meaned extremely significant difference compared with VBS-free group (P<0.01). The same as below.图2 毛蕊花糖苷与2型猪链球菌共同培养体系中上清的溶血活性Fig.2 Hemolytic activity in supernatant of VBS and SS2 co-culture system
2.3 毛蕊花糖苷对2型猪链球菌分泌SLY蛋白的影响
免疫印迹结果表明,经过不同浓度的毛蕊花糖苷处理后,猪链球菌菌液上清中的SLY蛋白分泌量没有明显区别,可见毛蕊花糖苷对2型猪链球菌分泌SLY蛋白没有影响(图3)。
2.4 毛蕊花糖苷与SLY蛋白相互结合分析
经分子模拟对接试验预测,毛蕊花糖苷主要结合于SLY蛋白的第4结构域,其中药物苯环中的羟基可与SLY蛋白2个氨基酸残基SER-388及GLN-441形成3条氢键相互作用,距离分别为2.1、2.4和2.6 nm,该结果表明,毛蕊花糖苷与SLY蛋白具有较高的结构亲和力(图4)。

图3 毛蕊花糖苷与2型猪链球菌共同培养体系中上清的SLY分泌量Fig.3 Expression of SLY in supernatant of VBS and SS2 co-culture system
2.5 SLY重组蛋白的表达与纯化
经SDS-PAGE电泳分析,在C端添加了6个组氨酸标签的SLY蛋白被成功原核表达。经纯化后SLY重组蛋白显示为单一条带,大小约为50 ku,符合预期蛋白大小。采用BCA蛋白浓度测定试剂盒测定重组蛋白为1 μg·μL(图5)。
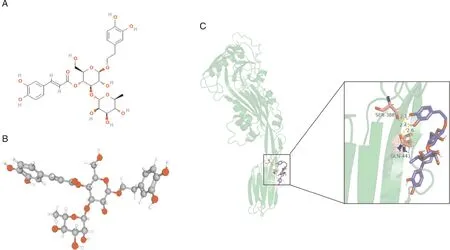
A,毛蕊花糖苷二维化学结构;B,毛蕊花糖苷三维化学结构;C, 毛蕊花糖苷的羟基与SLY残基结合细节。A, 2D chemical structure of VBS; B, 3D chemical structure of VBS; C, Details of binding between the hydroxyl group of VBS and the residues of SLY.图4 毛蕊花糖苷与SLY蛋白的分子对接构象Fig.4 The molecular docking conformation of VBS with SLY
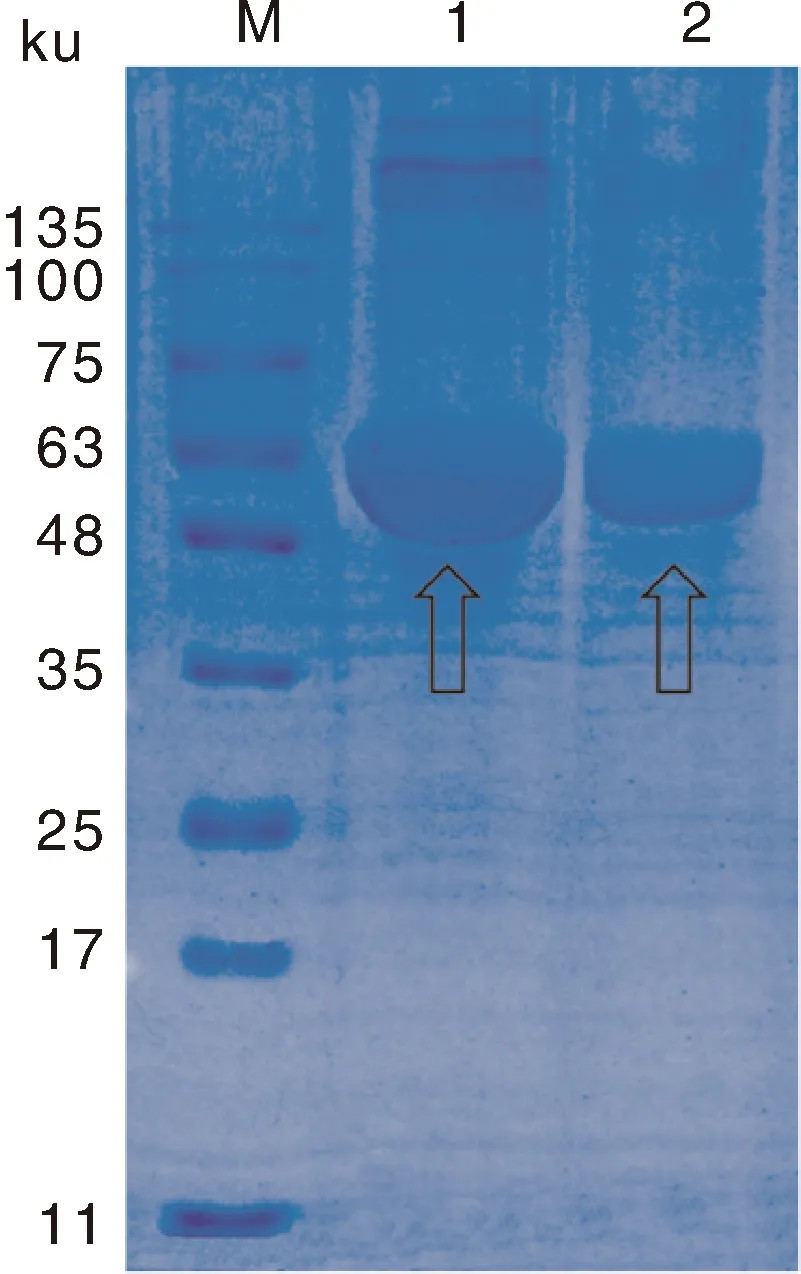
M, 蛋白质分子量标准;1,加IPTG诱导大肠埃希菌BL21表达SLY重组蛋白;2,纯化后的SLY重组蛋白。M, Protein molecular weight marker; 1, Lysate of IPTG-induced E.coil BL21 expressing recombinant SLY; 2, Purified recombinant SLY.图5 SLY重组蛋白的表达及纯化Fig.5 Expression and purification of recombinant SLY
2.6 毛蕊花糖苷对SLY蛋白溶血活性的抑制作用
将不同浓度的毛蕊花糖苷直接作用纯化的SLY蛋白后,结果显示,SLY蛋白介导的溶血活性随药物浓度的增加呈现出逐渐下降的趋势,呈现明显的浓度依赖性。其中16 μg·mL的毛蕊花糖苷便可将SLY溶血活性降至41.31%,显著地抑制了SLY蛋白活性(图6)。
2.7 毛蕊花糖苷对2型猪链球菌致病力的影响
为了进一步探究毛蕊花糖苷是否具有降低2型猪链球菌毒力的功效,本研究将猪链球菌作用于小鼠。结果显示,SS2+DMSO组小鼠经猪链球菌感染后在72 h内全部死亡,但是感染后给予毛蕊花糖苷的SS2+VBS组小鼠死亡率下降至70%,且明显延缓了小鼠的死亡时间,提高了小鼠的存活率,这表明毛蕊花糖苷可有效降低猪链球菌对小鼠的致病性(图7)。
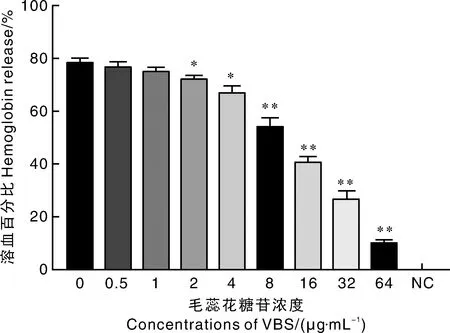
图6 毛蕊花糖苷对纯化后SLY溶血活性的作用Fig.6 Effect of VBS on the hemolytic activity of purified SLY
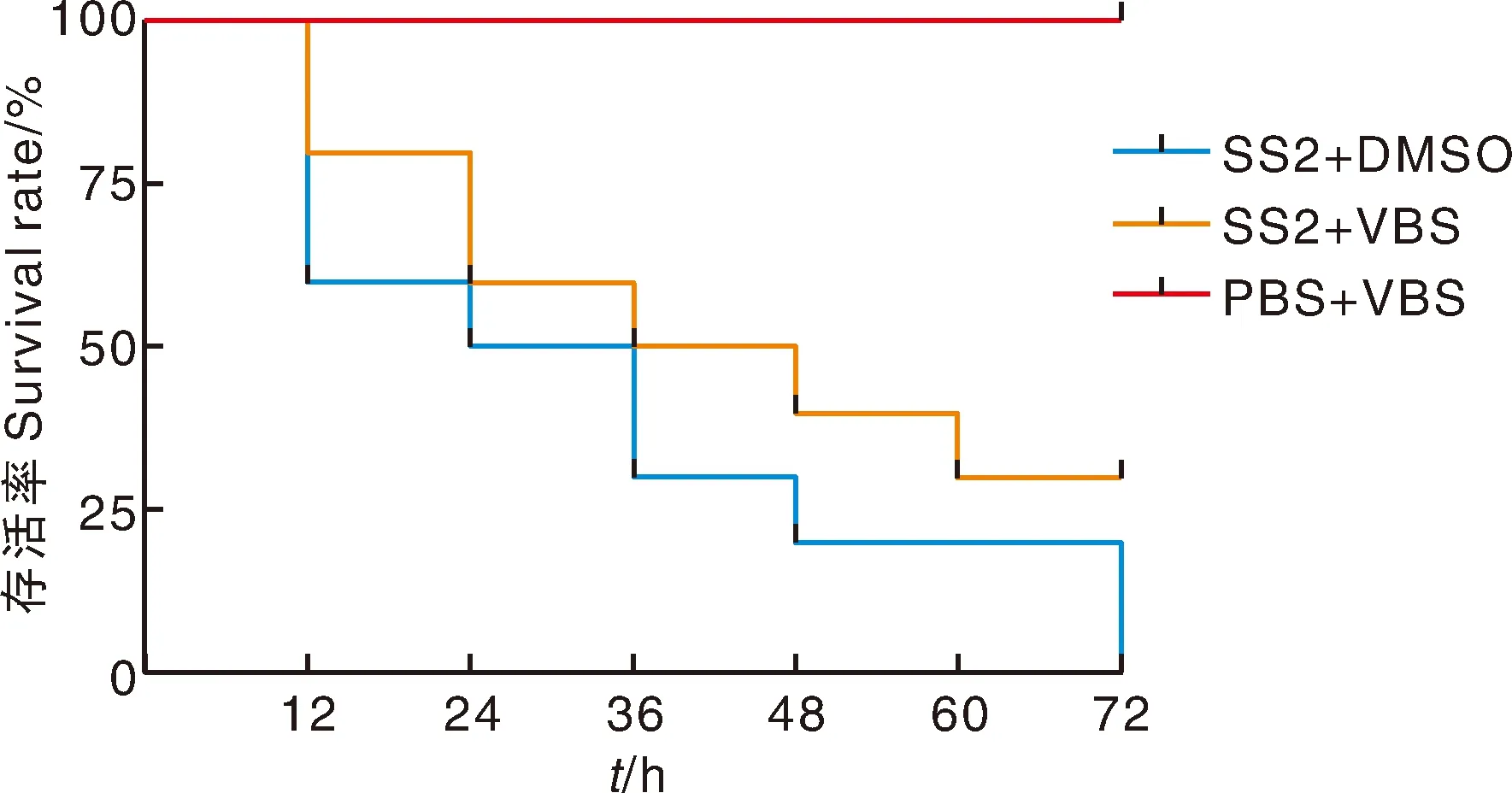
图7 毛蕊花糖苷对2型猪链球菌感染小鼠存活率的影响Fig.7 Effect of VBS on the survival rate of mice infected with SS2
3 讨论
目前临床上主要采用抗生素对猪链球菌进行治疗,抗生素主要以致病菌生存所必需的成分(DNA、蛋白质和细胞膜等)为靶标来达到杀菌的作用。虽然该方法可以有效清除病原菌,但是长期使用甚至滥用抗生素会给予细菌更多的生存选择压力进而使细菌耐药性加重。研究表明,病原菌的毒力因子是其致病环节中的重要一员,而与细菌毒力相关的基因或蛋白通常并不是致病菌生存所必需的。因此,抗毒力策略在有效降低致病菌对机体感染的基础上,对细菌生存选择压力较小,细菌不易产生耐药性,且该策略能给机体更多的时间通过自身免疫系统阻碍细菌感染。SLY作为2型猪链球菌重要的毒力相关因子,可通过降低其活性抵抗猪链球菌的感染。因此,SLY成为抑制2型猪链球菌感染的理想靶点。
在我国,针对细菌性传染病,中药及其化合物有着独特的治疗体系及显著的治疗效果。通过临床应用发现,和传统抗菌药物相比,中药及其化合物使用更加安全,对病原菌的生存压力更小,不易诱导耐药性的产生和发展。在本研究中,我们将单体化合物——毛蕊花糖苷作用于2型猪链球菌,发现药物在几乎不影响猪链球菌生长活性的基础上,可有效抑制其溶血活性。研究表明,猪链球菌的溶血活性主要通过SLY介导,故毛蕊花糖苷作用2型猪链球菌导致该病原菌溶血活性下降可能是其抑制了猪链球菌分泌SLY的过程,也可能是药物通过直接作用SLY蛋白导致其失去活性。为验证上述假设,本研究首先通过免疫印迹试验证实毛蕊花糖苷对猪链球菌的SLY分泌量几乎没有影响。其次,我们采用了分子对接的方式探索毛蕊花糖苷与SLY蛋白分子结构间的相互作用。分子对接是药物研发的新手段和新途径,具有高效、快速、经济等优势,已在药品研发过程中发挥了巨大的作用。目前已有不少中药单体被证实可靶向SLY蛋白抑制其溶血活性。经分子对接分析,桑色素可与SLY的第2结构域相互结合发挥作用,杨梅素主要结合于SLY第2及第3结构域的间隙,而白皮杉醇则可嵌入蛋白第2及第4结构域之间的联合区域。虽然结合位点不同,这些药物却均可改变SLY蛋白单体构象阻止其寡聚化,从而降低蛋白对宿主细胞膜的破坏。由此可见,SLY的生物活性功能与其空间结构密切相关,空间结构的变化会直接导致其生物功能的降低或丧失。与之前研究所报道的药物结合区域不同,本次分子对接结果表明毛蕊花糖苷可与SLY蛋白残基结合从而嵌入SLY的第4结构域。因此,我们推测毛蕊花糖苷结合SLY第4结构域后改变了蛋白的原有结构使其功能最终发生变化。体外红细胞溶血试验进一步证实了毛蕊花糖苷可直接抑制SLY蛋白介导的溶血活性。
此外,体内研究显示,经毛蕊花糖苷治疗后感染小鼠的死亡率降低,存活状况得到明显改善,这一结果直接表明毛蕊花糖苷可以降低猪链球菌致病力。但我们发现毛蕊花糖苷对感染小鼠尚未达到完全保护的效果,造成这一现象的原因可能在于除SLY之外,猪链球菌还会产生许多与其致病性相关的毒力因子,例如肽聚糖(peptidoglycan, PG)、荚膜多糖(capsular polysaccharide, CPS)、溶菌酶释放蛋白(muramidase-released protein, MRP)和细胞外蛋白因子(extracellular, EF)等,它们均参与了链球菌侵袭宿主的过程。本研究中毛蕊花糖苷虽抑制了SLY的溶血活性,但是其他毒力因子可能仍在发挥作用,从而导致毛蕊花糖苷无法完全消除链球菌对小鼠的致病性。因此开发对抗2型猪链球菌多种毒力因子的联合用药应成为下一步的研究重点。
CDCs家族目前已知包含了40余种成员,其中SLY、肺炎链球菌溶血素(pneumolysin, PLY)、产气荚膜梭菌溶血素(perfringolysin, PFO)、单增李斯特菌溶血素(listeriolysin, LLO)及化脓链球菌溶血素(streptolysin O, SLO)等蛋白成员氨基酸序列多处相近甚至相同,且具有高度相似的三级结构。同时该家族大多成员与SLY的致病机制相同,即在宿主靶细胞膜上形成孔洞破坏其完整性。目前毛蕊花糖苷已被证实可结合在PLY的第3及第4结构域的间隙,通过改变PLY空间构象抑制其溶血活性,从而降低肺炎链球菌(, pneumococcus)对小鼠的致病力。结合本次研究结果,我们猜测毛蕊花糖苷可能是一种广谱抗CDCs溶血活性的天然化合物,对CDCs家族其他成员的溶血活性也有抑制作用,之后我们将对这一猜想继续进行验证。
4 结论
在本研究中,我们发现毛蕊花糖苷不会影响2型猪链球菌的生长活性,但是该药物可对该病原菌产生的SLY活性产生显著的抑制作用。同时,毛蕊花糖苷可明显降低2型猪链球菌对小鼠的致病性,起到保护机体的作用。本研究结果表明,毛蕊花糖苷是一种潜在的SLY蛋白活性抑制剂,为后续临床开发抗2型猪链球菌的新药提供了一定的理论基础。
猜你喜欢
天津农业科学(2022年5期)2022-05-31
江苏农业科学(2020年8期)2020-06-01
家禽科学(2020年3期)2020-05-13
江苏农业科学(2019年14期)2019-09-23
湖北农业科学(2017年7期)2017-05-13
中文信息(2017年2期)2017-04-13
江苏农业科学(2016年11期)2017-03-21
智能计算机与应用(2016年4期)2016-09-26
健康之路(医药研究)(2015年2期)2015-10-21
江苏农业科学(2015年1期)2015-04-17
