
川芎α-淀粉酶抑制剂基因的原核表达与条件优化
2022-08-16 00:45邓楷煜安秋菊陈安琪俞继华张富丽
生物学杂志 2022年4期
邓楷煜,安秋菊,陈安琪,俞继华,张富丽,廖 海
(1.西南交通大学 生命科学与工程学院,成都 610031;2.四川省农业科学院分析测试中心,成都 610066)
α-淀粉酶抑制剂(α-amylase inhibitor,ASI)是具有Kunitz结构域的活性肽,其由180~200个氨基酸残基组成,含有2个二硫键。ASI能够与α-淀粉酶形成靶酶-抑制剂复合物,从而抑制α-淀粉酶的活性。ASI的分子结构中抑制淀粉酶的活性区域分布于C末端,主要的活性氨基酸残基为谷氨酸[1]。目前已从大麦、小麦与大米中分别发现该类型抑制剂的蛋白及其基因序列[1-4]。分析大麦来源的BASI与大麦淀粉酶-2(AMY2)复合物的晶体结构,发现Ca2+、水分子与BASI形成氢键网络,加强了复合物表面的静电作用,提高了复合物的稳定性[2]。ASI在植物的生长发育与抗逆防御等重要生理过程中起关键的调节作用[5],不仅对多种昆虫体内的α-淀粉酶发挥抑制作用,影响昆虫吸收营养,起抗虫的作用[6-8],还能抑制人体肠道内α-淀粉酶的活性,减少糖在人体内的消化吸收,具有降血糖、降血脂的功效[9-10]。目前,ASI已广泛应用在减肥、糖尿病治疗和抗虫害等领域。因此,为更好地开展ASI基因的功能鉴定、应用、结构生物学及转基因植物培育等研究,需要提高LASI在宿主中的表达效率。
中药川芎(LigusticumchuanxiongHort)是四川省的道地药材,在云南、甘肃等地均广泛分布,具有降糖、祛风止痛、行气等多种功效[11],但是川芎在生长过程中容易受害虫侵害[6]。课题组在前期已成功扩增出川芎α-淀粉酶抑制剂基因的全长cDNA序列(Ligusticumchuanxiongα-amylase inhibitor,LASI,GenBank登录号 KX580040),结合已有文献资料,推测川芎α-淀粉酶抑制剂(LASI)具有降糖作用与抗虫活性[5-6],但仍需进一步验证。文章以大肠杆菌为宿主细胞,确定LASI的最适表达载体与最优诱导表达条件。为达此目标,构建pET28a-LASI等4种不同的表达质粒,初步考察4种不同表达质粒在大肠杆菌中的表达效率,发现pET28a是LASI基因的较适合表达载体,随后以pET28a-LASI作为表达质粒,探究诱导时间、IPTG浓度与诱导温度对LASI重组蛋白表达的影响,并检测纯化后的LASI重组蛋白对猪源α-淀粉酶[12]的抑制活性。研究结果为LASI的大规模发酵生产、糖尿病治疗、抗虫及相关转基因植物培育[13-14]奠定理论基础。
1 材料与方法
1.1 材料
重组质粒与转化菌:LASI-T重组质粒由课题组构建完成并保存于TOP10菌株中。试剂与培养基:NdeI、XbaI、BamH I、NotI、NcoI、XhoI、EcoR I、T4 DNA连接酶、异丙基硫代半乳糖苷(IPTG)、卡那霉素、氯霉素均购自上海生工生物工程有限公司;蛋白Marker购自Thermo公司;猪源淀粉酶购自Sigma公司。抗体:Anti-His(Rabbit)、Anti-Rabbit IgG,HRP-labeled(Goat)购自Thermo公司。
1.2 表达载体的筛选
以LASI-T重组质粒为目标,依次设计引物(表1),经PCR扩增出的LASI基因片段分别连入载体pCznI的NdeI(CATATG)-XbaI(TCTAGA)位点之间、载体pGEX-4T-1的BamH I (GGATCC)-NotI(GCGGCCGC)位点之间、载体pET22b的NcoI(CCATGG)-XhoI(CTCGAG)位点之间、载体 pET28a的BamH I (GGATCC)-EcoRI(GAATTC)位点之间,获得重组质粒pCznI-LASI、pGEX-4T-1-LASI、pET22b-LASI及pET28a-LASI。将获得的重组质粒pCznI-LASI、pGEX-4T-1-LASI、pET22b-LASI及pET28a-LASI分别转入Rosetta感受态细胞中。
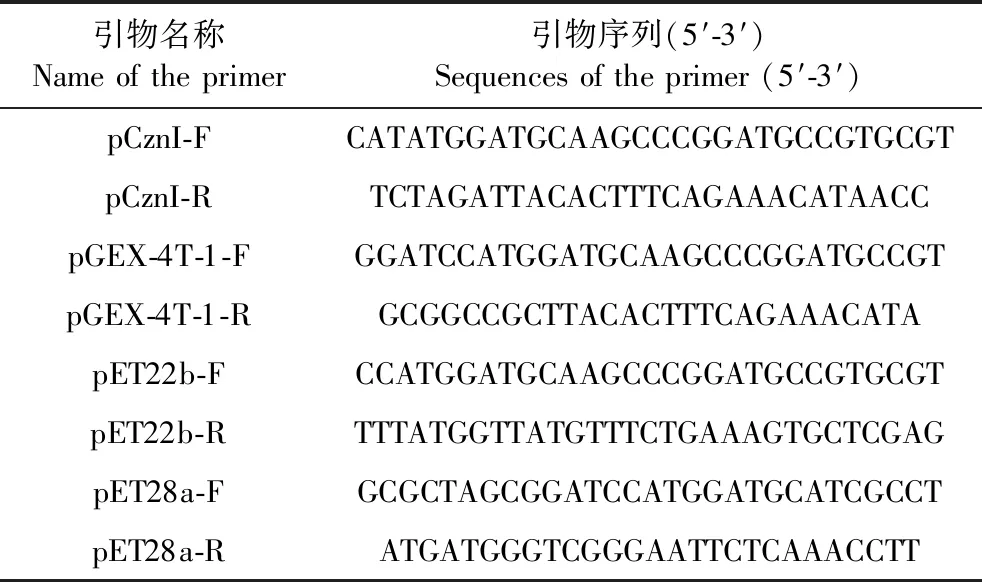
表1 引物设计Table 1 Primer design
1.3 转化菌的培养和诱导表达
在含50 μg/mL卡那霉素(Kan) 和25 μg/mL氯霉素(Chl)的LB液体培养基中活化含有pET28a-LASI重组质粒的Rosetta菌株。在37 ℃下摇床培养至OD600值大于0.6后,加入诱导剂(IPTG)使终浓度为1.0 mmol/L,在诱导温度为27 ℃、摇床转速为200 r/min和诱导时间为8 h的条件下诱导LASI蛋白高效表达。于超声细胞破碎机破碎菌体,随后取样进行蛋白电泳检测。以未加IPTG诱导的重组菌株为对照组。
在含50 μg/mL氨苄西林(Amp)的LB液体培养基中活化含有pCznI-LASI、pGEX-4T-1-LASI的Rosetta菌株。分别在37 ℃下摇床培养至OD600值大于0.6后,加入诱导剂(IPTG)使终浓度为0.5 mmol/L,在诱导温度为20 ℃,转速为220 r/min的条件下诱导蛋白表达。
在含50 μg/mL氨苄西林(Amp)的自诱导培养液中活化含有pET22b-LASI的Rosetta菌株。在37 ℃下摇床培养至OD600值大于0.6后,在诱导温度20 ℃、转速200 r/min的条件下诱导蛋白表达。
1.4 诱导适宜时间的筛选
经过初步筛选,在Rosetta菌株中含有pET28a-LASI重组质粒的蛋白表达量最高,因此选择该基因工程菌为目标菌株,在此基础上,分别探究诱导时间、IPTG浓度和诱导温度对pET28a-LASI重组蛋白表达的影响。
在37 ℃下将重组菌株培养至OD600值为0.6后,加入IPTG使终浓度为1.0 mmol/L。在27 ℃下诱导表达,培养1、2、4、8和24 h 后分别取样进行蛋白电泳检测。将含有pET28a空载的Rosetta菌株作为空白组,未加IPTG诱导的重组菌株作为对照组。
1.5 IPTG浓度的筛选
在37 ℃下将含有pET28a-LASI重组质粒的Rosetta菌株培养至OD600值为0.6后,将IPTG分别加入5个相同的培养瓶中,使IPTG终浓度分别为0.1、0.2、0.5、1.0和2.0 mmol/L,在27 ℃下诱导表达,8 h后取样进行蛋白电泳检测。将含有pET28a空载的Rosetta菌株作为空白组,未加IPTG诱导的重组菌株作为对照组。
1.6 诱导培养温度筛选
在37 ℃下将含有pET28a-LASI重组质粒的Rosetta菌株培养至OD600值为0.6后,加入IPTG使终浓度达到1.0 mmol/L。在17 ℃、22 ℃、27 ℃、32 ℃和37 ℃ 等5个温度下分别诱导表达8 h后取样进行蛋白电泳检测。将含有pET28a空载的Rosetta菌株作为空白组,未加IPTG诱导的重组菌株作为对照组。
1.7 表达产物纯化
在IPTG浓度为1.0 mmol/L,诱导时间8 h,诱导温度27 ℃的条件下,诱导表达LASI重组蛋白,于超声波细胞破碎机破碎菌体,并离心收集上清液,将上清液加入Ni2+亲和柱结合3 h,取经400 mmol/L咪唑缓冲液洗脱后的目的蛋白LASI,将样品富集后进行蛋白电泳检测。将含有pET28a空载的Rosetta菌株作为空白组,未加IPTG诱导的重组菌株作为对照组。LASI蛋白浓度利用改良型BCA蛋白浓度测定试剂盒和全自动酶标仪检测。具体相关实验操作步骤参见试剂盒说明书,酶标仪检测波长为562 nm。
1.8 Western Blot分析
以兔源His 标签抗体作为一抗(稀释比1∶1 000),羊抗兔IgG为二抗(稀释比1∶5 000)对纯化的蛋白进行Western Blot鉴定。操作步骤如下:将获得的LASI蛋白进行SDS-PAGE电泳,取5 μL样品上样。聚丙烯酰胺凝胶于90 V下跑完积层胶,后将电压增加到200 V至电泳结束。电泳结束后,在恒压100 V、恒流250 mA条件下转膜1.5 h。电转结束后,用PBST洗涤4次,每次5 min。随后将膜置于5%脱脂奶粉封闭液中37 ℃封闭1 h。用封闭液稀释一抗,并将膜置于一抗稀释液中于4 ℃过夜。次日将膜取出用PBST洗涤4次,每次5 min。用含5%牛奶的封闭液稀释二抗,将膜置于二抗稀释液中37 ℃反应1 h。反应完毕后,取出膜置于干净的盒子中洗涤4次,每次5 min。ECL显影,曝光。
1.9 LASI重组蛋白抑制活性检测
分别配制0.15%淀粉溶液、I2-KI溶液、1 mg/mL的淀粉酶溶液备用。检测LASI对淀粉酶的抑制活性参考Erlanger法[15]和碘-淀粉呈色法,淀粉遇碘变蓝[16-17],在波长为640 nm下采用可见光光度计进行检测(表2)。
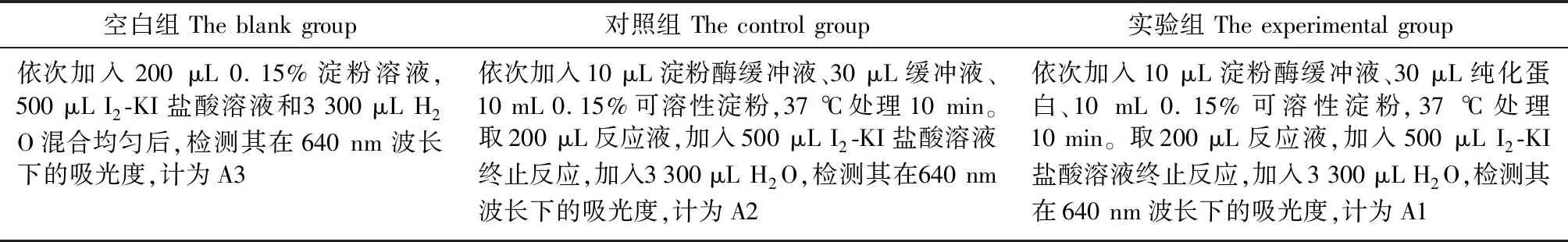
表2 抑制活性测定流程Table 2 Inhibitory activity assay process
淀粉酶抑制活性(IA)计算公式:IA=100%×{[(A3-A2) -(A3-A1)]/(A3-A2)}
2 结果与分析
2.1 不同表达载体LASI重组蛋白表达
分别采用pET28a、pCznI、pGEX-4T-1和pET22b构建重组表达质粒,于大肠杆菌Rosetta (DE3)中进行诱导表达,由实验结果(图1)可知,因诱导载体的不同,重组蛋白的表达产率差异较大。pCznI-LASI、pGEX-4T-1-LASI、pET28a-LASI均有表达,pET22b-LASI无明显表达,pET28a-LASI能成功诱导出LASI重组蛋白,并且有较多重组蛋白为可溶性形式。因此pET28a是LASI蛋白较好的表达载体。
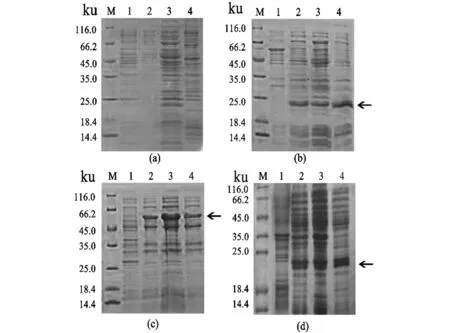
(a)pET22b-LASI原核表达(M:蛋白 Marker;1:pET22b-LASI未诱导;2:pET22b-LASI诱导后;3:pET22b-LASI诱导后上清液;4:pET22b-LASI诱导后沉淀)。(b)pCznI-LASI原核表达(M:蛋白 Marker;1:pCznI-LASI未诱导;2:pCznI-LASI诱导后;3:pCznI-LASI诱导后上清液;4:pCznI-LASI诱导后沉淀)。(c)pGEX-4T-1-LASI原核表达(M:蛋白 Marker;1:pGEX-4T-1-LASI未诱导;2:pGEX-4T-1-LASI诱导后;3:pGEX-4T-1-LASI诱导后上清液;4:pGEX-4T-1-LASI诱导后沉淀)。(d)pET28a-LASI原核表达(M:蛋白 Marker;1:pET28a-LASI未诱导;2:pET28a-LASI诱导后;3:pET28a-LASI诱导后上清液;4:pET28a-LASI诱导后沉淀)。图1 LASI基因的原核表达Figure 1 Prokaryotic expression of LASI gene
2.2 不同诱导时间LASI重组蛋白表达
由实验结果(图2)可知,在诱导温度和诱导剂(IPTG)浓度相同的情况下,1 h后pET28a-LASI重组蛋白开始表达,诱导时间在1~8 h,重组蛋白的表达量随着时间的延长有所增加,8 h后表达量逐渐趋于平稳,并没有明显增加。表明LASI重组蛋白的最佳诱导时间为8 h。
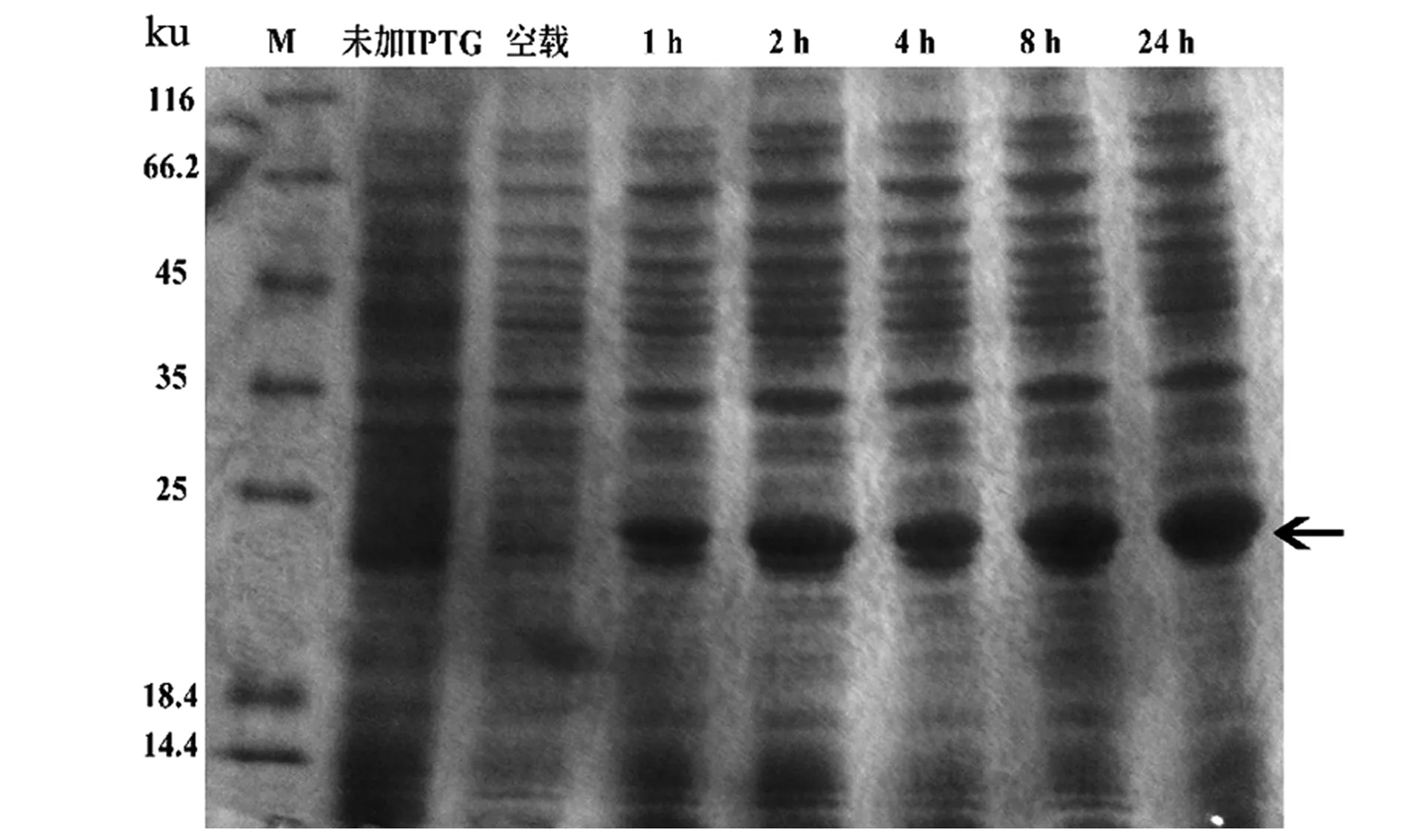
图2 诱导时间梯度对LASI蛋白表达的影响Figure 2 Effect of induction time gradient on LASI protein expression
2.3 不同IPTG浓度的LASI重组蛋白表达
由实验结果(图3)可知,在诱导温度和诱导时间相同的情况下,在IPTG终浓度为0.1 mmol/L时即开始表达出pET28a-LASI重组蛋白,当IPTG终浓度在0.1~1.0 mmol/L时,LASI重组蛋白的表达量随诱导剂浓度的增加并未大幅度增加,且当浓度达1.0 mmol/L时,表达量达到最大值。
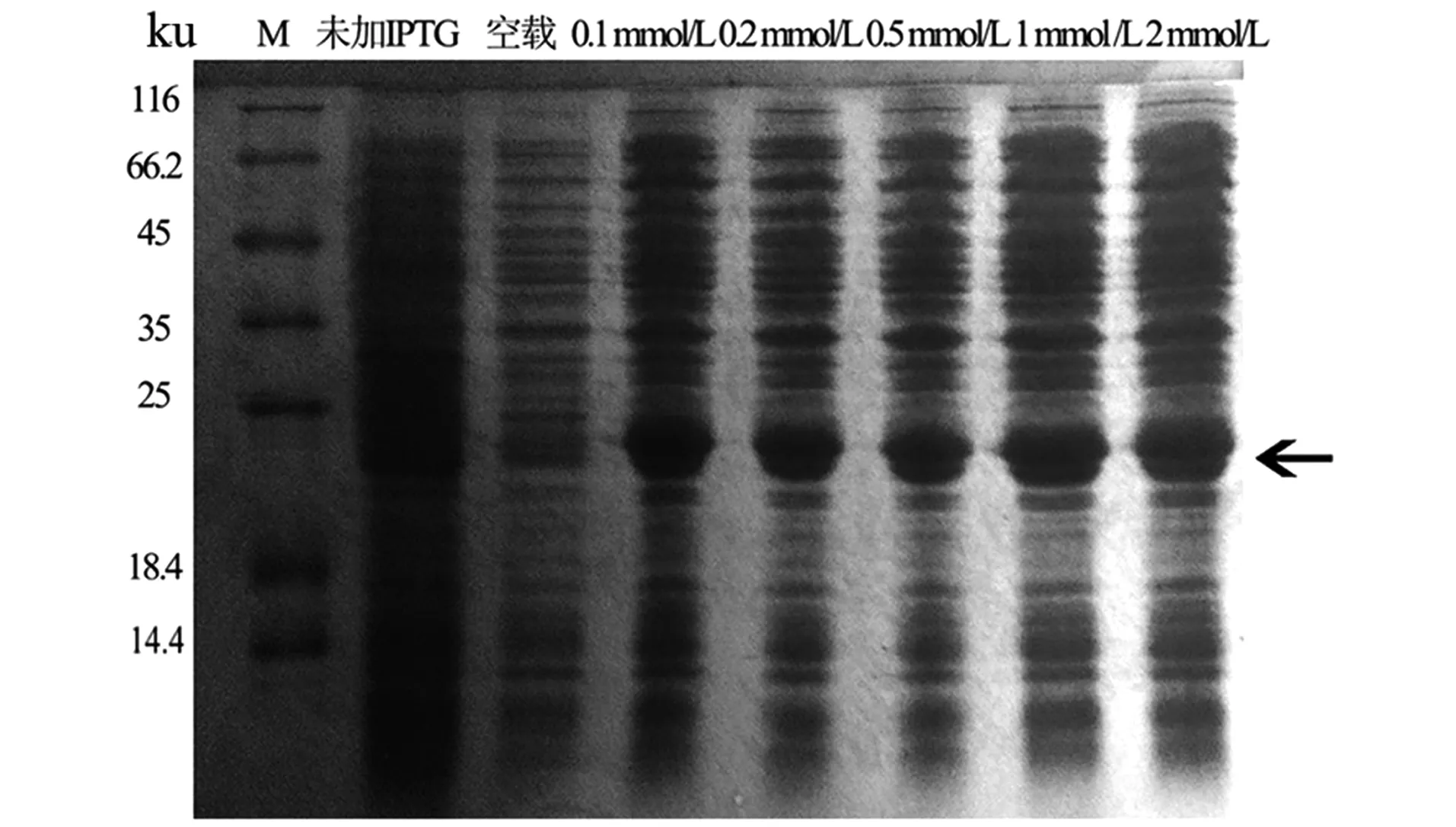
图3 不同IPTG诱导浓度对LASI蛋白表达的影响Figure 3 Effects of different IPTG induced concentrations on LASI protein expression
2.4 不同诱导温度LASI重组蛋白表达
由实验结果(图4)可知,在诱导时间和诱导剂浓度相同的情况下,诱导温度为17 ℃时,pET28a-LASI重组蛋白开始表达。蛋白表达量随着温度的增加而增加,但在27 ℃后表达量到达峰值且趋于稳定。表明27 ℃为诱导表达LASI重组蛋白的最佳温度。
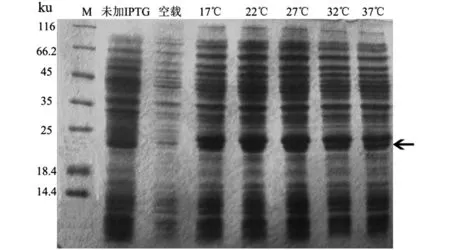
图4 不同诱导温度对诱导LASI蛋白表达的影响Figure 4 Effect of different induction temperature on the expression of LASI protein
2.5 LASI重组蛋白纯化结果
在最佳诱导条件(IPTG浓度为1.0 mmol/L、诱导温度27 ℃以及诱导时间8 h)下诱导表达LASI重组蛋白,将表达后的菌体经超声破碎后,破裂的菌体释放蛋白至重悬缓冲液中。经离心后取上层溶液,通过纯化柱纯化并进行SDS-PAGE电泳,实验结果如图5所示,纯化后的LASI重组蛋白显示出清晰的条带,检测LASI纯化蛋白的浓度为614 μg/mL,可用于后续活性检测。
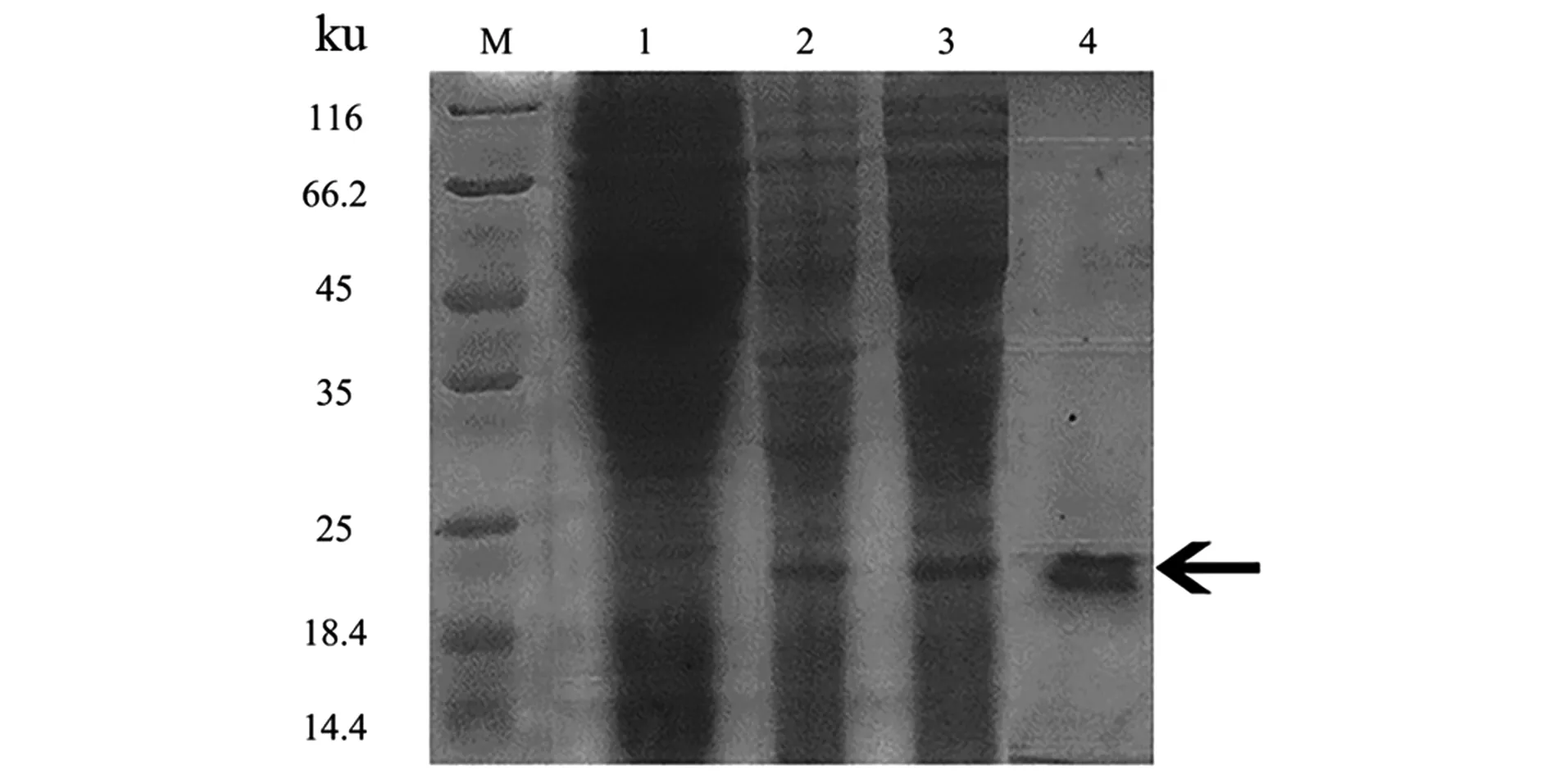
M:蛋白 Marker;1:pET28a-LASI未诱导;2:pET28a-LASI诱导后;3:pET28a-LASI诱导后上清液;4:纯化后的pET28a-LASI。图5 LASI蛋白纯化后SDS-PAGE电泳Figure 5 SDS-PAGE electrophoresis of purified LASI protein
2.6 LASI重组蛋白鉴定
Western Blot结果如图6所示,实验获得的LASI重组蛋白能与His单克隆抗体发生特异性结合,杂交带位置在22 ku附近,与LASI重组蛋白预期分子质量相符,表明得到了LASI重组蛋白。
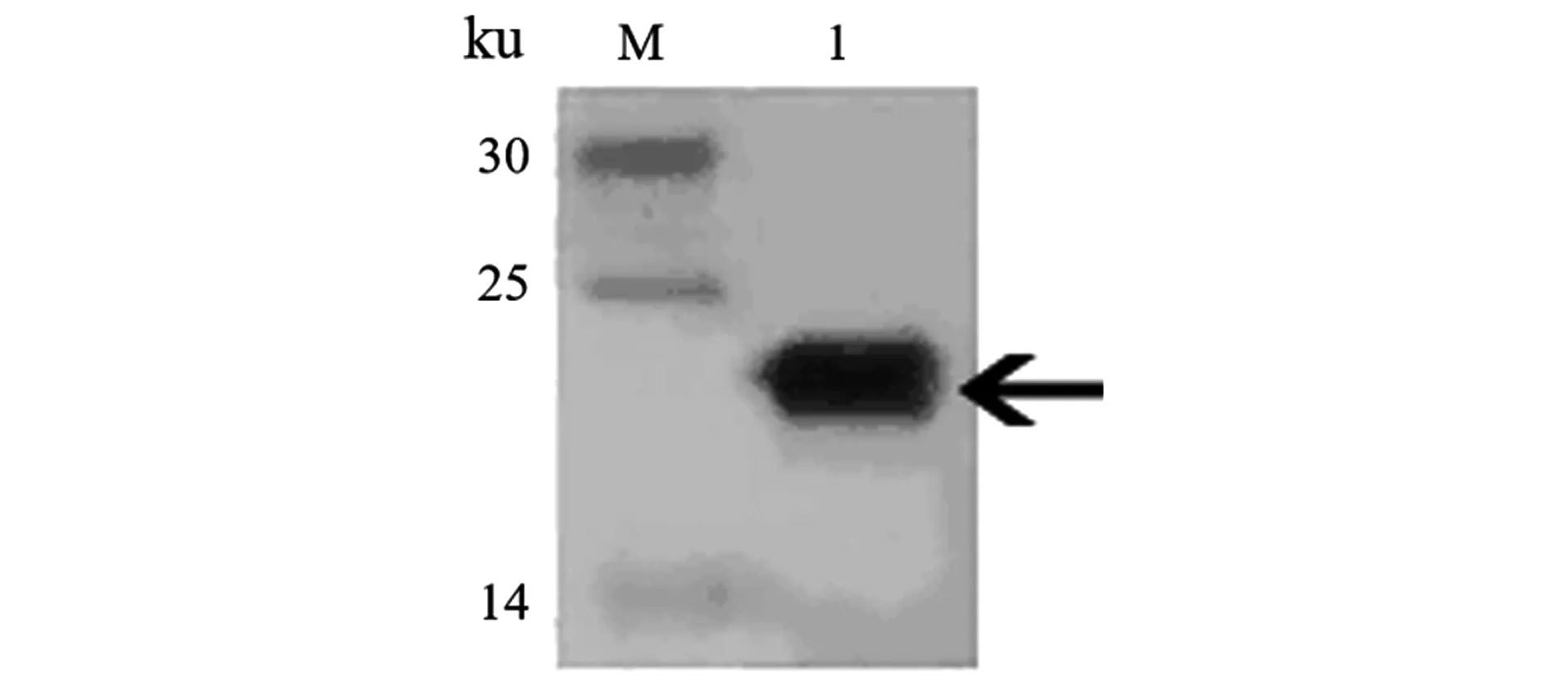
M:Marker;1:纯化后样品。图6 蛋白Western Blot鉴定分析Figure 6 Western Blot analysis of protein
2.7 LASI重组蛋白抑制活性
LASI对猪源α-淀粉酶的抑制活性通过碘-淀粉呈色法进行检测,结果如图7所示。当添加30 μg纯化的LASI时,观察到66%的α-淀粉酶残余活性,LASI对α-淀粉酶的最终抑制比为34%。由此可见,LASI重组蛋白对α-淀粉酶的活性有一定抑制作用。
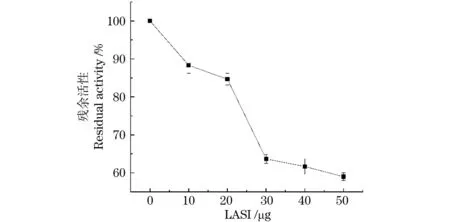
图7 LASI活性检测Figure 7 LASI activity detection
3 讨论与结论
表达载体的选择不仅直接影响重组蛋白的整体表达量,还与重组蛋白的可溶性表达密切相关。然而,不同基因采用何种表达载体并无明显规律性,只能通过实验筛选合适的表达载体。张姝等[18]通过构建pET28a- MDBG-1、pGEX-4T-1- MDBG-1两种表达质粒,发现pET28a的表达量高于pGEX-4T-1,邱淑彬等[19]通过比较两种不同原核表达载体,发现pGEX-6p-1原核表达系统可表达出足量的CP05-GFP蛋白,孙文超等[20]通过构建多种欧洲型PRRSV GP5蛋白原核表达载体,发现GP5蛋白在pGEX-4T-1载体中高效表达。构建pET28a-LASI等4种不同的原核表达重组质粒,诱导表达结果显示:以pET28a为载体时,LASI基因的表达效率较高,产生的LASI重组蛋白最多,因此pET28a为LASI基因的较好表达载体。
基因的表达还受到诱导时间等多种因素影响。适当延长诱导时间能增加蛋白的表达量,但是当蛋白总表达量达到峰值后,即使再延长诱导时间也不会显著增加蛋白表达量。罗非鱼Galectin-8基因在诱导2 h后即达到最佳表达量[21],水芹HSP90基因的最佳诱导时间为4 h[22]。结果表明,在诱导8 h后pET28a-LASI重组蛋白的表达量达到最佳。
诱导剂能够明显提高重组蛋白的整体表达量,但重组蛋白的可溶性表达会因诱导剂浓度偏高而有所降低,导致产生较多的包涵体,不利于后期纯化[23]。水芹HSP90基因在IPTG终浓度为0.2 mmol/L时,即可表达HSP90蛋白,但蛋白的表达量并没有随着IPTG浓度的增加而显示出大幅度的增加[22]。鲤鱼IL-17N基因随着IPTG浓度的变化,其蛋白表达量也没有明显差异[24]。结果表明:在 IPTG终浓度为0.1 mmol/L时,pET28a-LASI开始表达,且IPTG终浓度在0.1~1.0 mmol/L范围内,LASI基因的表达量随着IPTG浓度的增加而增加,但并未呈现线性增长关系,在IPTG终浓度为1.0 mmol/L时LASI基因的表达量达到峰值。
诱导温度在基因表达中也是至关重要的一个因素,过高或者过低的温度均不利于蛋白表达。如罗非鱼Galectin-8在25 ℃时[21]的表达量比37 ℃时的表达量明显减少。罗非鱼NCCRP-1[25]在20 ℃达到最佳的表达效果。水芹HSP90[22]的最佳诱导温度为37 ℃。结果表明,pET28a-LASI的最适诱导温度为27 ℃。
Kunitz类型的α-淀粉酶抑制剂含有两个二硫键,采用Rosetta为宿主细胞,在最优表达条件下,能够成功表达出有生物学活性的LASI重组蛋白,这为后续的抗虫、抗逆与降糖等研究奠定基础。
猜你喜欢
华人时刊(2022年9期)2022-09-06
成都医学院学报(2022年4期)2022-08-19
江西农业学报(2021年4期)2021-04-20
食品与生物技术学报(2021年5期)2021-01-16
华人时刊(2020年15期)2020-12-14
三农资讯半月报(2020年11期)2020-06-21
中国粮油学报(2019年4期)2019-07-12
天然产物研究与开发(2018年6期)2018-07-09
人人健康(2017年11期)2017-06-02
中国当代医药(2015年9期)2015-03-01
