
不同CO2浓度下闽楠幼苗光合作用及氮素分配的响应机制
2022-08-26 10:59韦小丽吴高殷
西南林业大学学报 2022年4期
王 晓 韦小丽,2 吴高殷 梁 娴
(1. 贵州大学林学院,贵州 贵阳 550025;2. 贵州省森林资源与环境研究中心,贵州 贵阳 550025)
自工业革命以来,大气中的二氧化碳(CO2)浓度日趋升高,政府间气候变化专门委员会(IPCC)第4次评估报告指出,目前大气中的CO2摩尔分数已经由工业化革命前的260~280 μmol/mol上升到2011年的393 μmol/mol,而且还在以每年1.5~2.0 μmol/mol的速度上升[1]。根据专家推测,到本世纪中,大气中CO2摩尔分数将会升高到550 μmol/mol左右,到21世纪末将上升到700~1260 μmol/mol[2]。大气中CO2浓度升高是工业革命以来全球范围内最重要的生态变化之一,直接影响植物的生长发与和代谢过程。光合作用是植物生长以及实现碳汇功能的重要生理过程,其以CO2为反应原料,效率受CO2浓度变化的影响。CO2浓度升高影响植物光合效率的同时也会对植物的其他生命活动产生重要影响[3−6]。
氮是植物生长必需的大量营养元素之一,Pettersson等[7]的研究指出,植物光合作用在高 CO2浓度下的适应响应取决于氮的供应水平,同时受植株整体水平的源库关系调节。氮素营养不足会限制植物对CO2浓度升高的响应,当氮成为限制因素时,核酮糖−1,5−二磷酸羧化/加氧酶(Rubisco)中的氮也可能会重新分配到其他酶类或者其他组织和器官,从而对Rubisco进行调节,这使得植物光合作用更加受到Rubisco的限制[8]。Wujeska-Klause 等[9]的研究表明,CO2浓度升高会限制植物对氮素吸收利用从而影响植物对CO2浓度升高的积极响应。
CO2浓度升高对闽楠(Phoebe bournei)光合作用的影响已有初步的报道,韩文军等[10]的研究结果表明,CO2体积分数加倍处理可以使闽楠的光合速率明显降低,使其出现严重的光合下调现象,同时闽楠叶绿素含量、可溶性糖含量、可溶性蛋白含量、光呼吸速率以及暗呼吸速率均降低,严重影响闽楠的生长。也有研究表明,CO2浓度升高会对闽楠幼苗可溶性糖等物质的积累以及碳氮代谢产生不利影响,进而阻碍闽楠生长[11−12]。CO2浓度升高影响闽楠幼苗光合效率的同时,是否影响CO2响应以及光响应特征参数,闽楠幼苗叶片氮素营养状况、分配及利用是否发生改变等相关报道未作深入研究。本研究以1年生闽楠幼苗为研究对象,研究不同CO2浓度升高对闽楠幼苗光合特性、氮素营养尤其是叶片氮素分配的影响,明确CO2浓度升高条件下闽楠幼苗光合作用尤其是氮素营养的表现,研究结果丰富CO2升高条件下闽楠幼苗光合下调现象出现机理研究的同时为CO2浓度升高条件下闽楠的栽培调控措施研究提供参考。
1 材料与方法
1.1 试验材料
供试材料为采自同一母株的种子培育的闽楠1年生容器苗。2019年1月选取长势一致的健壮容器苗(苗高为(11.26 ± 0.14) cm、地径为(3.54 ±0.36) mm)移栽至塑料花盆中(15.5 cm × 14.0 cm)。盆栽土壤为黄壤,盆栽土壤特性:pH(4.76)、全氮(0.63 g/kg)、碱解氮(22.1 mg/kg)、全磷(0.22 g/kg)、有效磷(6.2 mg/kg)、全钾(19.8 g/kg)、速效钾(213.7 mg/kg)。每盆装风干土1.5 kg,栽植幼苗1株,移栽后缓苗4个月选取长势一致健壮盆栽幼苗进行试验处理。
1.2 试验设计
采用开顶式气室(OTC)模拟大气CO2浓度升高。OTC的外形为正八边形棱柱状,边长1 m,对边直径2.42 m,高1.7 m;侧面为4 cm × 4 cm方钢框架,高透光性8 mm钢化玻璃。为聚拢气体并使气体散失速度放缓,OTC顶部开口向内斜45°。气室内CO2浓度采用GMP−220 CO2传感器装置控制(VAISALA,芬兰),采集后的数据传输到主机,主机通过运行设定的程序分别控制各气室的电磁阀的开关,将气室内的CO2浓度控制在目标浓度范围内。试验设置3个CO2浓度水平:自然水平(CK)、高浓度CO2水平1(T1)、高浓度CO2水平2(T2),CK气室通入自然空气(350 ± 70) μmol/mol,T1气室通入CO2气体,控制CO2浓度为(500 ± 100) μmol/mol,T2气室通入CO2气体,控制CO2浓度为(700 ± 10) μmol/mol,不同CO2浓度气室温度相差0.5~1 ℃。每个浓度设置3次重复,每个重复15株苗。于2019年6月16日—8月26日开始通气,处理结束后取样测各试验指标。
1.3 指标测定方法
1.3.1 叶绿素含量测定
每个重复选取5株,取幼苗中上部功能叶称量0.15 g,采用乙醇浸提,用紫外分光光度法测定浸提液在663 nm和645 nm下的吸光度(OD663、OD645),根据公式(1)~(2)计算叶绿素a含量(Ca)、叶绿素b含量(Cb)。
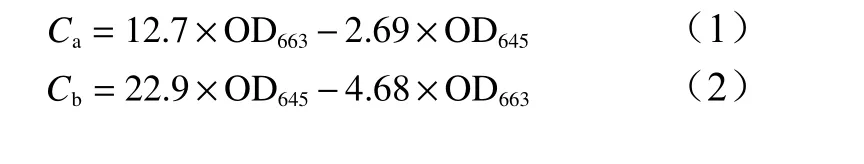
1.3.2 净光合速率测定
采用Li−6400XT便携式光合作用测量系统(LI−COR,美国),每个重复选取3株苗,每株选取中上部3片完全展开的功能叶,于晴天9:00—11:00测定,测定时采用 CO2小钢瓶控制aCO2气室测定时叶片叶室CO2浓度350 μmol/mol,T1气室测定时叶片叶室CO2浓度为500 μmol/mol,T2气室测定时叶片叶室CO2浓度为700 μmol/mol,利用系统自配红蓝光源补光,光量子通量密度设定为1200 μmol/(m2∙s)。
1.3.3 CO2响应曲线测定
采用Li−6400XT便携式光合作用测量系统(LI−COR,美国)测定不同处理闽楠幼苗叶片CO2响应曲线。测定前1000 μmol/(m2∙s)光强对叶片进行光诱导20 min左右,CO2浓度梯度为400、300、100、50、150、250、400、800、1200、1500 μmol/mol,最小等待时间120 s,最大等待时间200s。采用非直角双曲线模型对曲线进行拟合,测得最大RuBP羧化速率(Vc max)、RuBP再生光合电子传递速率(Jmax)。
1.3.4 光响应曲线测定
采用Li−6400XT便携式光合作用测量系统(LI−COR,美国)测定不同处理闽楠幼苗叶片光响应曲线。测定前1 000 μmol/(m2∙s)光强对叶片进行光诱导20 min左右,光合有效辐射强度为1800、1400、1000、800、500、250、150、100、60、30、0 μmol/(m2∙s),最小等待时间120 s,最大等待时间200 s。采用非直角双曲线模型对曲线进行拟合,测得光补偿点(LCP)、光饱和点(LSP)、最大净光合速率(Amax)、暗呼吸速率(Rd)、表观量子效率(AQE)。
1.3.5 叶片氮含量及叶面积测定
采用凯氏定氮法测得叶片氮含量(NM),采用扫描法计算叶面积,根据叶面积计算比叶质量(SLW)以及单位面积叶氮含量(Narea)。
1.3.6 光合机构氮素分配及光合氮利用效率(PNVE)计算
氮在叶片光合机构中的分配主要分3部分:RuBP羧化的氮素(NC)、光合电子传递和磷酸化的氮素(NB)及捕光组分的氮素(NL)。计算公式参考Niinemets 等[13]。


式中:Vcr为Rubisco比活度,Jmc为以细胞色素为基础的潜在光合电子流,CC为叶绿素含量,CB为结合在PSⅠ、PSⅡ以及LHCⅡ上的叶绿素含量。
1.4 数据统计与分析
用 Excel 2007进行数据的录入和图表的绘制,采用 SPSS 20. 0对数据进行统计分析,采用单因素方差分析和多重比较检验分析数据差异显著性,试验数据用平均值 ± 标准差表示。
2 结果与分析
2.1 CO2浓度升高对闽楠幼苗CO2响应曲线及特征参数的影响
由图1可知,当CO2浓度小于400 μmol/mol时,不同处理闽楠幼苗叶片净光合速率随CO2浓度增加快速增加,当CO2浓度高于400 μmol/mol时,净光合速率增加速度较慢且逐渐稳定趋于饱和状态;不同CO2浓度条件下Pn的基本表现为T1> CK > T2。

图1 不同处理对闽楠幼苗CO2响应曲线的影响Fig. 1 Effect of different treatments on CO2 response curve of P. bournei seedlings
由图2可知,对不同处理闽楠幼苗CO2响应曲线进行拟合得到Vc max和Jmax,不同CO2浓度升高条件对Vc max的影响表现为T1条件增加,T2条件降低,T1条件下闽楠幼苗Vcmax显著高于CK和T2(P< 0.05),分别比CK、T2提高6.79%、21.65%,且两两间差异显著(P< 0.05);不同CO2浓度对Jmax无显著影响。说明CO2浓度不同升高程度对闽楠幼苗最大电子传递速率没有显著影响,对最大羧化速率的影响因CO2浓度升高程度而不同。

图2 不同处理对闽楠幼苗最大羧化速率即最大电子传递速率的影响Fig. 2 Effects of different treatments on Vcmax and Jmax of P. bournei seedlings
2.2 CO2浓度升高对闽楠幼苗光响应曲线及特征参数的影响
由图3可知, 当光强度小于250 μmol/(m2∙s)时,不同CO2处理闽楠幼苗叶片净光合速率均快速增加;当光强度在250~800 μmol/(m2∙s)时,不同CO2处理闽楠幼苗叶片净光合速率增速逐渐降低,且不同处理间的差异逐渐增大;光强度达到800 μmol/(m2∙s)时,T2处理闽楠幼苗叶片净光合速率逐渐趋于稳定;当光强度大于800 μmol/(m2∙s)时,CK及T1各处理闽楠幼苗叶片净光合速率逐渐趋于稳定。不同光强度条件下基本表现为T1> CK > T2。由表1可知,拟合得到的不同CO2处理闽楠幼苗光响应特征参数中,不同CO2浓度处理对闽楠幼苗LCP、Rd以及AQE无显著影响;T1处理的LSP、Amax略高于CK,显著高于T2(P< 0.05),分别高40.67%和29.44%。说明适当增加CO2浓度会增加幼苗的光饱和点和最大净光合速率。

图3 不同处理对闽楠幼苗光响应曲线的影响Fig. 3 Effects of different treatments on photoresponse curve of P. bournei seedlings

表1 不同处理对闽楠幼苗光响应特征参数的影响Table 1 Effects of different treatments on characteristic parameters of photoresponse of P. bournei seedlings
2.3 CO2浓度升高对闽楠幼苗叶片氮素分配的影响
由表2可知,不同CO2浓度升高条件对闽楠幼苗叶片氮素在各机构的分配影响不同。CO2浓度升高对闽楠幼苗NL、NT影响不显著,对SLW、NM、Narea、NC、NB的影响依CO2浓度升高程度不同,即T1与CK无显著差异,但显著高于T2(P< 0.05)。T1CO2处理的SLW、NM、Narea、NC较T2处理分别增加8.71%、7.33%、16.70%、15.97%,NB减少14.45%。说明一定程度的CO2浓度升高对闽楠幼苗叶片氮素积累及分配不会产生显著影响,但较高程度的CO2浓度升高在降低闽楠幼苗叶片氮素含量的同时会减少氮素在RuBP羧化作用的分配。
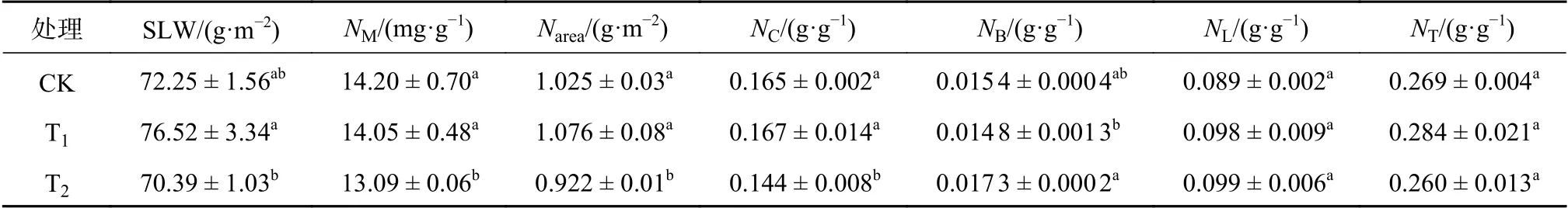
表2 不同处理对闽楠幼苗叶片氮素分配的影响Table 2 Effects of different treatments on nitrogen partitioning in leaves of P.bournei seedlings
2.4 CO2浓度升高对闽楠幼苗叶片光合氮利用效率的影响
不同CO2浓度处理对闽楠幼苗叶片光合氮利用效率的影响不同(图4),CK处理闽楠幼苗PUNE与T1没有显著差异,但两者的PUNE与T2差异显著(P< 0.05),T1、CK的PUNE分别比T2高17.34%、13.55%,说明过高的CO2浓度处理降低了闽楠的光合氮利用效率。

图4 不同处理对闽楠幼苗叶片光合氮利用效率的影响Fig. 4 Effects of different treatments on PNUE of P. bournei seedlings
3 结论与讨论
植物CO2响应曲线特征参数中Vcmax和Jmax是表征植物光合能力的重要参数[15]。本研究结果表明,CO2浓度升高对闽楠幼苗Jmax无显著影响,主要影响Vcmax,T1和T2较CK分别显著提高和降低闽楠幼苗Vcmax,说明适当的CO2浓度升高可以提高闽楠幼苗羧化反应效率,促进光合作用,而CO2浓度大幅度增加则不利于闽楠幼苗的碳同化。T1的Vcmax提高的原因可能是CO2浓度的适当升高增加了光合作用的底物进而提高羧化反应效率[16]。T2的Vc max降低原因可能是叶片Rubisco含量降低[11]。Vc max是RuBP的最大羧化速率,与Rubisco含量成正比,T2的闽楠幼苗叶片氮含量降低导致Rubisco含量减少的同时降低Vc max。光合作用是植物利用光能的过程,CO2浓度升高影响闽楠幼苗光合效率、叶绿素含量以及对CO2浓度响应的同时可能也会影响到对光照的响应[17]。本研究结果表明,CO2浓度升高对闽楠幼苗LCP、Rd以及AQE无显著影响,主要影响LSP和Amax,其中T2处理显著降低闽楠幼苗Amax,而T1处理对闽楠幼苗Amax则无显著影响。T2处理下闽楠幼苗Amax的降低说明闽楠幼苗的光能利用能力和对光环境的适应性都有所降低,这也可能是T2条件下闽楠幼苗出现光合下调现象的原因。
植物叶片的氮含量与光合作用具有显著相关性,通常情况下,植物叶片氮含量越高,光合效率也越高[18]。本研究结果中,T2处理显著降低闽楠幼苗叶片单位质量以及单位面积的氮素含量,而T1处理对闽楠幼苗叶片单位质量以及单位面积的氮素含量无显著影响,说明CO2浓度适当升高对闽楠幼苗叶片的氮素吸收不会产生不利影响,而CO2浓度大幅度升高则会影响闽楠幼苗叶片对氮素的吸收,因而抑制光合作用。有研究也指出,CO2浓度升高条件下植物光合效率变化是CO2浓度升高对植物氮素吸收代谢的影响所致[7]。CO2浓度升高对植物光合作用的影响与植物体内的氮素状态具有密切联系[19]。除了叶片氮含量影响叶片光合速率,氮素在叶片中的分配对叶片的光合速率以及环境变化的适应性也有重要的影响[20−21]。本研究结果表明,T1的闽楠幼苗叶片氮含量及其在RuBP羧化部分的分配较CK无显著变化,T2的闽楠幼苗叶片氮含量及其在RuBP羧化部分的氮素分配则显著低于CK和T1,说明CO2浓度显著升高影响闽楠幼苗叶片氮含量和氮素在RuBP羧化部分的分配。RuBP羧化是光合作用碳固定的重要反应,由Rubisco催化,Rubisco是植物叶片中最丰富的蛋白质,约占叶片可溶性蛋白的50%[22]。T2的闽楠幼苗叶片氮素在RuBP羧化部分减少会降低叶片内Rubisco的含量,影响碳同化的同时影响光合效率,而这也可能是T2处理闽楠幼苗出现光合下调现象的关键原因[11]。Zhang等[23]的研究也发现CO2浓度升高条件下入侵草类薇甘菊(Mikania micrantha)和飞机草(Chromolaena odorata)叶片氮含量降低的同时Rubisco含量也降低。
氮素是植物生长必须的大量矿质元素,氮素营养状况直接影响植物的各个生理过程[24−26]。光合氮利用效率即光合速率与叶片氮含量的比值,其是反映植物叶片氮素利用和光合生理的重要参数[27]。有研究表明,CO2浓度升高会降低桉树的光合氮利用效率[28]。本研究结果表明,T2显著降低闽楠幼苗叶片的光合氮素利用效率,T1对其则无显著影响,说明适当的CO2浓度升高不会影响闽楠苗光合氮利用效率,大幅度升高则对光合氮素利用效率起到抑制作用,同时CO2浓度大幅度升高处理闽楠幼苗氮素含量及光合速率均降低而光合氮素利用效率即两者比值同时降低,说明该处理闽楠幼苗光合速率下降幅度更大。T2下闽楠幼苗光合氮利用效率降低会影响幼苗体内各种酶类的合成进而影响生理代谢,其中光合作用的关键酶Rubisco的合成可能因为光合氮利用效率的降低受到不易影响,最终抑制光合作用,因此不同处理闽楠幼苗叶片的光合氮素效率也是导致光合速率不同以及闽楠幼苗出现光合下调现象的原因[11, 29]。
适当的CO2浓度升高不会影响闽楠幼苗的光合作用及氮素利用,CO2浓度大幅度升高则会降低闽楠幼苗叶片氮素含量以及氮素在光合机构RuBP羧化部分的分配,同时降低叶片光合氮利用效率,最终影响光合效率。改变闽楠幼苗叶片的氮素状态对CO2浓度升高条件下调控光合效率具有重要研究价值。
猜你喜欢
灌溉排水学报(2022年9期)2022-09-28
农业工程学报(2022年8期)2022-08-08
作物学报(2022年10期)2022-07-21
作物学报(2022年9期)2022-07-18
安徽农学通报(2022年9期)2022-06-14
作物学报(2022年3期)2022-01-22
物联网技术(2016年11期)2017-01-12
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
