
施肥对闽楠幼苗光合碳固定的影响*
2022-08-20 06:19冯金玲吴小慧黄蓝明杨志坚
林业科学 2022年5期
王 妍 冯金玲 吴小慧 黄蓝明 吴 娟 陈 宇 杨志坚
(1.福建农林大学林学院 福州 350002; 2.中国林学会 北京 100091)
光合作用是植物最重要的代谢,为一切生命活动提供必需的物质和能量,其同化合成的有机物质与植物生长密切相关(杨佳骏等, 2020)。它由光合磷酸化、碳同化以及光合产物合成等几个环节组成,各环节间互相配合、互相制约,在不同生长环境下相互作用大小不同(McClainetal., 2019; Zangetal., 2019; Liuetal., 2021)。光合作用除了受植物本身遗传特性影响外,还受各种环境因子影响,其中施肥是影响植物光合作用的主要因子之一(罗凡等, 2014; 罗杰等, 2017; 罗欢等, 2020)。
施肥改变植株体内的养分状态,与生理代谢关系密切,且影响植物光合作用(郝龙飞等, 2014)。光合碳固定作为植物体内的基础代谢,决定着光合产物的形成和转化、叶片内含物合成以及各种酶活性等(郑春芳等, 2020)。氮、磷和钾元素在植物光合作用和生理生化代谢过程中相互协调,相互制约(Lietal., 2018; 杨志坚等, 2021)。氮素是植物叶绿素和蛋白质的组成部分,氮素丰缺与叶绿素含量和酶活性密切的相系(刘连涛等, 2007; 刘敏等, 2019)。研究表明,施氮能提高植物幼苗的植株养分积累量,叶绿素含量、叶片净光合速率、蒸腾速率、胞间 CO2浓度、气孔导度、可溶性糖和可溶性蛋白含量,并随着施用量的增加呈先升后降的趋势(郑元等, 2016; 罗杰等, 2017; 王益明等, 2018; 胡倩等, 2018; Kuaietal., 2020)。但也有研究表明,硝酸还原酶和可溶性蛋白含量随施氮量的增加而增加(阚学飞等, 2010),施氮肥不能提高叶片的叶绿素含量、蒸腾速率、气孔导度和胞间 CO2浓度(郑元等, 2016; 施福军等, 2020; 马宗桓等, 2020)。磷素是磷酸盐和磷脂的组分,影响叶绿体光合磷酸化过程,控制植物能量代谢(陈隆升等, 2010)。施磷肥可提高光合速率、叶片胞间 CO2浓度和蒸腾速率,而适量的磷肥还可以降低叶片的丙二醛含量(王东光等, 2016; Gaoetal., 2020)。但也有研究认为,施磷没有影响净光合速率(罗凡等, 2014)。钾素能有效控制细胞渗透压、氧化磷酸化和光合产物转运等生物化学进程,进而调控光合作用(陆志峰等, 2016; Wangetal., 2020)。合理施钾肥可通过促进植物光合作用,增加叶片叶绿素含量、可溶性蛋白含量、净光合速、气孔导度、蒸腾速率以及水分利用效率(姚任科等, 2016; Guetal., 2021),但施钾肥显著降低水分利用效率(罗凡等, 2015)。除了氮磷钾施肥量外,不同的施肥方式显著改变植株的氮磷钾吸收,改变植物的光合作用和生理生化代谢,影响植株光合碳固定等生理活动(罗凡等, 2014; 贺维等, 2014; 苏兰茜等, 2020)。目前,不同施肥条件下如何通过改变植株养分吸收量有机地调控植物叶片光合特性、内含物含量及酶活性的机制以及它们与碳固定之间的关系尚不清楚。
闽楠(Phoebebournei)属樟科(Lauraceae)楠属,亚热带常绿阔叶树种,为中国特有的珍贵用材树种(刘宝等, 2014),而发展栽培人工林,亟需壮苗(楚秀丽等, 2014)。施肥是促进闽楠苗木优质和高效繁育的重要途径(李彬等, 2019; 施福军等, 2020)。闽楠光合作用除遗传基因外,还受到光、CO2、O3、水分、盐胁迫和肥料等因素影响(韩文军等, 2003; 王振兴等, 2012; 于浩等, 2018; 贾剑波等, 2019),目前,施肥对闽楠光合作用的影响研究主要集中在氮肥上(李彬等, 2019; 施福军等, 2020),而磷肥、钾肥和氮磷钾配合施肥对闽楠光合作用的影响,及调控光合产物形成、转化和酶活性,从而影响植株生长的研究未见报道。鉴于此,本研究分析不同氮、磷和钾施肥处理对闽楠幼苗光合色素含量、光合作用参数、叶片内含物及酶活性的影响,探讨氮、磷和钾施肥如何偶合调控闽楠光合碳固定,为闽楠壮苗培育和科学施肥提供理论依据。
1 材料与方法
1.1 试验地概况
试验地位于福建农林大学科技园田间苗圃(119°23′E,26°09′N),属于亚热带海洋性气候,极端气温最高42.3 ℃,最低-2.5 ℃,年平均温度19.6 ℃。降水量1 350 mm,年平均湿度约77%。平均日照数为1 848 h,≥10 ℃的有效积温 5 880 ℃,无霜期326天。气候温和,雨量充沛。
1.2 试验设计
试验采用“3414”最优回归设计肥效试验方案。设氮、磷、钾3个因素; 4个施肥水平(0,1,2,3),0水平为不施肥; 1水平为2水平的0.5倍; 2水平为试验地常用施肥量; 3水平为2水平的1.5倍,共14个处理。每个处理施肥量如表1(以N、P2O5、K2O计算)。试验地常用施肥量为每株0.532 g(N),0.133 g(P2O5),0.356 g(K2O)。每个处理设3 次重复,每次重复设30株幼苗。试验采用随机区组排列。供试材料为1年生闽楠实生苗。2018 年3 月,选取生长良好、长势均匀的闽楠幼苗,栽植于25 cm×35 cm塑料盆中,每盆栽植1株。栽培基质为V(红壤)∶V(蛭石)∶V(沙子)=6∶2∶2的混合基质,红壤成土母质主要为花岗岩,每盆中含有机质含量为23.12 g,pH值为5.30,全N 含量为6.0 g,全P 含量为0.14 g,全K含量为135.64 g。供试肥料为尿素(N,47%),过磷酸钙(P2O5,12%),氯化钾(K2O,60%)。试验中磷肥作为基肥结合上盆一次性施入。氮肥和钾肥按照闽楠年生长特点,分阶段施入不同的肥料量。4月上旬施入氮肥25%、钾肥20%; 6月上旬施入氮肥35%、钾肥25%; 8月上旬施入氮肥25%、钾肥35%; 10月上旬施入氮肥15%、钾肥20% (杨志坚等, 2021)。采取常规水分管理,保持田间持水量75%左右。
1.3 指标测定
1.3.1 光合生理指标测定 于2018年10月中旬晴天上午9: 00—10: 00期间进行光合生理指标测定。每个试验处理选择3株标准苗,每株选取当年生光照良好、方向相同、无病虫害、长势较一致的新梢顶芽下第3片成熟功能叶片3片,采用Li-6400便携式光合测定仪测定净光合速率(net photosynthetic rate,Pn),蒸腾速率(transpiration rate, Tr),气孔导度(stomatal conductance,gs),胞间CO2浓度(intercellular CO2concentration,Ci),水分利用效率(water use efficiency, WUE)为净光合速率(Pn)/蒸腾速率(Tr)的值,每个指标测3次。测定时饱和光强设为1 200 μmol·m-2s-1,空气流速为 500 μmol·s-1,温度控制在(25±1)℃,CO2浓度为400 μmol·mol-1(徐晓迪等, 2017)。在每次叶片光合参数测定后,采集叶片,迅速带回实验室用液氮处理,除去叶脉将剩余剪为 2 mm 左右细丝,混匀供测定光合色素、内含物含量及相关酶活性。
1.3.2 叶片内含物含量测定 可溶性糖和淀粉含量采用蒽酮比色法测定; 可溶性蛋白含量采用考马斯亮蓝G-250 法测定(向芬等, 2019); 丙二醛(MDA)含量采用硫代巴比妥酸法测定(刘连涛等, 2007)。
1.3.3 酶活性测定 多酚氧化酶(polyphenol oxidase,PPO)活性测定邻苯二酚比色法(逯红栋等, 2006); 谷氨酰胺合成酶(glutamine synthetase,GS)活性测定采用铁络合比色法; 硝酸还原酶(nitrate reductase, NR)活性采用活体法(王月福等, 2003); 酸性磷酸酶(acid phosphatase,ACP)活性参照林启美等(1991)的测定方法。
1.3.4 植株C增长量及N、P和K元素吸收量测定 2018年3月(施肥开始时),从所有参试植株中选15株长势均匀、健康完好、无病虫害的植株作为标准株。2018年12 月(施肥结束后),在每个处理90株参试植株中选取9株标准株。对标准株进行完株取样,装入干净塑料袋,带回试验室,洗净,阴干。样品在105 ℃条件下杀青15 min,并于75 ℃条件下烘干至恒质量,用电子天平(Metiler Toledo, AL204)称植株的干物质量。用植物粉碎机分别将植株磨碎后过0.5 mm 塑料筛制成分析样品,装入自封袋内用于测定全氮、全磷和全钾的含量,各处理重复3次。称取1.00 g样品,用H2SO4-HClO4消煮法制备待测液,用于N、P和K含量的测定,采用凯氏定氮法测定N含量(Shanghai Hongji,ATN-300),采用钼锑抗比色法测定P含量(Unico,UV-2600A ),采用原子吸收分光光度计法测定K含量(Beijing Dongxi,AA7002)(罗欢等, 2014)。植株C增量(N、P和K元素)(mg·plant-1) = 试验结束时植株生物量×C含量(N、P和K元素)- 试验开始时植株生物量×C含量(N、P和K元素),结果见表2。

表2 闽楠幼苗N、P、K和C元素的增量①Tab.2 Increment of N, P, K and C element in Phoebe bournei seedlings under N, P and K fertilization
1.4 数据处理
“3414”试验共14个处理,分析氮磷钾肥单因素作用,双因素互作及三因素互作。单因素作用分析,在磷钾肥固定“2”水平时,设T2-3、6和11分别为施氮水平的0、1、2、3水平; 在氮钾肥固定“2”水平时,设T4-7分别为施磷水平的0、1、2、3水平; 在氮磷肥固定“2”水平时,设T8-9、6、10分别为施钾水平的0、1、2、3水平。双因素互作分析,在钾肥固定“2”水平时,氮磷互作试验设T2、T3、T4、T5、T6、T7、T11和T12; 在磷肥固定“2”水平时,氮钾互作试验设T2、T3、T6、T8、T9、T10、T11和T13; 在氮肥固定“2”水平时,磷钾互作试验设T4、T5、T6、T7、T8、T9、T10和T14。T1-14为氮磷钾三因素互作试验。用Excel 2013软件进行数据处理,采用SPSS 16.0进行差异显著性检验(LSD法,显著性水平设为0.05)、相关分析、通径分析及极差分析。用Excel 2013软件作图。
2 结果与分析
2.1 施肥对闽楠幼苗叶片光合色素的影响
不同N、P、K施肥量显著影响闽楠幼苗叶片光合色素(P<0.05)(图1)。所有施肥处理的叶绿素a、叶绿素b、类胡萝卜素、叶绿素总含量及叶绿素a/b均大于T1(对照),T10处理叶绿素a、叶绿素b、类胡萝卜素、叶绿素总含量均最高,T11处量叶绿素a/b比值最大,说明N、P、K施肥有利于提高叶绿素a、类胡萝卜素和叶绿素总含量,且在高钾施肥水平促进作用最大,在高氮施肥水平有利于提高叶绿素a/b比值。叶绿素b含量只有T11小于T1,其余均大于T1,T3处理含量最高,说明除高氮施肥水平抑制叶绿素b含量外,其余氮磷钾施肥均促进叶绿素b含量提高。通过极差分析可知,氮磷钾不同施肥方式显著影响叶片光合色素含量(表3),氮磷钾配施极差最大,其次为氮钾配施。
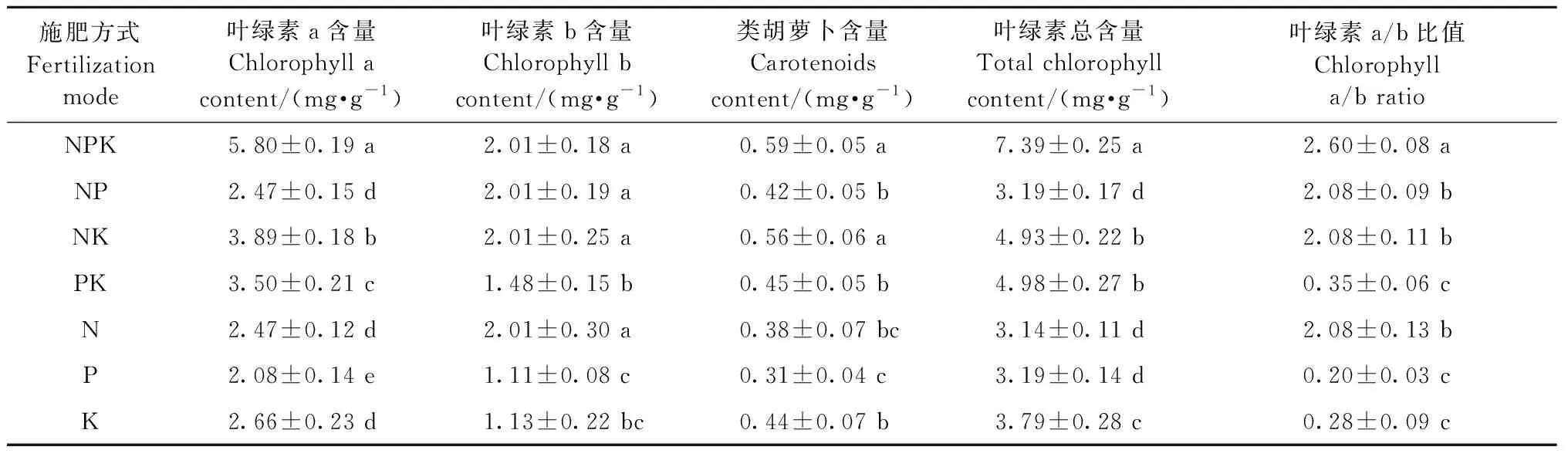
表3 氮磷钾施肥方式对闽楠幼苗叶片光合色素影响的分析①Tab.3 Analysis of the effect of NPK fertilization modes on photosynthetic pigment in leaves of P. bournei seedlings
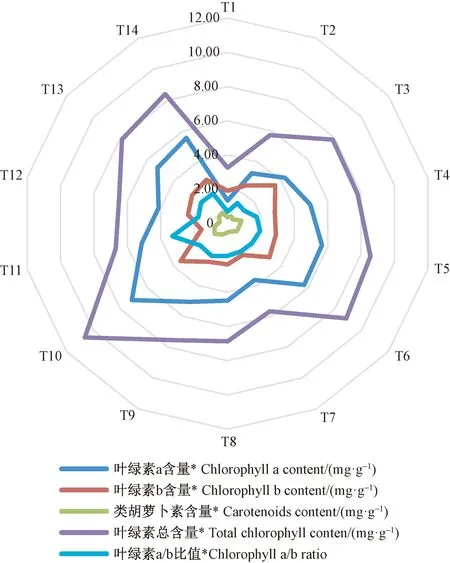
图1 不同氮磷钾施肥处理对闽楠幼苗叶片光合色素含量的影响Fig. 1 Effects of different N, P and K fertilization treatments on photosynthetic pigments content in leaves of P. bournei seedlings*表示同一指标不同氮磷钾施肥处理间在0.05水平上差异显著。下同。* on the legend indicate significant difference at 0.05 level among N, P and K fertilization treatments in the same index. The same below.
2.2 施肥对闽楠幼苗叶片光合作用的影响
进入叶片胞间CO2是光合作用的底物,直接反映光合作用的大小; 适当的蒸腾速率有利于植物吸收水分,保持固有的水分供细胞利用,叶片胞间的CO2和蒸腾作用中的 H2O吸收利用,均可受施肥等外界环境影响(张毅龙等, 2014; De Souza Kulmannetal., 2021)。本研究中,不同氮磷钾施肥处理显著影响闽楠幼苗光合作用(P<0.05)(图2)。所有处理的蒸腾速率(Tr),净光合速率(Pn),水分利用效率(WUE)和叶片胞间CO2浓度(Ci)气孔导度(gs)均大于T1(对照),说明氮磷钾施肥提高闽楠幼苗的光合、蒸腾和水分利用能力。其中,T4处理Tr最低,WUE和Ci最高; T6处理Pn最高,Ci最低; T10处理Tr最高,T13处理Ci最高,说明中水平的氮磷钾施肥最有利于提高净光合速率,无磷施肥最有利于水分利用和提高胞间CO2浓度,但不利于蒸腾作用; 而高水平施钾却最有利于蒸腾作用。通过极差分析可知,氮磷钾不同施肥方式显著影响光合作用指标(表4)。在所有光合作用指标中,极差最大的为氮磷钾配施; 在Ci、Tr和WUE指标中为极差值排在第二磷钾配施,在gs和Pn指标中为氮钾配施; 极差最小的为,在Ci和WUE指标中为单施钾肥,在Tr、gs和Pn指标中为单施磷肥。

图2 不同氮磷钾施肥处理对闽楠幼苗叶片光合作用的影响Fig. 2 Effects of different N, P and K fertilization treatments on photosynthesis in leaves of Phoebe bournei seedlings

表4 不同氮磷钾配施方式对闽楠幼苗叶片光合作用的影响Tab.4 Analysis of the effect of NPK fertilization modes on photosynthesis in leaves of P. bournei seedlings
2.3 施肥对闽楠幼苗叶片内含物的影响
不同氮磷钾施肥处理显著影响闽楠叶片内含物的含量(P<0.05)(图3)。所有处理的叶片可溶性糖含量,可溶性蛋白和MDA含量均小于T1(对照),而淀粉含量均大于T1,说明氮磷钾施肥有利于叶片淀粉含量的提高,抑制了叶片可溶性糖含量,可溶性蛋白和MDA含量。T14处理可溶性糖含量最低,T8处理可溶性蛋白含量最低,T12处理MDA含量最低,T6处理淀粉含量最高。通过极差分析可知,氮磷钾不同施肥方式显著影响叶片内含物含量(表5)。在所有光合作用指标中,极差最大的为氮磷钾配施; 其次为氮磷配施; 而极差最小的,在可溶性糖、淀粉和MDA含量指标中为单施钾肥,在可溶性蛋白含量指标中为单施磷肥。

图3 不同氮磷钾施肥处理对闽楠幼苗叶片内含物含量的影响Fig. 3 Effects of different N, P and K fertilization treatments on in clusion content in leaves of P. bournei seedlings
2.4 施肥对闽楠幼苗叶片酶活性的影响
不同氮磷钾施肥处理显著影响闽楠叶片酶的活性(P<0.05)(图4)。所有处理的叶片硝酸还原酶(NR)的还原酶活性,氨酰胺合成酶(GS)活性和酸性磷酸酶(ACP)活性均大于T1(对照),而多酸氧化酶(PPO)活性均小于T1,说明氮磷钾施肥提高叶片NR、GS和ACP酶活性,抑制PPO酶活性。T5处理的NR最大,T6处理的ACP酶活性最大,T9处理GS和PPO酶活性最大,T8处理PPO酶活性最小,说明不同酶活性对不同氮磷钾施肥响应不同,低水平施P最有利于NR酶活性的提高,低水平施K最有利于GS提高,不施钾PPO酶活性抑制最大,对中等施氮磷钾最有利于ACP酶活性提高。通过极差分析可知,氮磷钾不同施肥方式显著影响叶片酶活性(表6)。在所有光合作用指标中,极差最大的为氮磷钾配施; 极差值排在第二的,在NR、GS和ACP酶活性指标中为磷钾配施,在PPO酶活性指标中为氮磷配施; 极差最小的,在NR和ACP酶活性指标中为单施钾肥,在NR和GS酶活性指标中为单施氮肥,在PPO酶活性指标中为单施磷肥。
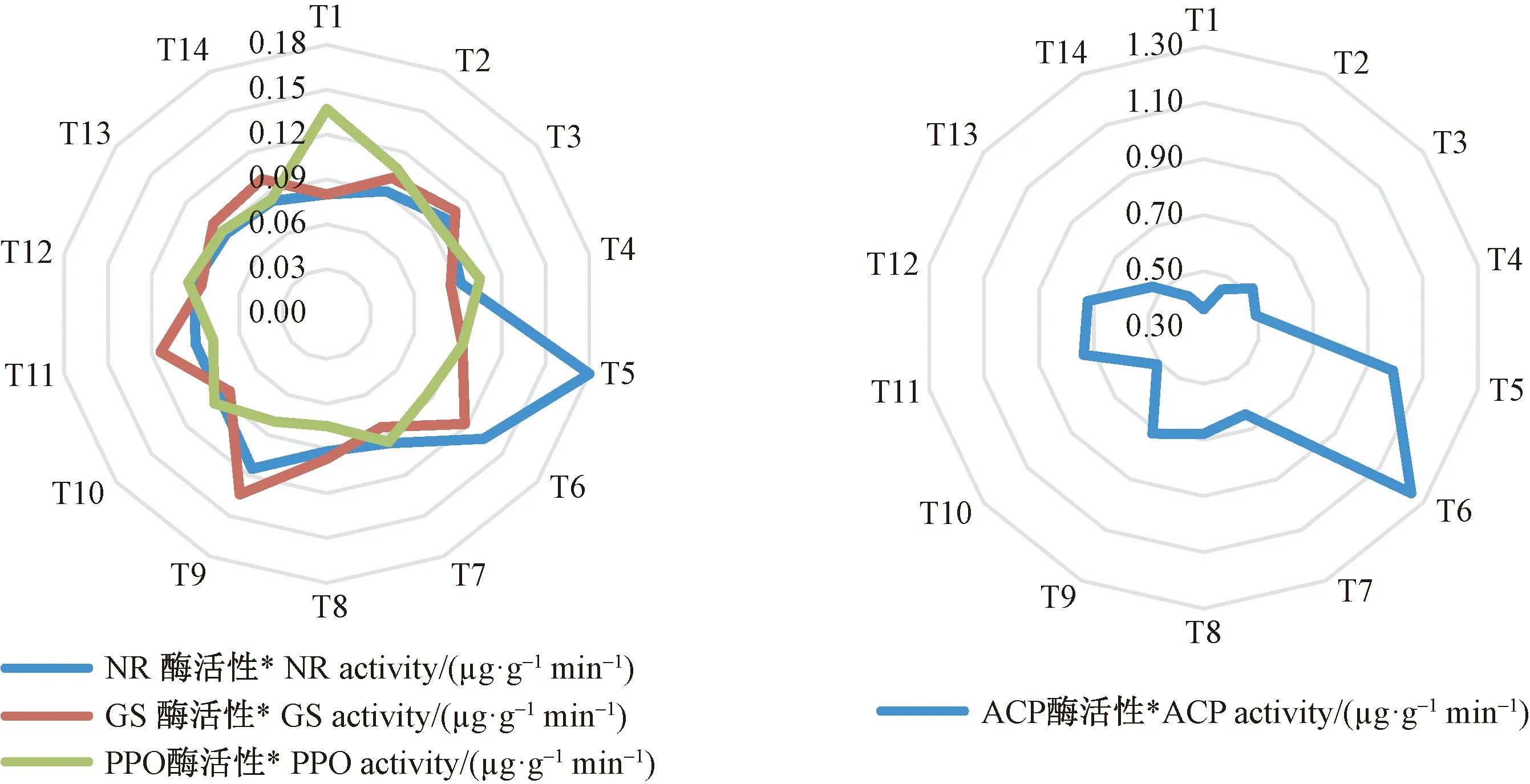
图4 不同氮磷钾施肥处理对闽楠幼苗叶片酶活性的影响Fig. 4 Effects of different N, P and K fertilization treatments on enzyme activity in leaves of P. bournei seedlings
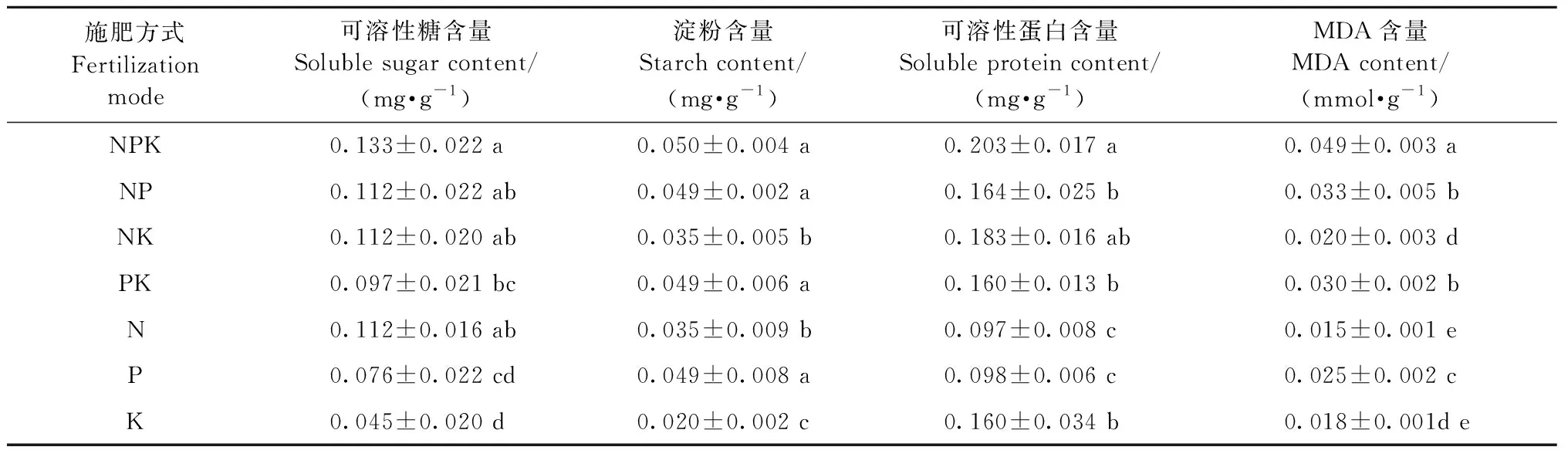
表5 氮磷钾配施方式对闽楠幼苗叶片内含物含量的影响Tab.5 Analysis of the effect of NPK fertilization modes on inclusion content in leaves of P. bournei seedlings

表6 氮磷钾配施方式对闽楠幼苗叶片酶活性的影响Tab.6 Analysis of the effect of NPK fertilization modes on enzyme activity in leaves of P. bournei seedlings
2.5 闽楠幼苗叶片各指标与植株N,P,K吸收量的相关性
通过对不同施肥方式下植株N、P和K元素的吸收量和叶片色素含量、光合指标、内含物含量及酶活性各指标之间相关关系分析,判断植株N、P和K元素的吸收与叶片各指标的影响程度,如表7所示。N增量与叶绿素a含量、叶绿素总含量、Pn、WUE、ACP酶活性、NR酶活性和C增量呈极显著正相关(P<0.01),与叶绿素b含量、类胡萝卜素含量、叶绿素a/b比值、Tr、gs、淀粉含量、GS酶活性呈显著正相关(P<0.05),与可溶性糖、可溶性蛋白和MDA呈负相关。P增量与gs、Pn、ACP酶活性、PPO酶活性和C增量呈极显著正相关(P<0.01),与叶绿素b含量、WUE、NR酶活性呈显著正相关(P<0.05)。K增量与叶绿素a含量、叶绿素b含量、叶绿素总含量、Tr、Pn、淀粉含量、ACP酶活性、NR酶活性和C增量呈极显著正相关(P<0.01),与WUE呈显著正相关(P<0.05)。

表7 闽楠幼苗叶片各指标与植株N,P,K吸收量的相关分析①Tab.7 Correlation analysis of the N, P, K uptake of the plant and the indexes of leaves in P. bournei seedlings
2.6 闽楠幼苗光合碳固定主导影响因素
通过逐步回归分析,筛选出影响闽楠幼苗光合碳固定的主导因素。建立了不同施肥方式下N,P和K吸收量、叶片色素含量、光合指标、内含物含量及酶活性共21个指标,对闽楠幼苗C增量的逐步回归方程,得出植株C增量=103.483+10.584·X1-23.067·X2-0.504·X3-131.159·X4+18.697·X5+1 644.908·X6-144.948·X7+161.728·X8-578.593·X9(P<0.05,R=0.956), 其中X1为叶绿素总量,X2为叶绿素a/b比值,X3为Ci,X4为Tr,X5为WUE,X6为淀粉含量,X7为可溶性蛋白含量,X8为ACP酶活性,X9为NR酶活性。由此可得,叶绿素总量、叶绿素a/b比值、Ci、Tr、WUE、淀粉含量、可溶性蛋白含量、ACP和NR酶活性是施肥影响闽楠幼苗C增量的主要因素。
从逐步回归分析中筛选到的9个主要因素指标进一步与闽楠幼苗C增量作通径分析得出(决定系数为0.917,剩余通径系数为0.353)(表8),叶绿素总量、WUE、淀粉含量和ACP酶活性的直接通径系数均为正值,对C增长有促进作用,最大为ACP酶活性,其次是淀粉含量,最小的为叶绿素总量。而叶绿素a/b含量、Ci、Tr、可溶性蛋白含量和NR酶活性的直接通径系数为负值,对C增长有抑制作用,最大为Ci,其次为Tr,最小为NR酶活性。植株C增长与叶绿素总量、WUE、淀粉含量和NR酶活性呈显著正相关(P<0.05),与ACP酶活性呈极显著正相关(P<0.01)。叶绿素总量通过WUE对植株C增量的间接通径系数最大,叶绿素a/b比值通过ACP酶活性对植株C增量的间接通径系数最大,Ci通过WUE对植株C增量的间接通径系数最大,Tr通过淀粉含量和WUE对植株C增量的间接通径系数最大,WUE通过Ci对植株C增量的间接通径系数最大,淀粉含量通过ACP酶活性对植株C增量的间接通径系数最大,可溶性蛋白含量通过淀粉含量对植株C增量的间接通径系数最大,ACP酶活性通过淀粉含量和NR酶活性对植株C增量的间接通径系数最大,NR酶活性通过WUE和ACP酶活性对植株C增量的间接通径系数最大。

表8 闽楠幼苗植株C增量的通径分析Tab.8 Path analysis of plant C increment in P. bournei seedlings
3 讨论
3.1 氮、磷、钾施肥促进闽楠光合色素合成
叶绿素是光合作用的光敏催化剂,其含量和比例变化可反映植物适应和利用环境因子的能力(Xiongetal., 2015; 周佳民等, 2019)。闽楠幼苗N增量与叶片光合色素均呈显著正相关,与叶绿素a含量和叶绿素总量呈极显著正相关,在高氮施肥水平有利于提高叶绿素a/b比值,说明施氮肥可以显著提高闽楠幼苗光合色素含量,特别是叶绿素 a,可能由于闽楠幼苗属阴生植物(刘宝等, 2014),氮作为叶绿素主要组成元素,施氮后叶片提高叶绿素a含量可以捕捉更多红光(许改平等, 2014),提高叶绿素b可以充分吸收蓝紫光,提高类胡萝卜素含量保护叶绿素不受光氧化损害(Cheminantetal., 2011; 刘建福等, 2014),但叶绿素 a 在闽楠幼苗叶片利用光能时却起关键性作用(刘宝等, 2014),这与施福军等(2020)的研究结果相同。P吸收量只与叶绿素b含量呈显著正相关,说明施磷肥更利于叶绿素b含量的提高,可能施磷能更有效地促进光化学量子产额的增加(王阳等, 2021),为叶绿a向叶绿素b转化提供能量(王平荣等, 2009)。K吸收量与叶绿素a含量、叶绿素b含量和叶绿素总量呈极显著正相关,且高水平施钾对闽楠的光合色素促进作用最大,可能施钾可以激化闽楠幼苗体内多种酶的活性,为叶绿素a和b合成提供辅酶(杜俊卿等, 2004)。由此,氮磷钾三因素配施对各种光合色素影响最大,其次为氮钾配施,最小的为单施磷肥; 并在T10(N2P2K3)处理中,各光合色素含量均最高。
3.2 氮、磷、钾施肥提高闽楠叶片光合作用
N增量与Pn、WUE呈极显著正相关,与Tr和gs呈显著正相关,可能闽楠施氮后,加大气孔导度,既提高CO2的固定量,也增大了蒸腾速率,但降低了单位碳同化的水分耗散,提高水分利用效率(张毅龙等, 2014),说明施氮是通过提高水分利用效率来提高闽楠的光合作用。P增量与gs、Pn呈极显著正相关,与WUE呈显著正相关,无磷施肥处理(T4)最有利于水分利用和提高胞间CO2浓度,但不利于蒸腾作用,可能磷素能促进光合作用的酶促反应和能量传递,提高细胞气孔CO2的利用率,增强水分利用,减少了气孔内的压力,促进叶片气体交流,从而提高光合作用(王东光等, 2016; De Souza Kulmannetal., 2021),说明施磷肥是通过提高C同化效率来增强闽楠的光合作用。K增量与Pn呈极显著正相关,与WUE呈显著正相关,高水平施钾肥最有利于蒸腾作用,单施钾肥方式中胞间CO2浓度(Ci)极差最小,这可能因为K+促使闽楠叶片气孔开放,提高CO2的吸收和蒸腾作用,从而提高光合作用(罗凡等, 2014),说明施钾肥是通过叶片气孔开放提高闽楠光合作用。因此,N、P和K肥三因素配施,才能从扩大气孔,提高水分利用效率和碳同化效率这三个环节互相配合去提高闽楠的光合能力,并在T6(N2P2K2)处理中达到最大。
3.3 氮、磷、钾施肥调控闽楠光合产物代谢
本研究中,N增量与C增量呈极显著正相关,与叶绿体总量呈极显著正相关,与淀粉含量呈显著正相关,与可溶性糖、可溶性蛋白和MDA含量呈负相关。可能闽楠吸收N后,促进叶片光合作用,产生可溶性糖,进入氮代谢,合成蛋白,减少叶片可溶性糖含量(施福军等, 2020),新产生的蛋白一部分,保护了细胞膜的完整性,降低MDA含量,延缓叶片衰老(王渭玲等, 2003; Namvaretal., 2015);另一部分提高了叶绿体中的类囊体蛋白,减少叶片中可溶性蛋白含量(罗杰等, 2017),促进叶片光合作用,产生更多可溶性糖,进而促进碳代谢,提高淀粉含量,说明闽楠施氮是通过增强氮代谢来提高碳固定。P增量与C增量呈极显著正相关,与叶片的内含物成分无显著相关,说明P仅显著影响闽楠叶片的光合同化作用,而对于光合产物的代谢无显著影响。K吸收量与淀粉和植株C增长量呈极显著正相关,可能闽楠吸收K后,提高相应酶活性,增强叶片光合作用,产物增多,促进细胞碳代谢,提高淀粉含量,为苗木生长储存能量(Geigenberger, 2011; 李亚姝等, 2020)。
酰胺合成酶(GS)是参与氨同化过程的关键酶(向芬等, 2019),硝酸还原酶(NR)是植物体内硝态氮同化的调节酶和限速酶, 这2种酶共同调节氮代谢,影响光合碳代谢(唐晓清等, 2013; Zhangetal., 2017)。氮磷钾施肥提高闽楠叶片GS和NR酶活性,N、P和K吸收量均与NR酶活性呈极显著相关,N和P吸收量与GS酶活性呈显著相关性,氮肥、磷肥和钾肥两因素配施中,GS和NR酶活性在磷钾肥配施影响最大; 单因素配施中,单施氮肥影响最小。说明氮磷钾施肥可加强闽楠幼苗叶片氮代谢,其中硝态氮代谢强于氨态氮代谢,与甘薯(Dioscoreaesculent)的研究相同(汪顺义等, 2016),但施氮肥对氮代谢的影响最小。酸性磷酸酶(ACP)是一种水解酶,在植物碳水化合物的转化及蛋白质合成中起重要作用(刘攀道等, 2019; 代庭伟等, 2020)。氮磷钾施肥提高闽楠叶片ACP活性,N、P和K吸收量均与ACP呈极显著相关,说明闽楠幼苗施肥后提高碳水化合物的转化及蛋白质的合成。多酚氧化酶(PPO)在是植物酚类物质合成的关键酶,为植物防卫蛋白(马长青等, 2013; Fuerstetal., 2014)。氮磷钾施肥降低闽楠叶片的PPO活性,N、P和K吸收量均与PPO无显著相关关系,说明闽楠幼苗氮磷钾施肥后,降低了酚类物质合成,N,P和K吸收提高了闽楠幼苗环境适应性,促进生长。由此,不同氮、磷、钾施肥可极显著的影响叶片内含物含量和各种酶活性,T6(N2P2K2)处理效果最好,氮磷钾三因素施肥影响最大,其次为双因素施肥,最小为单因素施肥。
3.4 氮、磷、钾施肥调控闽楠光合碳固定
为明确氮磷钾施肥调控光合生理的机制,将所有的光合生理指标与C增长量进行逐步回归分析,并进一步作通径分析,得出N、P、K施肥调控闽楠叶片生理活动和光合作用,促进C固定。施肥后,当闽楠幼苗吸收N,NR酶活性增强,促进氮代谢,细胞内蛋白质增多,一方面促使叶绿体内囊体蛋白增多,提高叶绿体含量,另一方面ACP酶活性增强,在内囊体蛋白和ACP酶的共同作用下,闽楠叶片叶绿体a/b比值增大,使闽楠幼苗叶片的光合效能增强。当闽楠幼苗吸收P,叶片光合磷酸化作用增强,细胞能量ATP增多,进一步促进蛋白质合成,细胞膜蛋白增多,降低了丙二醛含量,保护了细胞的完整性,使细胞光合能力更强。当闽楠幼苗吸收K,开放叶片气孔,气孔导度增大,提高气孔CO2和水分利用效率,增强叶片净光合能力。闽楠幼苗N、P和K吸收量增大后,均增强了光合能力,提高光合产物(可溶性糖)的输出,在ACP酶的作用下,进入氮和碳代谢,产生淀粉,为其生长提供物质和能量。同时当叶片中淀粉含量达到一定值后,淀粉通过负反馈ACP酶和WUE,降低叶片的光合作用,从而使闽楠光合产物产出和利用达到平衡。
4 结论
N、P、K配施后,闽楠叶片的对光合色素含量、水分利用效率、光合磷酸化、碳同化效率、叶片内含物含量和相应酶活性影响最大,其次为氮钾配施,最小的为单施磷肥,T6(N2P2K2)处理效果最好,进一步分析作用机制认为:1) 施NPK肥后,提高了闽楠叶片的光合色素含量,特别是叶绿素a,进而提高了闽楠幼苗的光合、蒸腾和水分利用能力,利于叶片淀粉含量的提高,抑制叶片可溶性糖含量,可溶性蛋白和MDA含量,提高叶片NR、GS和ACP酶活性,抑制PPO酶活性。2) N、P和K肥对光合碳固定作用不同。施氮肥后,NR酶活性增强,促进氮代谢,主要提高叶绿素 a,增强水分利用效率,提高光合作用,提高淀粉含量,促进光合碳固定。 3) 施磷肥则主要提高叶绿素b含量,增强叶片光合磷酸化作用,降低了MDA含量,保护细胞完整性,提高C同化效率,但对于光合产物代谢无显著影响。 4) 施钾肥可同时提高叶绿素a和b含量,增大气孔导度,提高气孔CO2和水分利用效率,增强叶片光合作用,促进光合碳固定。 5) N、P和K元素相互协调,相互制约的影响着光合碳固定,其中叶片ACP酶、WUE和淀粉含量是施肥调控光合碳固定的关键点。
猜你喜欢
今日农业(2022年14期)2022-11-10
北京航空航天大学学报(2022年5期)2022-06-06
财会月刊·下半月(2022年4期)2022-04-25
国际商业技术(2022年4期)2022-04-21
当代陕西(2022年6期)2022-04-19
农业工程学报(2022年1期)2022-03-25
河南科技(2022年1期)2022-03-11
妇女生活(2019年1期)2019-01-17
江苏农业科学(2017年19期)2017-11-22
农家顾问(2016年7期)2016-05-14
