
自噬在光气诱导急性肺损伤中的作用
2022-08-17 02:33:46何改花李宏伟于卫华刘江正涂永梅孔德钦李文丽
空军军医大学学报 2022年3期
何改花 ,李宏伟,于卫华,刘江正,涂永梅,龙 子,孔德钦,刘 瑞,李文丽
(空军军医大学军事预防医学系军事毒理学与防化医学教研室,陕西省自由基生物学与医学重点实验室,教育部特殊作业环境危害评估与防治重点实验室,陕西 西安710032)
光气,学名碳酰氯,是生产中必不可少的高产量中间体,主要用于生产重要化学品和各种类型的塑料、药品、农药、染料和日用品[1]。光气还是一种剧毒的窒息性气体,在空气中分散度强,能较快形成杀伤浓度,被列为三类 A 级有机剧毒品。光气全球年度总产量超过850万吨[2],而我国年生产量超过300万吨,是世界上光气产量和使用量最大的国家。由于光气易生产、来源广,其泄露导致众多人员伤亡和严重社会危害的威胁始终存在。
光气主要通过吸入途径暴露,高浓度光气可导致人出现急性肺损伤(acute lung injury,ALI),严重者可发生急性呼吸窘迫综合征,甚至在短时间内窒息死亡。我们团队从上世纪50年代开始对光气进行研究,是国内最早研究光气肺损伤的课题组。近20年本教研室对光气的中毒机制、防治研究虽然取得了很大的进展[3],但光气中毒机制仍然不清,目前仍无有效救治办法[4]。光气的防治是世界范围内的难题之一,也是我国亟待解决的公共卫生问题。国内外报道,光气致损机制主要有炎症作用、酸烧伤作用、氧化损伤作用、心肺神经失调和Ⅱ型肺泡损伤学说,而炎症作用是导致光气诱导ALI的重要原因之一。大量文献表明,光气中毒后,炎症细胞因子大量释放,通过扳机样作用,触发中性粒细胞释放大量的髓过氧化物酶,导致炎症介质大量释放,产生肺损伤[5]。然而,光气导致炎症损伤的调控机制仍然不清。自噬是通过溶酶体途径对细胞内物质和细胞器降解再利用的过程,是维持细胞稳态和细胞更新的重要方式,在免疫、炎症的调控中非常重要[6]。近期的研究表明,自噬在ALI中发挥了重要的调控作用,如增强自噬减轻了结晶二氧化硅诱导的炎症细胞浸润,并减轻肺损伤[7]。自噬是否在光气诱导的ALI中发挥作用未见报道。因此,深入探讨自噬在光气中毒肺损伤中的作用及机制具有重要的临床意义。本文研究了自噬在光气染毒动物和细胞模型中的变化及机制,旨在从新的角度探讨光气中毒诱导肺损伤的新机制,并为寻找光气中毒救治的新靶点提供实验基础。
1 材料与方法
1.1 材料
光气(10 L/150 bar气瓶)购自德国林德公司;光气鼻吸入染毒设备由德国拜耳公司吸入毒理实验室赠送;光气细胞染毒设备购自德国Cultex公司;雄性Sprague-Dawley(SD)大鼠购自空军军医大学实验动物中心;人支气管上皮细胞(BEAS-2B)购自iCell公司;BCA 蛋白定量试剂盒购自Thermo 公司;自噬双标腺病毒(mRFP-GFP-LC3)购自汉恒生物公司;LAMP2a抗体购自Abcam公司(ab12506);P62抗体购自Abcam公司(ab109012);LC3-Ⅱ/Ⅰ抗体购自 Abcam公司(ab109012);总RNA提取试剂盒、反转录试剂盒和扩增试剂购自TIANGEN公司。
1.2 方法
1.2.1 实验分组与动物模型 所有实验均按照空军军医大学动物福利和使用委员会批准的程序进行[SCXK(陕)2019-001]。大鼠在恒温[(23±1)℃]和恒湿 [(55±5)%]条件下饲养,并遵循严格的12 h/12 h明暗周期。将40只SD雄性大鼠(SPF级,体质量180~220 g)随机分为4组:对照组、染毒后3 h组、染毒后6 h组、染毒后12 h组,每组各10只。对照组大鼠鼻吸入空气,染毒组大鼠鼻吸入41 mg/m3光气30 min,构建光气诱导的ALI动物模型。鼻吸入染毒参照文献[8]的方法。大鼠经鼻吸入光气染毒后,置于正常环境饲养,分别在3 、6、12 h使用20 g/L戊巴比妥钠腹腔麻醉后,解剖大鼠,收集肺组织以及肺泡灌洗液(bronchoalveolar lavage fluid,BALF)。
1.2.2 BALF总蛋白测定 肺组织称质量后,在左主支气管结扎左肺,将灌洗针缓慢插入气管,随后用棉绳在气管中间1/2处打结。使用注射器吸取3 mL 37 ℃生理盐水,注入右肺内,停留30 s后吸出,继续重复灌洗一次后将灌洗液吸出,注入EP管中。另取3 mL生理盐水重复以上操作,将2次收集到的灌洗液混合在同一EP管中备测。将收集到的BALF以 3 000 r/min离心5 min,收集上清液并分装。使用BCA试剂盒测定BALF中的总蛋白浓度,具体步骤严格根据试剂盒说明书操作,最后使用全波段酶标仪(Infinite M200PRO,奥地利)检测BALF的总浓度A562 nm值并计算蛋白浓度。
1.2.3 肺组织HE染色切片制作及观察 取大鼠左肺上叶, 经过40 mL/L多聚甲醛固定—蒸馏水清洗—乙醇梯度脱水—二甲苯透明—石蜡包埋—切片(LeicaRM2135型轮转式切片机,4 μm)—苏木素-HE染色等步骤后,使用正置光学显微镜(Nikon,日本)观察HE染色切片并拍照。
1.2.4 Western blotting测定蛋白表达 取肺组织50 mg(冰上操作),加入RIPA裂解液,经组织研磨器(新芝生物,宁波)充分研磨后,在室温放置5 min,12 000 r/min离心10 min,提取上清液。配制不同浓度的SDS-PAGE分离胶,以浓缩胶90 V,分离胶120 V的电压使用SDS-PAGE垂直电泳仪(Bio-Rad,美国)进行电泳,电泳结束后使用Trans-blot蛋白转膜仪(Bio-Rad,美国)将蛋白转移至PVDF膜上,随后将膜浸泡在50 g/L脱脂牛奶中,室温封闭2 h,封闭完成后,使用TBST洗净残余封闭液体。根据不同的目的蛋白敷相应的一抗, 4 ℃摇床过夜,次日使用TBST洗涤3次,每次10 min,加入二抗(1∶5 000),室温孵育2 h后,使用TBST洗涤3次,每次10 min,最后用全自动化学发光仪(Bio-Rad, 美国)显影。使用Image J软件分析条带的灰度值,计算目的蛋白的相对表达量。
1.2.5 细胞培养 使用含100 mL/L胎牛血清的DMEM高糖培养基在37 ℃、50 mL/L CO2的条件下培养BEAS-2B细胞(人源支气管上皮细胞)。当处于对数生长期的细胞生长至密度为80%~ 90%时,使用2.5 g/L胰蛋白酶消化,1 000 r/min离心5 min收集细胞,计数后按密度接种于不同类型培养板,分组处理后进行检测。
1.2.6 细胞染毒与分组 将BEAS-2B细胞以1×107个/L的密度接种于Transwell六孔板细胞培养小室中,对照组3个小室,染毒组3个小室。待细胞贴壁后,吸去小室中的液体,将小室放入气液界面细胞暴露系统(Cultex Technology, 德国)中。对照组给予空气,染毒组以3.2 mg/m3光气动态染毒60 min,构建光气诱导的细胞损伤模型。
1.2.7 细胞自噬流的测定 将BEAS-2B细胞接种至共聚焦皿中,接种密度为1×108个/L,待细胞贴壁后,转染自噬双标腺病毒(mRFP-GFP-LC3)4 h,构建mRFP-GFP-LC3转染BEAS-2B细胞,然后光气暴露1 h,继续培养12 h,完毕后使用40 mL/L多聚甲醛固定、封片,激光共聚焦显微镜(Olympus,日本)拍照分析。
1.2.8 实时荧光定量PCR测定炎症因子mRNA的表达 使用总RNA提取试剂盒提取细胞RNA。使用逆转录试剂盒将RNA逆转录为cDNA,采用SYBR Green PCR Master Mix进行实时定量PCR。实验中所使用引物的详细信息见表1。

表1 基因引物序列
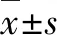
2 结果
2.1 光气染毒大鼠ALI模型的建立
2.1.1 光气染毒对大鼠肺湿质量和肺体比的影响 与对照组相比,光气染毒后大鼠肺湿质量和肺体比值在染毒后3 h开始增加,到染毒后6 h明显增加(P<0.05),12 h显著增加(P<0.01,图1)。结果表明,光气染毒后大鼠肺湿质量、肺体比与染毒后时间存在时间效应关系。
aP<0.05, bP<0.01 vs对照组(n=10)。图1 光气染毒后不同时间点大鼠肺湿质量和肺体比的变化
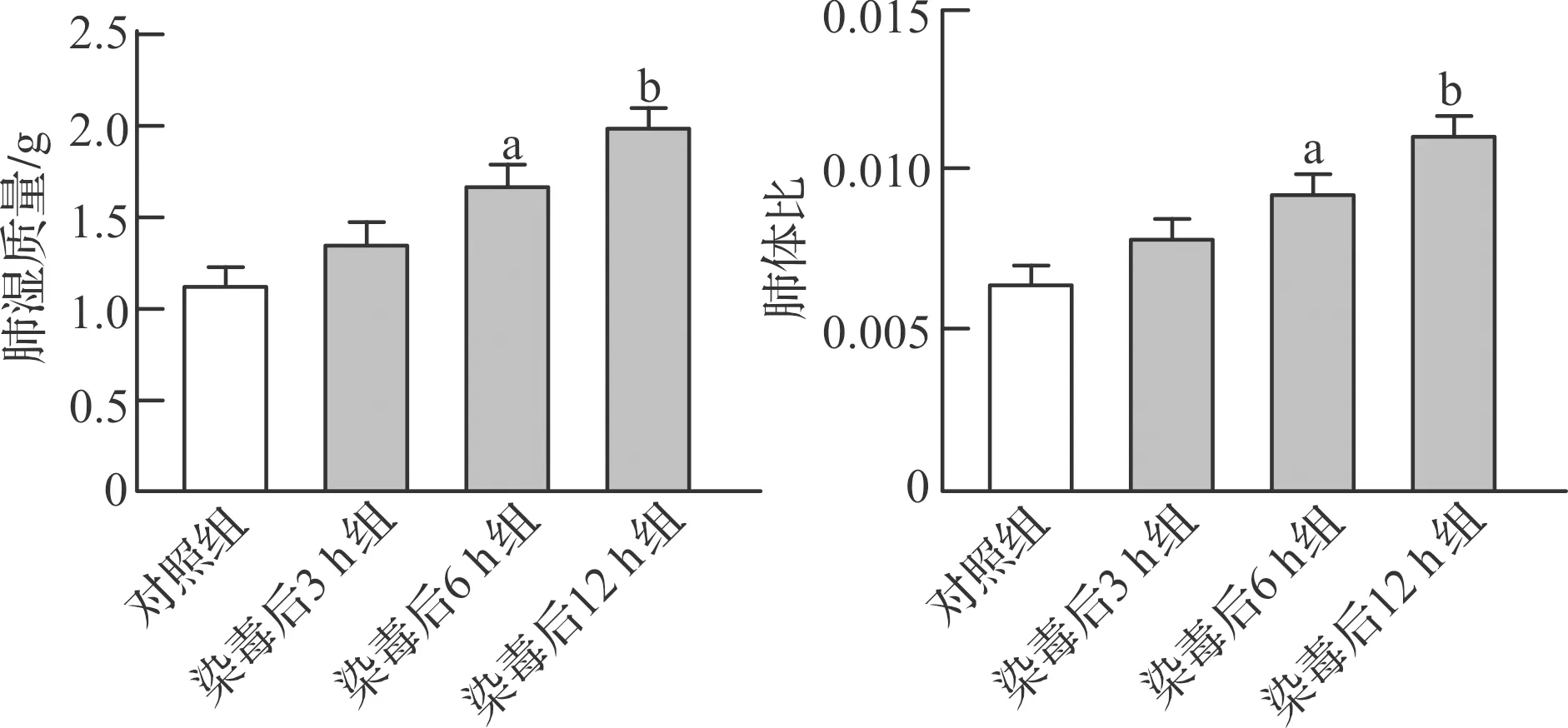
2.1.2 光气染毒对大鼠肺组织病理学的变化 HE染色观察肺组织病理结构变化,结果表明对照组大鼠肺组织结构正常,表现为肺泡壁薄,肺泡结构完好 (图2A);光气染毒后3 h组大鼠肺间质增厚,炎症细胞浸润增多 (图2B);染毒后6 h组大鼠肺泡腔有少量渗出液,炎症细胞显著增多 (图2C);染毒后12 h组大鼠肺组织炎症细胞大量增多,肺泡结构破坏,肺泡腔融合并可见大量渗出液 (图2D)。
A:对照组;B:染毒后3 h组;C:染毒后6 h组;D:染毒后12 h组。HE ×200。图2 光气染毒后不同时间点大鼠肺组织病理改变
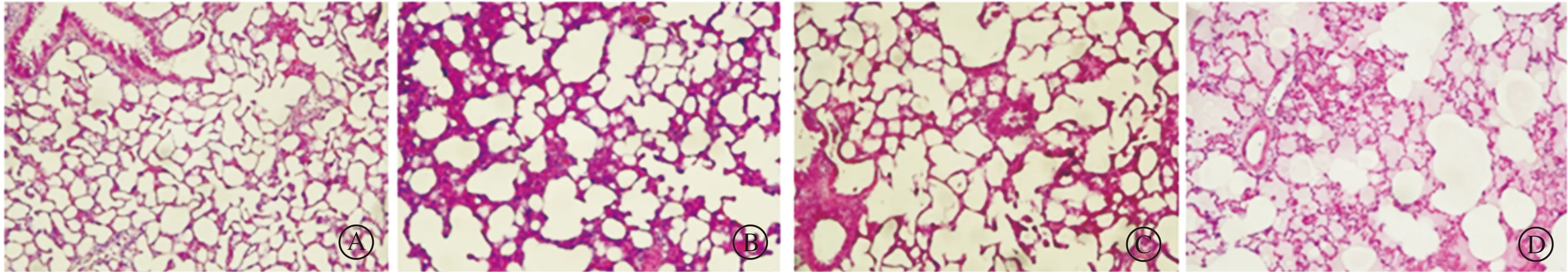
2.1.3 光气染毒对大鼠BALF中总蛋白含量的影响 与对照组相比,光气染毒后3 h大鼠BALF中总蛋白浓度开始增加,到染毒后6 h有明显增加 (P<0.05),12 h增加最显著(P<0. 01,图3)。
aP<0.05, bP<0.01 vs对照组(n=10)。图3 光气染毒后不同时间点大鼠BALF中总蛋白浓度的变化
2.2 光气染毒大鼠ALI中自噬相关分子的变化
为了观察自噬在光气诱导大鼠ALI中的作用,用Western blotting检测自噬相关蛋白的变化(图4A)。结果显示,与对照组相比,溶酶体相关膜蛋白 2a型(LAMP2a)和微管相关蛋白 1 轻链 3 (LC3-Ⅱ/Ⅰ) 蛋白在染毒后 6、12 h显著降低(P<0.01, 图4B~C), P62蛋白染毒后6、12 h显著增加(P<0.01, 图4D)。以上结果提示光气染毒后6、12 h显著抑制了肺组织自噬的发生。
2.3 光气致人支气管上皮细胞损伤模型的建立
光气染毒BEAS-2B细胞12 h后,经PCR测定,与对照组相比,IL-1β、IL-6和IL-8 mRNA水平显著升高(P<0.01,图5)。

A:肺组织LAMP2a、LC3-Ⅱ/Ⅰ及P62蛋白水平的表达;B:LAMP2a蛋白相对灰度值变化;C:P62蛋白相对灰度值变化;D: LC3-Ⅱ/Ⅰ蛋白相对灰度值变化。LAMP2a:溶酶体相关膜蛋白2a型,LC3:微管相关蛋白1轻链3。 aP<0.05, bP<0.01 vs对照组。图4 光气染毒后不同时间点大鼠肺组织自噬相关蛋白表达的变化

A: IL-1β mRNA相对表达;B:IL-6 mRNA相对表达;C:IL-8 mRNA相对表达。 bP<0.01 vs对照组(n=3)。图5 光气染毒细胞后对炎症相关mRNA表达的影响
2.4 自噬在光气染毒BEAS-2B细胞损伤中的作用
2.4.1 光气染毒对BEAS-2B自噬流的影响 为了观察自噬在光气诱导的BEAS-2B细胞中的作用,使用自噬双标腺病毒(mRFP-GFP-LC3)感染BEAS-2B细胞,结果见图6。经自噬双标腺病毒(mRFP-GFP-LC3)感染BEAS-2B细胞4 h,光气染毒1 h后,换完全培养基培养12 h,发现染毒组与对照组相比,绿色荧光蛋白 (green fluorescent protein, GFP)和红色荧光蛋白 (red fluorescent protein, mRFP)斑点显著减少,表明溶酶体和LC3减少(图6A~B),GFP图片和mRFP图片重合后出现的黄色斑点和红色斑点均减少,表明自噬体和自噬溶酶体均减少(图6C)。

A:LC3荧光斑点代表图;B:GFP 和 mRFP 点数/细胞的平均数;C:GFP图像和mRFP图像 merge 后每个细胞自噬体和自噬溶酶体的平均数。 aP<0.05, bP<0.01 vs对照组GFP 和mRFP。GFP:绿色荧光蛋白;mRFP:红色荧光蛋白。 标尺为20 μm。图6 光气染毒BEAS-2B细胞后自噬流的变化
2.4.2 自噬激动剂雷帕霉素干预对光气染毒诱导BEAS-2B细胞损伤的影响 光气染毒BEAS-2B细胞后,给予0.5 μmol/L雷帕霉素处理12 h。结果显示,给予雷帕霉素后,与染毒组相比,对照组及染毒+RaPa组IL-1β、IL-6、IL-8 mRNA水平均显著降低(P<0.05,图7)。
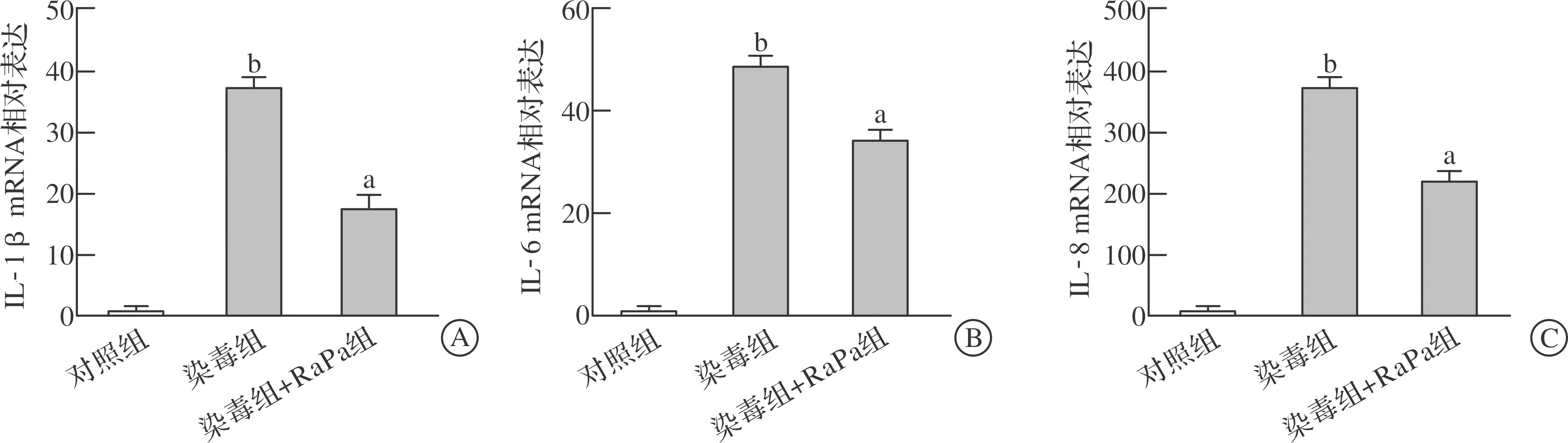
A: IL-1β mRNA相对表达;B:IL-6 mRNA相对表达;C:IL-8 mRNA相对表达。 RaPa:雷帕霉素。 aP<0.05 vs 染毒组, bP<0.01 vs对照组 (n=3)。图7 雷帕霉素干预对光气染毒诱导细胞炎症因子表达的影响
3 讨论
光气作为一种军用窒息性化学战剂,被广泛用于第一次和第二次世界大战中。侵华日军在我国曾生产和使用大量的化学武器,造成我国大量军人和平民伤亡。日本战败撤出我国时,在我国19个省市遗弃、埋藏了40多万枚化学弹药,即日本遗留的化学武器,简称“日遗化武”,其中大量为光气炸弹或炮弹。1986年在山西等地发现了数千发光气弹,天津、珠江口等地也挖出了大量光气炮弹,含光气的日遗化武对我国人民的生命安全造成了巨大的现实威胁。光气作为一种双用途化学战剂,也是平时化学生产中必不可少的高产量中间体,主要用于生产重要化学品的原料,例如亚甲基二苯基二异氰酸酯和甲苯二异氰酸酯。光气也用于生产各种类型的塑料、药品、农药、染料和日用品。许多化工产品如氯仿、三氯乙烯、塑料等燃烧也会产生光气。我国是世界上光气产量和使用量最大的国家,年产量超过300万吨,职业性接触人员众多。目前,我国光气安全生产态势严峻,许多中小型工厂光气生产设备陈旧,很容易造成光气泄露,导致众多人员伤亡和严重社会危害。如2004年6月15日,福州市发生光气泄漏事故,导致260人光气中毒。天津、大连、宿迁等地也都曾发生过光气泄露致人伤亡事故,损失惨重,教训深刻。目前,光气中毒的治疗是世界范围内的难题之一,也是我国亟待解决的公共卫生问题。
光气中毒主要靶器官是肺组织,引起中毒者发生ALI,严重者可窒息死亡,但近百年来,国际上始终未阐明光气肺水肿的发生机制。本课题组开展了大量研究,提出了酸烧伤、氧化应激、炎症作用、细胞凋亡等学说,并针对这些中毒机制,开展了大量光气中毒药物的筛选,虽然取得了一定进展,但仍然没有找到一种治疗光气致ALI的特效药物[9-10]。因此,深入解析光气导致ALI的分子机制,对于平战时光气中毒的救治具有重要意义。我们团队从20世纪50年代开始对光气进行研究,是国内最早研究光气肺损伤的课题组。近10年,我们与德国拜尔公司毒理室国际知名的吸入毒理学家JUERGEN PAULUHN教授进行了合作研究,建立了与国际接轨的吸入毒理学研究平台,并发表了大量相关研究文章和论著[11-13]。本文利用该平台,给予大鼠动态鼻吸入染毒1 230 mg/m3的光气30 min,大鼠肺湿质量、肺体比和BALF总蛋白在染毒后6 h明显增加,12 h增加到最高[14]。光镜下观察肺组织HE切片显示,光气染毒组大鼠肺间质增厚、炎性细胞增多、肺泡腔有渗出液。以上结果表明我们成功建立了鼻吸入光气染毒大鼠ALI模型。
为了更好地在细胞水平研究光气中毒机制及分子调控,必须建立光气染毒细胞损伤模型。目前,国际上未见此模型的相关报道。我们科室利用德国Cultex公司生产的国际先进的动态细胞染毒设备,真实地模拟了符合人体气血屏障特点的气液界面环境,暴露效率高,用3.2 mg/m3的光气动态染毒60 min构建体外光气染毒人支气管上皮细胞模型。我们的研究发现,光气暴露后导致BEAS-2B细胞发生显著的炎症反应,提示炎症反应在光气诱导的ALI中可能发挥了重要作用[15]。研究表明,光气暴露可引起大量中性粒细胞和单核巨噬细胞聚集在肺泡表面,释放大量的髓过氧化物酶,导致炎症介质大量释放,促进肺泡上皮细胞和肺毛细血管内皮细胞损伤,组织液大量渗出或漏出,引起ALI[15]。因此,我们将炎症指标作为光气染毒细胞损伤模型建立的评价指标。本实验发现光气染毒后,人支气管上皮细胞IL-1β、IL-6和IL-8 mRNA水平均显著升高,证明我们成功建立光气染毒细胞损伤模型,可用于下一步实验。
自噬是一种自我降解的过程,在去除错误折叠和聚集的蛋白质,清除受损细胞器(如线粒体、内质网),以及消除细胞内病原体方面发挥“管家”的作用[16]。在正常情况下,自噬不影响细胞的生存和正常功能,只有当机体自噬平衡被打破时(自噬增强或者不足),才会引起细胞功能异常,导致相关组织病理损害加重或者坏死。自噬对于细胞的生存和死亡具有双重作用,适度自噬有利于细胞生存,自噬不足和过度自噬均可导致细胞凋亡。自噬可通过抑制细胞炎症因子而恢复细胞的正常状态,从而减轻组织炎症和相关损伤[17],已经发现自噬可能通过直接抑制炎症复合物和间接清除炎症刺激物如受损的细胞器来保护细胞免受持久炎症的侵害。研究报道[18],在脂多糖诱导的ALI小鼠模型中,肺组织中哺乳动物雷帕霉素靶蛋白(mTOR)磷酸化显著增高,自噬显著降低。在人支气管上皮细胞中,脂多糖增加mTOR磷酸化,抑制自噬,而敲减mTOR可通过增强自噬,减轻脂多糖诱导的支气管上皮细胞炎症反应。也有研究报道,自噬抑制氧化铜纳米粒子诱导的ALI[19]。上述研究提示,自噬可能在肺损伤的发生和发展过程中扮演了重要角色。
我们前期观察到光气所致ALI肺组织多种细胞发生凋亡[20-21],而在某些情况下,自噬抑制凋亡是细胞的存活途经,二者通路相互关联、互为调控,但自噬是否也参与光气所致ALI目前未见研究报道。自噬相关蛋白LC3-Ⅱ/Ⅰ比值、P62蛋白和LAMP2a蛋白表达改变被广泛作为自噬流活化或抑制的标志物,通常认为LC3-Ⅱ/Ⅰ升高、LAMP2a蛋白增多、P62蛋白水平下降代表了自噬活化[22],反之表示自噬被抑制。本实验在动物模型中观察到,大鼠肺组织LAMP2a和LC3-Ⅱ蛋白表达在光气染毒后6 h开始下降,而P62蛋白表达在6 h开始增加,表明光气诱导大ALI过程中肺组织自噬降低。在细胞模型中我们观察到同样的现象,光气染毒感染自噬双标腺病毒(mRFP-GFP-LC3)BEAS-2B细胞,重合后的黄色斑点和红色斑点均减少,表明自噬体和自噬溶酶体都减少,提示光气所致支气管上皮细胞损伤中自噬流降低。通过光气染毒体内和体外模型,我们证明了光气染毒后6~12 h显著抑制了肺组织自噬的发生,自噬抑制可能是光气中毒的重要发病机制之一。
为进一步验证自噬在光气所致ALI中的作用,我们给予光气染毒的BEAS-2B细胞0.5 μmol/L自噬激动剂雷帕霉素处理12 h,结果表明,雷帕霉素干预组与光气染毒组相比,细胞内IL-1β、IL-6、IL-8 mRNA表达水平均显著下调,提示雷帕霉素激活自噬能够有效减轻光气所致细胞的验证反应。因此,我们进一步确证自噬抑制可能是光气诱导ALI的重要机制之一,尤其是参与了光气诱导肺组织炎症损伤过程。
综上所述,我们证实光气染毒能造成大鼠ALI和支气管上皮细胞发生炎症反应,同时抑制肺组织自噬。自噬激动剂雷帕霉素干预可有效减轻光气对细胞的炎症反应,提示光气染毒后自噬降低可能是光气所致ALI的重要机制 。下一步我们将在动物模型上使用自噬激动剂或抑制剂验证该结论。同时,光气诱导自噬降低的具体机制也不明确,可能是自噬流的阻断或者是自噬流过度活化所致,具体机制还需要我们进一步研究。
猜你喜欢
数学物理学报(2021年4期)2021-08-30 08:27:48
新世纪智能(数学备考)(2020年10期)2021-01-04 00:37:50
福建轻纺(2020年12期)2020-12-16 08:23:08
科学导报·学术(2020年19期)2020-07-09 11:21:37
氯碱工业(2020年10期)2020-03-01 17:44:02
实验动物科学(2019年6期)2019-12-02 12:48:26
环境与生活(2018年7期)2018-09-10 13:55:56
中成药(2017年12期)2018-01-19 02:06:52
首都公共卫生(2017年3期)2017-11-29 01:23:09
中国交通信息化(2017年8期)2017-06-06 07:16:47
