
杉木生长性状的空间与竞争效应及其对遗传参数估计的影响*
2022-08-20 06:18史月冬叶代全施季森边黎明
林业科学 2022年5期
史月冬 郑 宏 叶代全 施季森 边黎明
(1.南京林业大学林木遗传与生物技术教育部重点实验室 南京 210037; 2.南京林业大学南方现代林业协同创新中心 南京 210037; 3.福建省洋口国有林场 南平 353000)
加性遗传方差(基因的加性效应方差)是林木遗传分析中的重要参数,其可确定一个群体目标性状的遗传结构和变异量,影响林木育种值预测和优良单株选择的准确性及产生的遗传增益,为林木育种策略制定提供理论依据(康向阳, 2019)。为避免估计加性遗传方差时产生较大偏差,应在构建的林木遗传分析模型中估算重要的遗传变异来源以及可识别的空间、竞争效应。林木个体间的竞争效应普遍存在,是遗传和环境共同作用的结果,受树种自身遗传特性和相邻个体空间分布的影响,群落中生态位越相似的物种,竞争和选择压力越大,种内竞争往往比种间竞争更为激烈(Adleretal., 2018)。在林木遗传测定中,Muir(2005)首次采用整合竞争效应的模型,利用似然法估计方差分量,得到遗传参数,并结合加性遗传与竞争效应构建指数选择方法,选择优良个体。Cappa等(2008)采用贝叶斯法估计林木个体间的竞争效应和总加性遗传效应,发现火炬松(Pinustaeda)个体基因型间存在显著的竞争效应,且竞争效应的遗传变异约占总变异的10%。Costa e Silva等(2017)在遗传水平上分析蓝桉(Eucalyptusglobulus)的竞争效应,认为拟合竞争效应的模型可减小生长性状遗传变异的估计值。在林木遗传试验分析中,理想的模型是在常规试验设计基础上,整合空间与竞争效应(Cappaetal., 2015)。对农作物的研究表明,同时拟合空间与竞争效应能够显著增加加性遗传方差,降低残差方差,选择精度可达84%(Stringeretal., 2011)。Cappa等(2016)对花旗松(Pseudotsugamenziesii)多地点试验的遗传分析发现,35年生时径生长在遗传和环境水平上显示出强烈的竞争效应,但竞争效应对高生长的影响不显著,整合空间与竞争效应的模型比其他模型的拟合效果更好;采用贝叶斯法,对比不同模型,也得出整合空间与竞争效应模型方法的优良性(Cappaetal., 2015);在包含竞争与空间效应模型的个体亲缘关系矩阵中,利用个体基因组信息,可以提高育种值预测的准确性(Cappaetal., 2017)。
杉木(Cunninghamialanceolata)为亚热带常绿针叶树种,是我国南方重要的造林用材树种之一(吴中伦, 1955),具有生长快、干形通直圆满、出材率高、材质好、纹理直和防虫防腐等优点,其遗传测定林主要营建在丘陵山地,株行距一般为2 m×2 m(施季森等, 1993)。目前,基于最佳线性无偏预测(best linear unbiased prediction, BLUP)(Henderson, 1975)的混合线性模型方法已广泛应用于杉木遗传试验分析(陈兴彬等, 2018),能够估计试验设计和空间效应(边黎明等, 2015; 2020; Bianetal., 2017),但是没有考虑试验林所存在的竞争效应。
本研究的目的在于运用整合试验设计效应、空间效应和竞争效应的混合线性模型方法,以不同林龄阶段杉木自由授粉子代的生长性状为目标,比较模型拟合效果,选择不同林龄阶段的最优模型,估计方差分量和遗传参数,分析空间与竞争效应对遗传参数估计值和选择效果的影响,以更加准确地鉴定性状的遗传变异,为杉木高世代育种策略制定提供参考。
1 材料与方法
1.1 试验地概况
试验林营建在福建省洋口国有林场南山工区新林地(林班号: 2-1-3f),位于武夷山支脉的低山丘陵,属亚热带湿润季风气候,年均气温18.5 ℃,最高气温40.3 ℃,最低气温-6.8 ℃,年均降水量1 880 mm,无霜期280天,相对湿度82 %,海拔230~300 m,土壤为花岗岩发育的山地红壤(郑宏等, 2016)。造林前,试验地经过炼山、烧杂和带状整地,株行距为2 m×2 m。
1.2 试验材料与试验设计
参试的自由授粉家系采自杉木三省种子园,其家系分别来自广西(1-11号)、福建(12-22号)、湖南(23-33号),对照(CK)为福建省洋口国有林场杉木第1代种子园混合种子。1980年在福建省洋口国有林场育苗, 1981年春季造林,采用完全随机区组试验设计,重复30次,单株小区。分别于1985年(林龄5年)、1991年(林龄11年)和2020年(林龄40年)调查每个个体的树高和胸径,作为幼林龄阶段、中林龄阶段和过成熟林阶段的代表。1991年调查后,采取伐去被压木、濒死木和枯立木的措施。
1.3 模型与方法
单性状和多性状的遗传分析基于最佳线性无偏预测(BLUP)的混合线性模型方法(Henderson, 1975)。空间模型自相关最优值和竞争效应估计分别采用Echidna软件(www.echidnamms.org)和BreedR包(https:∥github.com/famuvie/breedR)。
1.3.1 基本模型(试验设计效应模型,standard mixed linear model, MLM)
y=Xβ+Za+e。
(1)
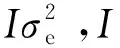
1.3.2 空间模型(一阶自回归混合线性模型,first-order autoregressive mixed linear model,AR1)
y=Xβ+Za+ξ+η。
(2)

1.3.3 竞争模型(competition model, CM)
y=Xβ+Zdad+Zcac+e。
(3)
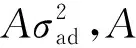
总加性方差矩阵为:
(4)
G0为加性遗传方差-竞争效应方差之间的方差-协方差结构,⊗表示克罗内克积(Kronecker producetion)。
1.3.4 空间-竞争模型(spatial-competition model, CSM)
y=Xβ+Zdad+Zcac+ξ+η。
(5)
1.3.5 模型比较 赤池信息准则(Akaike information criterion, AIC)大小表示模型拟合的优良程度,AIC越低说明模型拟合程度越好(Akaike, 1974)。
AIC=-2×(logL-p)。
(6)
式中:lgL为模型迭代收敛的似然值;p为模型中随机效应的数量。模型随机效应的显著性根据方差值与其标准误差之比判定,当比值介于0~1之间则认为效应不显著; 当比值介于1~2之间,根据似然比进行检验; 当比值大于2时,效应显著。
1.3.6 狭义遗传力
(7)
(8)
式(7)和(8)分别代表基本模型与空间模型、竞争模型与空间-竞争模型的狭义遗传力计算方法。
1.3.7 遗传相关 遗传相关包括同一个体不同性状的加性遗传相关(raiaj)、同一个体不同性状的竞争遗传相关(rcicj)(Hernndezetal., 2019)、不同个体间同一性状的加性-竞争遗传相关(raici)不同个体间不同性状之间的遗传相关系数(raicj)。不同遗传相关系数的估计公式如下:
(9)
(10)
(11)
(12)
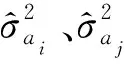
1.3.8 育种值准确性 育种值真实值与预测值之间的相关性称为育种值准确性(r),计算公式为:
(13)
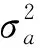
1.3.9 遗传增益
(14)
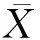
2 结果与分析
2.1 性状表型特征值
生长性状的统计特征值如表1所示。林龄5年时,CK胸径均值为6.5 cm,比林分总体胸径均值低10.95%。11号家系胸径均值最大,比对照组高19.5%。林龄11年时,CK胸径均值为13.6 cm,比林分总体胸径均值低14.46%,11号家系胸径均值比对照组家系高20.3%。林龄40年时,CK胸径均值为28.1 cm,比林分总体胸径均值高8.74%。幼林龄对照组树高均值为4.3 m,比林分总体树高均值低10.18%; 中林龄对照组树高均值为12.0 m,比林分总体树高均值低1.72%; 过成熟林对照组树高均值为20.0 m,比林分总体树高均值高0.10%。测定林单株在过成熟林阶段数量减少的原因是1991年采取了留优去劣择伐的过程。生长性状变异中等偏弱,胸径变异系数略大于树高,幼林龄杉木表现出林分分化趋势,中林龄变异系数有所下降,说明此时林分较为整齐,间伐后杉木生长潜力得到充分表达,过成熟林变异系数有一个增大的变化过程,符合杉木试验林在天然状态下的生长规律。

表1 生长性状的统计特征值Tab.1 Statistical characteristics for growth traits
2.2 空间效应与竞争效应
在拟合基础模型的基础上,计算中心株残差与表型邻株之间的皮尔森相关系数(表2),用于量化试验林个体不同方向的竞争效应。幼林龄阶段,杉木胸径在列方向之间的相关性为负,说明胸径在列方向存在一定竞争性,树高在各个方向之间相关性皆为正值,树高间不存在明显竞争。中林龄阶段,性状之间相关性在各个方向均为负值,行方向值超过对角线值,说明竞争效应强弱与林分个体间的距离有关,也说明竞争效应在中林龄表现强烈。过成熟林阶段,林分个体在行、列和对角线上依旧存在个体间的竞争效应。生长性状行之间的自相关性小于列之间的自相关性,列自相关指数大于0.8,试验林列之间的环境异质性较为强烈,这种异质性可能与地形的变化存在关联。中林龄阶段,环境异质性偏低的原因可能在于林分间个体生长较为整齐;过成熟林阶段,试验林列之间的环境异质性程度高于行之间,表明地形对试验林生长性状存在影响。
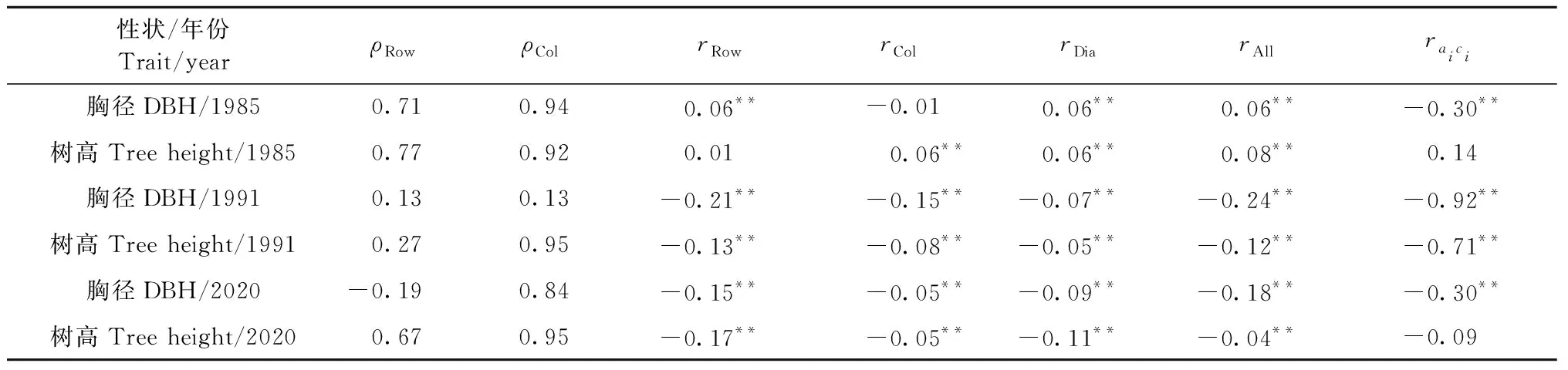
表2 中心株的基础模型残差与1阶邻株表型的皮尔森相关系数①Tab.2 Pearson correlation coefficients between the residual from the standard mixed linear model of the focal tree and the phenotypic mean of its first-order neighbours
2.3 模型选择与遗传参数估计
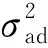
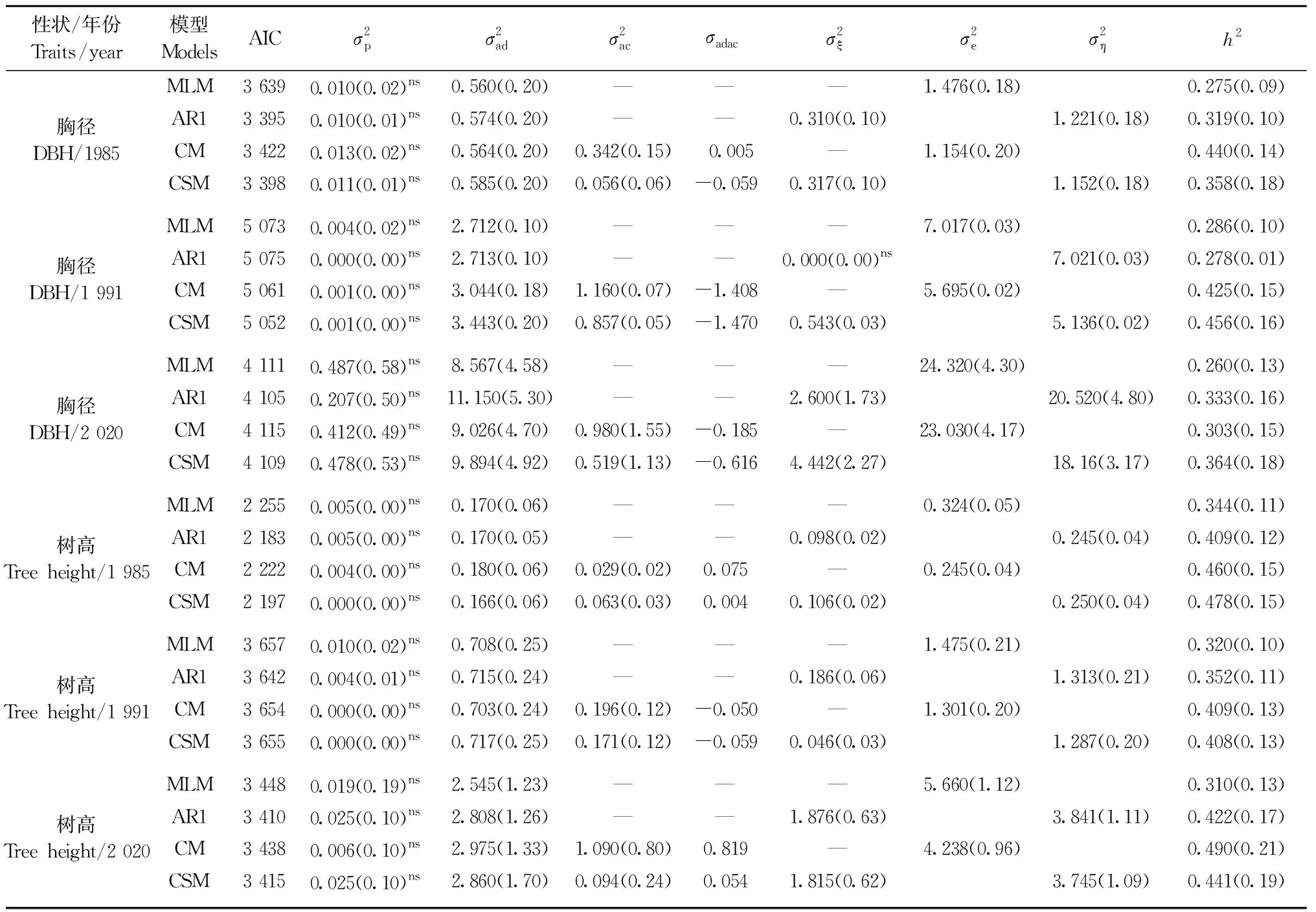
表3 4种模型及其遗传参数估计值的比较①Tab.3 Comparison of four models and their estimated values of genetic parameters
不同林龄空间-竞争模型中的遗传相关如表4所示, 性状间加性遗传相关(raiaj)极为显著,说明控制胸径与树高基因位点大部分相同,二者之间存在一因多效作用(边黎明等, 2020)。竞争遗传相关表明,同一个体不同性状之间竞争效应相关系数也处于极显著水平。不同个体相同性状间加性-竞争遗传相关(raici)为负显著,胸径间负相关范围介于-0.30~-0.92,树高负相关介于0.14~-0.71。1991年性状负相关绝对值最大,说明林分间表现出强烈竞争作用。不同个体不同性状加性-竞争遗传相关表明,林分中目标单株胸径受邻株树高竞争效应作用小,而树高受邻株胸径竞争竞争作用大,其原因可能是林分间性状对光资源的争夺加剧引起的(Brotherstoneetal., 2011)。不同竞争遗传相关表明,胸径比树高更能感受到竞争的敏感程度。

表4 生长性状间的遗传相关①Tab.4 Genetic correlations between the growth traits
2.4 育种值预测的准确性及优良单株选择
混合模型中拟合空间、竞争效应可能影响选择的效率和准确性,对应在实际应用中则是不同模型的育种值排名是否出现较大变化。利用斯皮尔曼相关检验2020年各模型胸径、树高加性效应值之间的相关性,结果表明2个性状各模型加性效应值以及竞争模型总育种值(加性效应值与竞争效应值之和)之间表现出极强正相关性(图1)。
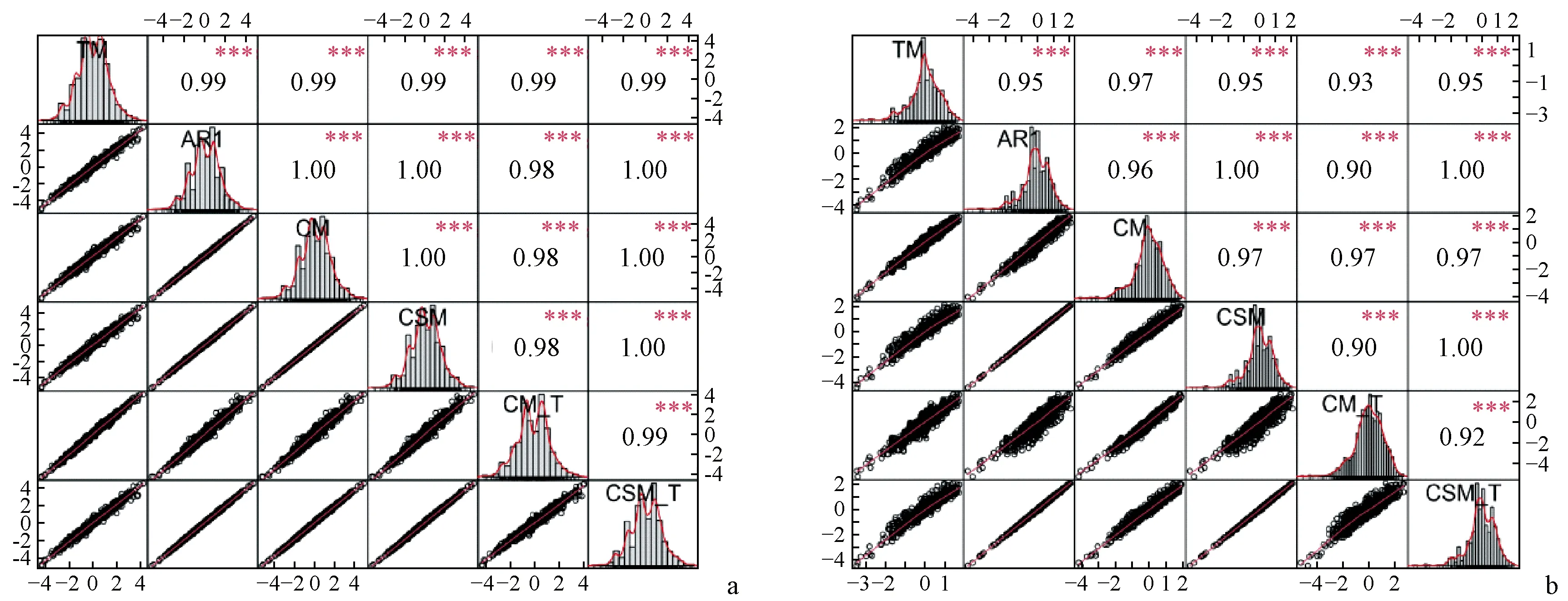
图1 生长性状加性效应值间的相关性Fig. 1 Correlations among the additive effect values of growth traitsa、b分别表示胸径、树高的加性效应值(TM、AR1、CM、CSM)以及总育种值(CM_T、CSM_T)之间的斯皮尔曼相关系数。a,b show the Spearman correlation coefficients between the DBH and tree height (TM, AR1, CM, CSM) and the total breeding values (CM_T, CSM_T) respectively.
提高杉木选择强度也是提高选择效益的重要手段,对林龄40年的个体进行优良单株选择,设定单株选择强度为5%,结合4个模型的加性效应值进行排序,每个模型选择23个单株(表5),其中135、230、369号为10号家系, 525和1002号为13号家系, 688和1024号为27号家系, 165、228、353、738和840号为当地对照组家系,结果发现当地对照组家系存在较好的本土适应性。
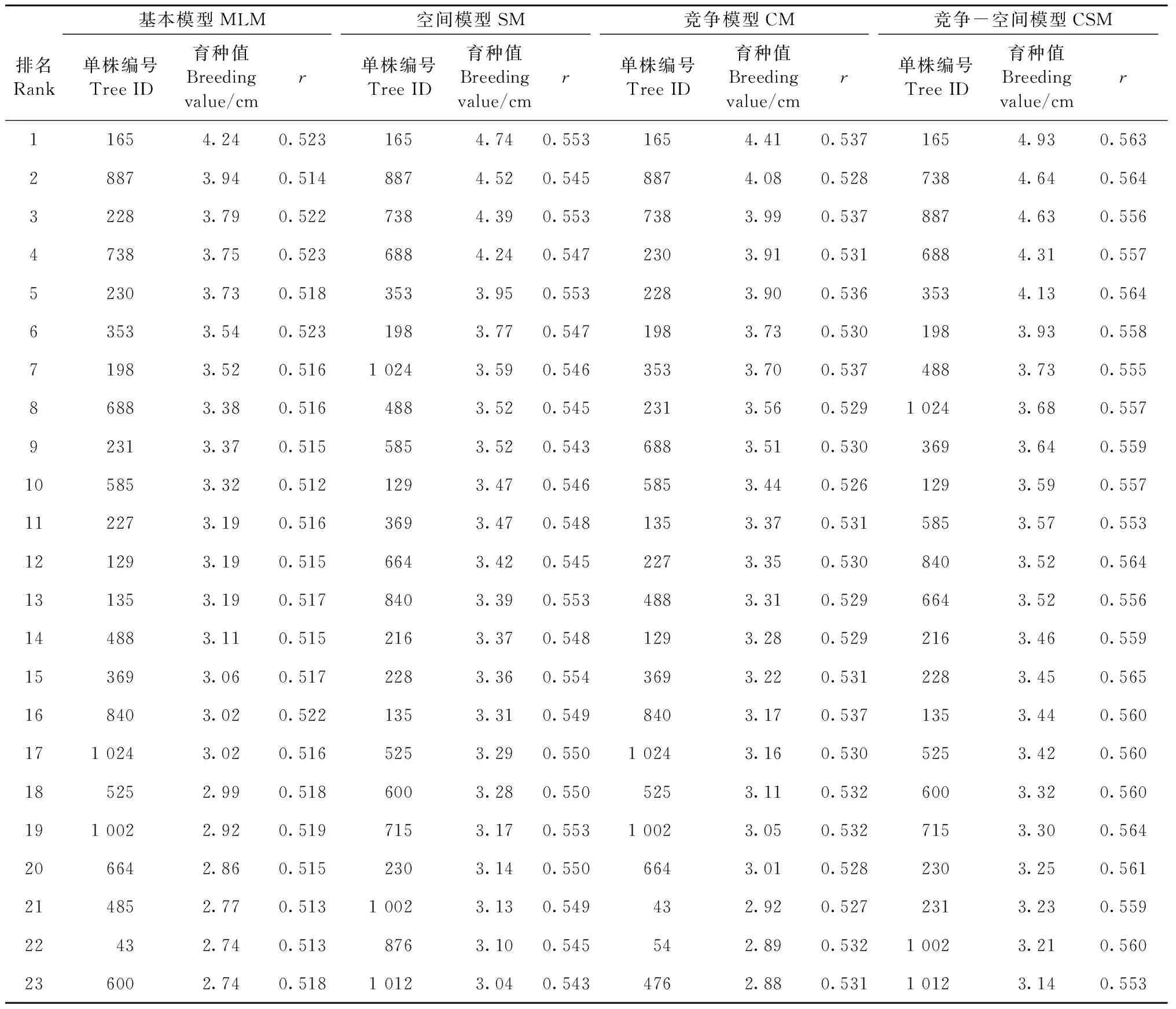
表5 优良单株的育种值排名Tab.5 Ranking of elite trees’ breeding values
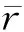
3 讨论
3.1 试验林个体保存率与竞争效应的关系
竞争效应在林分间是不平衡的,弱小单株更易受到林分中更大个体的抑制作用(Cannelletal., 1984),竞争效应与林分规模和资源分配有关,单位面积林木密度是决定竞争效应的重要因素(Bijmaetal., 2007)。本研究试验林幼林龄、中林龄阶段林分保持率在85%以上,而2020年林分保持率为45%,由于缺乏试验林间伐后的原始数据,故不能定量比较试验林在不同林分密度下竞争效应的差异程度。
3.2 空间-竞争效应对性状生长的影响
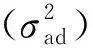
本研究通过2种方式定量试验林中的竞争效应。残差相关表明,林分间目标株受行、列、对角线之间邻近单株的竞争程度是不同的,表现为邻株之间距离越远竞争效应越弱。在对日本落叶松(Larixkaempferi) (董雷鸣, 2018)和柠檬桉(Corymbiacitriodorasubsp.variegata)(Hernndezetal., 2019)的研究中均表明,对角线方向单株对中心株的竞争性最弱。遗传相关表明,生长性状在中林龄阶段表现出显著的竞争效应。不同树种对竞争效应反应时间各不相同,对花旗松研究表明,胸径性状与材积性状分别在15和20年表现出较为明显的竞争效应(Yeetal., 2008);日本落叶松Carr)在19年生时生长性状竞争效应强烈(董雷鸣, 2018),表明树种在中林龄竞争效应强烈。胸径对竞争效应的敏感程度大于树高对竞争的敏感程度,可能是胸径易受林分间距离及林分规模的影响进而导致竞争效应加剧,因此竞争效应会对胸径的生长产生较大影响。
3.3 拟合空间-竞争效应对遗传参数估计的影响
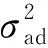
3.4 拟合空间-竞争效应对选择的影响
在模型中拟合竞争效应会改变选择的准确程度。本研究表明,拟合空间、竞争效应可以提高育种值预测的准确性。对柠檬桉胸径研究中表明采用拟合空间-竞争效应的试验进行分析,亲本和后代的育种值精度分别提高17%和28%(Hernndezetal., 2019)。在樟子松(Pinussylvestrisvar.mangolica)研究表明,拟合竞争、空间效应对亲本和后代的生长性状选择可提高46.03%和44.68%的育种值准确性,材性育种值准确度提高20%(Dutkowskietal., 2006)。目前的研究表明,拟合空间-竞争模型可得到更高的目标性状遗传增益,即遗传增益的提高是由拟合竞争效应而增加的。对比日本落叶松是否拟合竞争效应选择,二者仅有24%的个体同时入选,说明竞争效应对选择有较大影响。本试验研究表明,拟合竞争效应对于试验林没有显著影响,杉木试验林已经超过最佳选择的时机,但是,竞争效应对于选择的精准度依然有借鉴意义。
3.5 空间-竞争模型的适用性
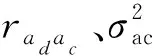
4 结论
当试验林中明显存在竞争效应时,采用竞争-空间模型拟合参数的程度更好,忽略环境异质性与竞争效应会导致更低的加性遗传参数值与更高的残差值。
林分个体间的竞争效应可从残差相关水平、遗传相关水平中得到量化。林分目标单株受到对角线方向单株的竞争性要弱于受到行、列方向单株的竞争。不同林龄阶段竞争效应是不同的,中林龄阶段生长性状所受到竞争效应最大,径生长对竞争的敏感性大于高生长。
猜你喜欢
作物学报(2022年1期)2022-11-05
水产养殖(2022年8期)2022-09-21
广东农业科学(2022年7期)2022-09-14
湖南林业科技(2022年4期)2022-08-29
防护林科技(2021年3期)2021-09-12
华人时刊(2021年21期)2021-03-09
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
动漫界·幼教365(中班)(2019年10期)2019-10-28
南方农业·下旬(2014年2期)2014-09-23
