
益生菌对宿主肠道屏障功能的影响
2022-08-18 03:58王超越韩瑨吴正钧郭诚笑唐月方
食品与发酵工业 2022年15期
王超越,韩瑨,吴正钧*,郭诚笑,唐月方
1(光明乳业股份有限公司乳业研究院,上海乳业生物工程技术研究中心,乳业生物技术国家重点实验室,上海,200436) 2(上海海洋大学 食品学院,上海,201306)
消化道是微生物和宿主免疫系统相互作用的主要场所,在抵御病原体的同时允许固有微生物生存。肠黏膜具有约300 m2的表面积,是人体与外界环境接触面积最大的结构。肠黏膜屏障的完整性是维持消化道稳态的关键,完整的肠黏膜屏障能够保护机体免受肠道细菌、内毒素及抗原的侵害,保证机体健康。肠道通透性增高(通常称为“肠漏”),会造成肠腔内容物移位,进入机体循环,促使机体释放大量炎性细胞因子,诱发异常的黏膜免疫反应[1]。肠道屏障损伤与多种疾病有关,包括胃肠疾病,如乳糜泻 (celiac disease, CeD)、炎症性肠病(inflammatory bowel disease, IBD)、结直肠癌(colorectal cancer, CRC)等和肠外疾病(如慢性肝病、Ⅰ型糖尿病、肥胖)[2]。因此,维持肠黏膜屏障功能的完整性将为临床疾病的干预提供新思路。
益生菌被定义为:当使用一定量后,对宿主健康产生有益作用的活的微生物。益生菌可作用的部位包括肠道、上呼吸道、泌尿生殖道和皮肤。在食品中常用的益生菌包括双歧杆菌、乳杆菌(如嗜酸乳杆菌、植物乳杆菌)以及部分酵母菌[如布拉氏酵母菌(Saccharomycesboulardii)]等。在人体中,益生菌可通过调节宿主肠道微生态发挥改善精神健康、降低胆固醇、改善胰岛素敏感性、调节免疫、预防过敏等作用[3-5]。作为一类具有良好应用潜力的生物治疗剂,益生菌通过对肠道屏障的调节作用,极大地影响着宿主的健康状态及疾病的发生和发展,其作用机制也不断被揭示。
1 肠道屏障
肠道屏障是一个由机械、化学、免疫和菌群屏障组成的动态系统(图1)。4个屏障共同作用,维持消化道功能的稳态,保护机体免受肠道致病菌、内毒素及抗原的侵害,从而维持机体健康。
1.1 机械屏障
机械屏障主要由定殖在隐窝底部的肠道干细胞分化而成的各种肠上皮细胞及细胞间连接复合体构成。
相邻肠上皮细胞间隙由紧密连接(tight junction, TJ)、黏附连接、桥粒和缝隙连接等4种连接复合体进行封闭,形成紧密的具有动态通透性屏障功能的柱状细胞单层,阻止肠腔中微生物细胞、菌体碎片及内毒素等有害物质透过肠道黏膜进入血液循环,同时允许营养物质和水进入体内,是抵抗感染和炎症的主要防御机制。
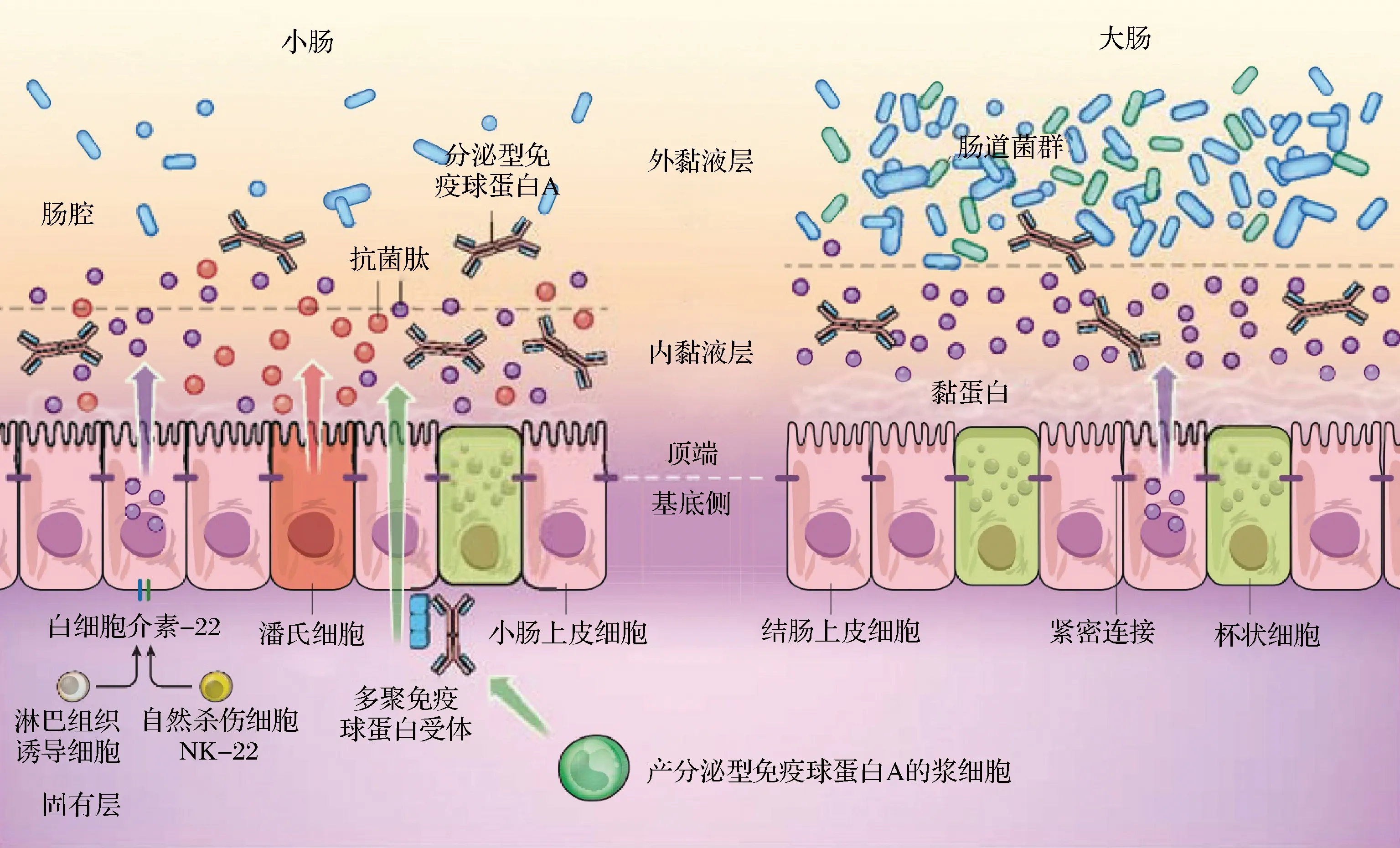
图1 肠道屏障组成示意图[6]Fig.1 Schematic diagram of intestinal barrier composition[6]
肠道机械屏障还包括肠蠕动功能,进入肠道的食物随着肠道节律性蠕动进行下移,将肠细胞表面一些致病性细菌一起带走,减少了有毒有害细菌在肠黏膜表面的停留时间,防止细菌易位引起的炎症性疾病。
1.2 化学屏障
化学屏障包括消化液、溶菌酶、黏多糖、其他化学物质和肠道分泌的抗菌物质[7]。消化道黏液主要由杯状细胞分泌到管腔中的高度糖基化的黏蛋白组成,还包括碳水化合物、脂类和水。消化道黏液保护上皮细胞的方式有以下几点:形成将上皮细胞和肠腔环境隔开的水溶性黏液层;黏液中的碳水化合物和细菌进行黏附,阻止细菌与上皮细胞接触;黏液中存在补体成分、防御素和乳铁蛋白等能够直接杀伤微生物的活性成分;黏液在随管壁平滑肌运动而排出体外时将细菌等有害物质一起排出。另外,肠道中还存在各种具有杀菌和溶菌作用的蛋白、溶菌酶、胆汁等成分。
1.3 菌群屏障
正常情况下,肠道内的益生菌和致病菌相互依赖相互制约达到一种平衡状态,构成肠黏膜的菌群屏障。肠道中比例最高的专性厌氧菌通过以下几点保护肠道健康:附着在肠道黏膜表面, 通过定殖拮抗作用抑制病原菌的黏附,具有抗菌和调节肠道免疫反应的作用;分泌乳酸和短链脂肪酸(short chain fatty acid,SCFA)等次级代谢物,降低机体肠道pH和氧化还原电位,阻止不耐酸的腐败菌和致病菌的生长;分泌细菌素抑制肠道兼性厌氧菌和外来菌的定殖和生长;作为肠道中优势菌群与致病菌争夺养料,抑制致病菌的生长。
1.4 免疫屏障
免疫系统由免疫器官、免疫细胞、可溶性细胞因子和细胞受体调节。黏膜上皮内淋巴细胞CD8+具有细胞毒性作用,被病原体激活后可释放穿孔素、端粒酶和丝氨酸醋酶等细胞因子,从而杀伤肠道中的病原菌,防止病原菌的入侵。另外,吞噬细胞一方面通过吞噬作用抵抗并消除一部分病原体,另一方面将病原微生物的信息传递给B淋巴细胞,B淋巴细胞进行增殖并转化为浆细胞,进而产生分泌型免疫球蛋白A (secretory immunoglobulin A, sIgA),sIgA又通过使病原微生物发生凝集或与病原微生物结合阻断其表面的特异性结合位点,使其丧失黏附功能。共生微生物群启动肠道先天性免疫和适应性免疫,可协调保护宿主免受外来病原体入侵,同时维持肠道内环境稳定[8-9]。
2 益生菌对肠道屏障功能的调节作用
2.1 促进上皮细胞增殖分化
动物出生后,隐窝中的干细胞随着微生物的定殖逐渐成熟,分化为各种具有特殊功能的肠上皮细胞。肠上皮细胞通过增殖、分化和凋亡不断地进行着更替,维持上皮的稳态和再生。其中,潘氏细胞每18~23 d更新一次,其他肠上皮细胞每3~5 d更新一次。研究发现,罗伊氏乳杆菌D8的代谢物吲哚-3-甲醛通过芳香烃受体刺激固有层淋巴细胞分泌白细胞介素22,诱导STAT3磷酸化,从而加速肠上皮细胞的增殖[10]。另一项研究表明,罗伊氏乳杆菌能够通过增加 R-spondins 的表达从而激活Wnt/β-catenin通路达到刺激肠上皮细胞增殖的目的[11]。
2.2 降低肠道屏障的通透性
在正常状态下,肠道屏障的透过性具有选择性,允许必需营养素和水通过,限制毒素和病原体的透过。当肠道屏障功能受损,出现通透性增加时,肠腔内有害细菌的细胞成分及其代谢产物进入宿主循环系统后,可通过刺激宿主过度的炎症反应导致消化道和自身免疫性疾病。除了促进上皮细胞增殖分化外,益生菌还可以通过上调宿主肠上皮细胞对TJ蛋白的表达,从而有效维持黏膜完整性。YI等[12]将断奶仔猪分为对照组、正常饮食加罗伊氏乳杆菌LR1和抗生素处理3个组。结果发现,与抗生素组和对照组相比,益生菌组仔猪肠上皮绒毛高度与隐窝深度的比值升高,空肠和回肠黏膜TJ蛋白表达升高。在另一项研究中,给腹腔注射脂多糖(lipopolysaccharide, LPS)诱导屏障功能损伤的小鼠灌胃罗伊氏乳杆菌ZJ617和鼠李糖乳杆菌GG(LactobacillusrhamnosusGG, LGG),发现ZJ617和LGG均能显著逆转LPS诱导的occludin和claudin-3表达下降[13]。从动物实验中观察到的益生菌维持屏障完整性的结果,为进行临床研究提供了可能性[14-15]。
2.3 促进黏蛋白基因的表达和分泌
黏液层的厚度通常可达到50~800 μm, 在健康个体中,贴近上皮层30 μm厚度区域内的黏液几乎不存在细菌。黏液是肠腔内细菌遇到的第一道屏障,病原菌需要穿透黏液层才有可能引发感染。致病菌降解黏液层的方式多种多样,包括减少黏蛋白的二硫键数量、分泌蛋白酶、分泌糖苷键水解酶(口腔及部分肠道菌群)等。黏液中发挥阻断肠腔内容物与肠上皮层接触作用的主要是黏蛋白。黏蛋白是一类大分子、高度糖基化的蛋白(碳水化合物含量超过80%),糖基部分将蛋白部分包裹在内部,防止其被机体分泌的蛋白酶水解。在小肠中,黏蛋白主要为MUC2。
研究发现,鼠李糖乳杆菌CNCM I-3690能通过增加杯状细胞的数量和促进黏蛋白的表达来改善结肠屏障功能[16]。副干酪乳杆菌CBAL74发酵的脱脂乳在85 ℃下加热20 s灭活后进行喷雾干燥,得到仅含有灭活菌体的发酵产物FM-CBAL74,FM-CBAL74能以剂量依赖的方式显著地诱导Caco-2细胞的生长和分化,上调TJ蛋白和MUC2基因的表达,但研究者未明确是FM-CBAL74中的何种成分产生的作用[17]。另一项研究也表明,受沙门氏菌感染的小鼠在食用植物乳杆菌MTCC 5690发酵乳后,MUC2的表达有所增加,肠屏障功能得到改善[18]。除了食用单独的益生菌或益生菌发酵乳外,连续7天口服VSL#3益生菌混合物可通过上调肠上皮细胞中MUC2、MUC3和MUC5AC基因的表达,使大鼠基底管腔黏蛋白含量增加60%[19]。
2.4 竞争黏膜定殖位点,形成菌膜屏障
细菌表面的结构决定其在宿主肠道内的定殖能力。益生菌通过细胞表层蛋白(surface layer proteins, SLPs)和菌毛等黏附在肠道黏液上,通过抑制和排斥作用减少细菌性肠道病原体的定殖,在维持肠道内稳态和促进肠道健康方面发挥作用[20]。SLPs是覆盖在乳杆菌菌体表面的一类呈规则晶格状排列的蛋白,由蛋白质或糖蛋白亚基组成。研究发现,与仅用大肠杆菌O157:H7感染的HEp-2上皮细胞相比,使用从瑞士乳杆菌R0052分离的SLPs提取物预处理HEp-2上皮细胞可导致大肠杆菌O157:H7的黏附减少[21]。KIM等[22]从短乳杆菌FSB-1的SLPs中分离出凝集素,该凝集素能识别大鼠结肠黏蛋白的末端糖链并与之结合,从而减少致病菌的黏附。
2.5 调节肠道菌群组成
人类胃肠道拥有一个复杂而动态的微生物生态系统,由数万亿微生物组成。随着对肠道菌群组成和功能的不断认识,其对肠道健康的重要性越来越受到关注。研究发现,肠道菌群及其代谢产物能够调节先天性和适应性免疫细胞的发育、稳态和功能,抑制有害菌的生长,维持肠道上皮组织的完整性[23]。由于肠道菌群可以吸收宿主自身难以消化吸收的成分及分泌到肠腔中的物质,所以肠道菌群的组成会随着机体摄入的食物成分的改变而改变。不当的饮食会造成肠道菌群失调进而诱发多种肠道疾病。摄入添加益生菌的食品可调节肠道菌群组成,从而达到预防甚至是治疗相关疾病的目的。相关研究发现,在乙二醇摄入前补充植物乳杆菌N-1可以预防乙二醇诱导的大鼠肠道菌群失调和肠道屏障损伤[24]。另一项研究发现,瘤胃乳杆菌FXJWS27L3和FXJSW17L1处理能显著增加小鼠肠道菌群多样性,平衡肠道菌群的组成和分布,缓解葡聚糖硫酸钠诱导的结肠炎[25]。
2.6 调节肠道黏膜免疫反应
益生菌通过细菌-上皮-免疫细胞之间的互作加强先天和适应性免疫反应。肠上皮细胞和黏膜免疫细胞表面的Toll样受体(toll like receptors, TLRs)和NOD样受体可以特异性识别病原体相关分子模式,迅速感应菌群变化,激活核因子κB(nuclear factor κB, NF-κB)和丝裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)信号通路,启动宿主免疫防御反应。在正常生理条件下,肠黏膜免疫系统对固有菌处于免疫耐受状态,维持肠道微生态平衡。
肠黏膜上散布着一种叫派伊尔结的圆顶状结构,其中富含大量的免疫细胞,包括T淋巴细胞、B淋巴细胞、巨噬细胞和树突状细胞。派伊尔结是与消化道相关淋巴组织调节肠道对病原体免疫反应的关键。在一项研究中,大剂量卡那霉素扰乱小鼠肠道菌群后,小鼠血清免疫球蛋白E(immunoglobin E, IgE)水平升高,血清IgA和粪便IgA水平降低,派伊尔结细胞数量减少;而乳酸菌有效地降低卡那霉素处理后小鼠血清IgE水平,恢复血清IgA和粪便IgA水平,增加派伊尔结细胞的数量,上调了回肠组织中ZO-1和occludin的mRNA水平。表明乳酸菌可以通过改善肠道屏障的完整性和增加派伊尔结中IgA的产生来增强肠道免疫功能,调节由抗生素引发的肠屏障功能损伤[26]。
3 益生菌在肠道疾病中的应用
3.1 炎症性肠病(inflammatory bowel disease, IBD)
IBD是一种慢性炎症引起的消化道疾病,包括克罗恩病(Crohn’s disease, CD)、溃疡性结肠炎(ulcerative colitis, UC)和未确定型结肠炎(indeterminate colitis, IC),可根据炎症的位置进行区分[27]。尽管IBD的确切病因尚不清楚,通常认为是遗传易感个体对固有微生物群出现炎性反应的结果,饮食和压力被认为是IBD的潜在诱因。此外,肠道菌群组成异常引发的肠道炎性反应,造成黏膜屏障损伤,增加肠腔内容物和细菌易位,可能引发宿主持续的炎性反应。研究发现,IBD患者的肠道微生物群与健康人不同。在IBD患者中,梭状芽胞杆菌科或黏附侵袭性大肠杆菌菌株比普通人群中更常见[28-29]。
在过去几十年里,益生菌在调节菌群组成及其在治疗IBD中的效果引起了广泛的关注。益生菌可通过降低肠腔pH值、竞争营养物质、分泌和诱导宿主产生抗菌化合物(细菌素和防御素)、阻止细菌黏附于上皮细胞等途径防止病原菌的过度生长。此外,益生菌还可以通过免疫调节作用,如降低NF-κB的活化、促进树突细胞的成熟、刺激细胞因子(interleukin-10, IL-10)的合成、增强自然杀伤细胞(natural killer, NK)的杀伤活性、促进派伊尔氏淋巴集结中抗原呈递细胞的活化改善IBD。
尽管有证据表明益生菌对溃疡性结肠炎具有免疫调节作用,但大多数研究没有证明益生菌补充剂对CD的治疗作用。
3.2 结直肠癌(colorectal cancer, CRC)
Cancer Today (WHO) 目前的统计数据显示,CRC是全球第三大确诊癌症类型,2020年新发病例为1 931 590例(占癌症总病例的10%),死亡病例为935 173例(占癌症总死亡率的9.4%)。CRC的发病率在欧洲、北美和澳大利亚高于亚洲国家。病因学表明,遗传因素和环境因素共同导致CRC的发生。
益生菌可以通过一系列机制预防CRC的发生[30]。大多数情况下,这些机制组成性地发挥作用,对肠道菌群组成产生积极影响,并防止结肠和直肠受到经常诱发癌变的环境应激因素的影响。主要作用机制如下:
(1)调节肠道菌群的组成,减少致癌物的产生。肠道菌群产生的代谢酶通常将不同的前体化合物转化为致癌化合物。结肠和直肠的细胞经常暴露在不同的致癌化合物中。调节肠道菌群的酶表达,可以间接控制致癌化合物的产生。研究表明,经常食用益生菌可以改善肠道菌群的种类和丰度,从而减少肠道内致癌化合物的产生和慢性炎症的发生[31]。另外,细菌细胞壁肽聚糖可通过与致癌物物理结合或降低化合物的活性来直接灭活致癌化合物[32]。
(2)调节免疫系统。肠道菌群和肠道免疫系统的互作在维持体内平衡中起着重要的作用。上皮细胞中TLRs通过识别不同的微生物相关分子模式启动一系列免疫反应,失调的肠道微生态系统导致MAPK途径激活和NF-κB易位,从而刺激不同的促炎细胞因子如IL-8和NO的分泌,最终导致IBD和CRC的发生。益生菌通过调节抗炎细胞因子(IL-10和TGF-β2)和促炎细胞因子的产生影响机体的免疫应答。某些益生菌还可以激活吞噬细胞,使免疫反应处于一种警戒状态,在癌细胞的早期发展阶段将其清除。
(3)增强肠上皮屏障。TJ蛋白复合物的破坏或细胞旁通透性的增大通常会导致IBD的发生,并常伴有IBD相关的CRC[33]。益生菌能分泌SCFA,这些SCFA在改善肠道屏障功能方面显示出积极的效果。研究发现,SCFA如丁酸盐、丙酸盐和乙酸盐对乙醇诱导的屏障功能损伤具有保护作用[34]。乙醇处理会破坏肠上皮细胞间TJs和上皮细胞骨架并引起代谢应激的增加,而SCFAs通过激活Caco-2细胞中腺苷酸活化蛋白激酶[adenosine 5’-monophosphate (AMP)-activated protein kinase, AMPK]信号通路能够减轻代谢应激并修复TJs[35]。
(4)诱导癌细胞凋亡。益生菌促进癌细胞凋亡的作用在许多体外实验中得到了证实,通常伴有凋亡相关蛋白的表达,如死亡配体受体、caspase-3、caspase-8、caspase-9、Bax/Bak、Bcl-2和Bcl-x蛋白[36]。RABIEI等[37]研究发现,干酪乳杆菌UT1的发酵上清液以时间和剂量依赖的方式促进人结肠癌细胞HCT116的凋亡。另一项研究发现,植物乳杆菌-12产生的粗胞外多糖通过增加HT-29细胞中凋亡蛋白如Bax、细胞色素C、caspase-3、caspase-8和caspase-9酶的表达,降低抗凋亡蛋白Bcl-2的表达,诱导HT-29细胞凋亡[38]。
3.3 乳糜泻(celiac disease, CeD)
乳糜泻,又称麦胶性肠病,是一种多因素慢性炎症性小肠疾病,CeD的发生与遗传因素(HLA-DQ2和HLA-DQ8)、自身抗原(组织转谷氨酰胺酶2)和环境因素(麸质蛋白)有关。HLA-DQ2/8是一种常见的基因型,约30%~40%的人群中存在这种基因,但只有2%~5%的携带者会发展成CeD。麸质蛋白是CeD的主要环境诱因,但麸质蛋白的摄入并不能完全解释这种疾病的发生,一些人在接触麸质蛋白多年后才患上这种疾病,这表明还有其他因素在CeD的发生中起作用[39-40]。越来越多的证据表明,肠道菌群与消化道疾病密切相关[41]。肠道菌群的变化可能改变肠腔内麸质蛋白的代谢、肠道通透性和宿主免疫系统,并可能导致麸质蛋白敏感性肠病[42]。目前治疗CeD的主要方法是坚持严格的无麸质饮食(gluten free diet, GFD),虽然GFD可以减轻大多数患者的CeD症状,但并不能将患者的肠道菌群完全恢复到健康个体状态[43-44]。
益生菌可能通过以下3种机制影响CeD:
(1)通过产生能有效切割麸质蛋白肽中蛋白水解抗性序列的酶来降低麸质蛋白的免疫原性,消除或减少CeD的诱因,从而防止其发作。CAMINERO等[45]利用小鼠实验研究了麸质蛋白在条件致病菌(铜绿假单胞菌)和十二指肠共生细菌(乳酸菌)作用下的代谢情况,并对代谢产生的肽激活麸质蛋白特异性T细胞的能力进行了考察。结果发现,从CeD患者十二指肠分离的铜绿假单胞菌可通过其弹性蛋白酶活性将麸质蛋白降解为免疫原性肽,激活CeD患者的麸质蛋白特异性T细胞,而来自健康受试者的乳杆菌可通过产生蛋白酶将这些免疫原性肽继续降解为非免疫原性肽,从而降低CeD的发病率。
(2)降低肠屏障的通透性,防止免疫原性多肽进入固有层引发CeD。醇溶蛋白可引起zonulin的释放,导致肠上皮细胞骨架重排,降低occludin和ZO-1蛋白之间的相互作用,增加肠道通透性,使肠腔中的抗原转移到黏膜下层,从而引发CeD[46]。因益生菌具有增强肠道屏障,降低肠道屏障通透性的作用,这种情况可通过益生菌给药来预防。
(3)通过调节肠道菌群平衡以及宿主免疫反应减少CeD的发生。研究发现,CeD患者的粪便样品(代表失衡的肠道菌群)能显著增加外周血单核细胞中TNF-α的产生和CD86的表达,同时降低IL-10的产生和CD4的表达,而长双歧杆菌ES1和双歧杆菌ES2处理的外周血单核细胞与CeD患者的粪便样品共孵育可使促炎细胞因子的产生减少,IL-10的产生增加[47]。
4 小结与展望
肠道屏障功能对肠道健康至关重要,屏障功能受损会引发多种肠道疾病。益生菌可通过促进上皮细胞增殖分化、降低肠道上皮细胞通透性、促进黏蛋白的表达和分泌、竞争黏膜黏附位点、调节肠道菌群组成及调节免疫反应等途径改善肠道屏障功能,维持机体健康。益生菌作为生物治疗剂具有安全、高效的特点,不仅可避免由抗生素的大量使用产生的细菌耐药性,还可调节肠道菌群和增强机体免疫力。
值得注意的是,益生菌对肠道屏障的调节作用具有菌株特异性和宿主特异性。不同菌株对同一宿主可能产生不同的影响,同一菌株对不同宿主产生的效果也会受到宿主的生理、免疫、饮食、药物使用等因素的干扰。研究发现,乳酸菌在体外对HT-29细胞TJ蛋白的调节作用表现出显著的种间和种内差异,另外,这些菌株在缓解葡聚糖硫酸钠诱导的小鼠结肠炎症状(包括疾病活动指数、组织损伤和炎症)方面也表现出不同的效果,并且这种缓解效果与其修复TJ蛋白的能力高度相关[48-49]。因此,需要对特定菌株进行更全面的研究,进一步明确相关菌株改善肠屏障功能的具体作用机制及主要的生物效应物质,为菌株的应用提供强有力的科学依据。
然而,活益生菌在给宿主肠道健康带来有益影响的同时,也会给免疫功能低下的人群带来一些潜在的副作用,如腹胀、腹泻、过度免疫刺激、全身感染、败血症和抗生素耐药基因的转移等。为了避免这些风险,人们越来越关注灭活益生菌或益生菌发酵代谢产物。经热处理的益生菌细胞、无细胞上清液和纯化的生物活性成分具有与活益生菌相似的效应,并且在安全性、储存、运输方面更具有优势。因此,将其作为活益生菌的替代制剂,通过改善肠道屏障完整性来预防和治疗各种胃肠道疾病可能是一种更有效的策略。
猜你喜欢
江苏安全生产(2022年8期)2022-11-01
小资CHIC!ELEGANCE(2021年36期)2021-10-15
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
艺术评鉴(2020年5期)2020-04-30
人大建设(2018年10期)2018-12-07
中外食品工业(2018年1期)2018-03-13
家庭百事通·健康一点通(2017年11期)2017-11-29
中外食品工业(2017年7期)2017-07-07
小星星·阅读100分(高年级)(2015年11期)2015-11-28
