
温带地区不同水分调节行为树种的木质部栓塞特性及其光合响应
2022-08-16 00:38唐华欣杨永清蒋海港李杰林姚银安
生物学杂志 2022年4期
唐华欣,杨永清,蒋海港,刘 伟,李杰林,姚银安
(1.重庆师范大学 生命科学学院,重庆 401331;2.西南科技大学 生命科学与工程学院,绵阳 621000)
水分亏缺是树木经常遭遇的非生物胁迫,树木等维管植物对干旱的抗性取决于维管系统在水分胁迫期间维持水分运输的能力。这可以通过两个策略来实现:第一种为保守型用水策略,将气孔部分关闭维持一定的水势,及时降低叶片蒸腾来实现,其副作用是降低光合速率,这种策略也被称为等水调节对策。第二种为冒险型用水策略,水分亏缺条件下随着叶气饱和水汽压差(VPD)的增加,叶水势降低,从而维持土壤到达叶片的水分驱动力;其光合作用、气孔导度和蒸腾水平仍然保持在较高水平,其副作用是水分散失较快;这种策略也被称为非等水调节对策。一些树种兼有上述两种策略,一些植物甚至在不同阶段采用不同用水策略[1-2]。
当干旱达到一定程度后木质部液流发生空穴化而导致植物导水率下降,产生栓塞现象。木质部出现栓塞后,植物恢复速度减慢,植物死亡率上升。木质部栓塞是理解和预测植物耐旱性的关键特征[3]。植物抗栓塞能力已成为评估植物耐旱性重要指标,这种能力通过栓塞脆弱性曲线(vulnerability curve,VC)来反映,该曲线描述木质部水势与栓塞程度的关系。虽然植物抗栓塞能力作为上述水势调节的水力学指标近年来受到研究者的关注,但是不同用水策略树种的气孔气体交换过程如何响应木质部栓塞作用研究甚少。
研究对温带地区分布较广的7种典型树种,鸡爪槭(Acerpalmatum)、赤皮青冈(Cyclobalanopsisglauca)、北美红杉(Sequoiasempervirens)、辽东冷杉(AbiesholophyllaMaxim.)、欧美黑杨(Populusnigra)、麻栎(Quercusacutissima)和云杉(PiceaasperataMast.)进行研究,采用自然干燥法分析其木质部栓塞特性。对上述7种盆栽植物进行控制水分条件的干旱胁迫处理,根据木质部栓塞VC曲线创造导水率损失率在50%~80% (P50-P80水势)的中度-重度干旱条件,分析叶片气体交换参数和叶绿素荧光参数。揭示不同水分调节树种的栓塞特性,明确在不同栓塞信号条件下两种树种的水分/固碳调节特点,以及相应的光化学响应特征。
1 材料与方法
1.1 材料
鸡爪槭、赤皮青冈、北美红杉、辽东冷杉、欧美黑杨、麻栎和云杉等7种木本植物为研究对象,共选取生长相对一致的7个物种盆栽幼苗共105盆。
1.2 方法
1.2.1 木质部栓塞脆弱性测定
2020年6月至7月,于早晨6:00—7:00进行采样。采用改良的自然干燥法,测定枝条VC曲线。选取各树种间的个体株高、基茎在统计学上没有显著差异,剪取一年生枝条至少10枝后,将枝条立刻插入装有清水的桶中,并且用黑色塑料袋进行遮光带回实验室。使用便携式植物水势压力室测定其水势,冲洗法测定枝条导水率[4]。
1.2.2 干旱胁迫环境中气体交换指标和叶绿素荧光参数测定
7种植物每种植物15盆,将每种植物分成3组,放置在具有钠灯补光系统的温室内进行控制处理;所有植物在胁迫处理前一天的傍晚将试验盆浇透水,使土壤达到饱和含水量,然后在第二天黎明前进行叶水势测定[黎明前Ψleaf与土壤水势(Ψsoil)达到平衡]。并于当日上午9:00—11:00选取每株植株的成熟功能叶第3~8张中部,采用LI-6800全自动便携式光合仪测定植物叶片气体交换和叶绿素荧光参数,对照组始终维持土壤饱和含水量,另两组不再浇水进行持续干旱,每天测定7种植物的叶水势直到导水率损失率在50%或者80%的对应水势;光照环境在11:00遮荫维持散射光条件,温湿度与凌晨相同,使其叶水势在测定期间维持稳定。
试验进行3种水分处理:对照(CK),干旱胁迫(P50:导水率损失50%、P80:导水率损失80%)。每个处理5盆,每盆1株。在胁迫前1 d进行第1次测定,之后不再浇水,在水势达到P50时测定第2次,在水势达到P80时测定第3次,共测定3次。
1.3 数据分析
用R Studio和Microsoft Office Excel 2003对数据进行处理并制作相关图表。
2 结果与分析
2.1 不同树种的导水率损失与栓塞特点
7种温带树种的VC曲线,以水势为自变量,木质部导水率损失百分数(PLC)为因变量,使用单威布尔函数进行回归绘制,计算水势的置信区间时采用bootstrap方法。木质部水势绝对值与植物栓塞程度呈现负相关关系,即当水势降低时,导水率损失率上升,木质部的栓塞程度也会随之增加。
欧美黑杨VC曲线表现为较典型“S”型,说明在轻度干旱条件下,欧美黑杨能维持较低的PLC水平。与欧美黑杨相反,其余6种树种VC曲线均表现出“R”型(图 1)。P50越小表明其达到50%导水率(从而显著影响植物正常功能)的水势越低、干旱程度越严重,因此抗栓塞脆弱性越强。通过表1总结7种树种的P50栓塞脆弱抗性依次为麻栎<辽东冷杉<欧美黑杨<北美红杉<云杉<鸡爪槭<赤皮青冈。
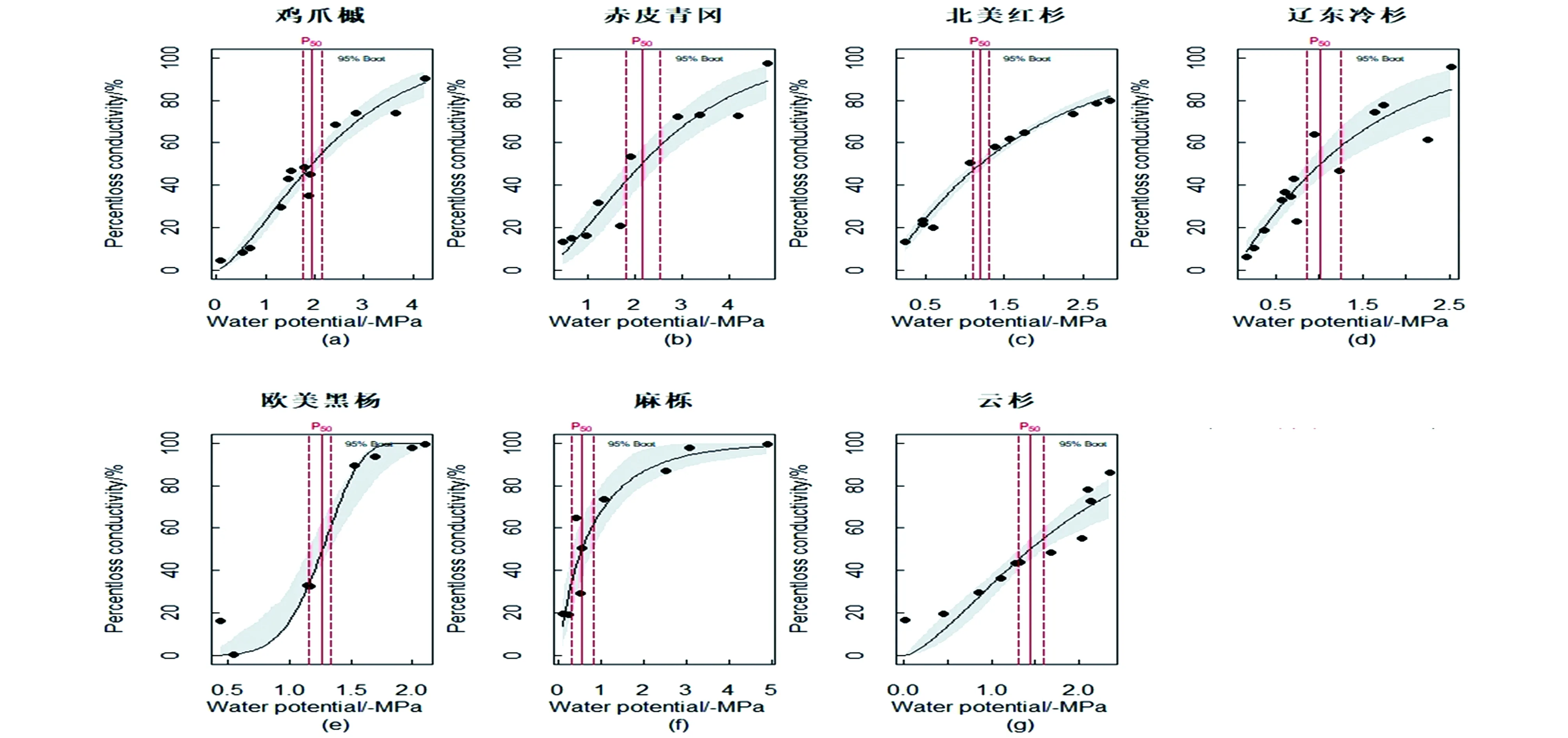
图1 7种温带树种木质部导水率损失百分数PLC和枝条水势的VC曲线Figure 1 VC curve of the percentage loss of water conductivity (PLC) in xylem and branch water potential of 7 temperate tree species
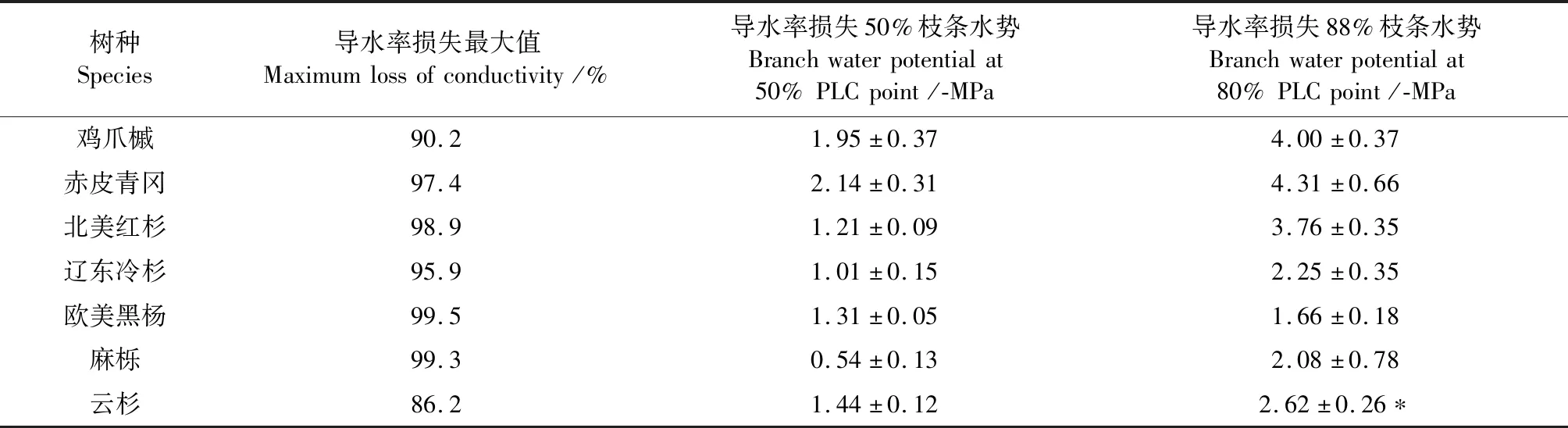
表1 7种温带树种栓塞VC曲线的特征值Table 1 Eigenvalues of embolic VC curves of seven temperate tree species
导水率损失88%是植物生存的临界栓塞水平,可以看出极端栓塞对应的P88水势从大到小依次为欧美黑杨<麻栎<辽东冷杉<云杉<北美红杉<鸡爪槭<赤皮青冈,与P50顺序基本一致。从P88的变化可以看出,赤皮青冈、鸡爪槭和北美红杉都具有较高的耐低导水率、耐极端干旱能力,而欧美黑杨、麻栎两种被子植物最低,两种松科类裸子植物居中。云杉导水率损失率最低仅为86%,显示其对维持生存的忍受能力甚至弱于通常树种88%的导水率。综上7种树种的栓塞抗性顺序为麻栎<欧美黑杨<辽东冷杉<云杉<北美红杉<鸡爪槭<赤皮青冈。
2.2 不同树种在不同栓塞条件下的气体交换特点
从图2可知,7种植物达到木质部导水率降低50%和80%所需时间天数与其抗栓塞能力一致。表明其栓塞特点能够很好地预测植物抗旱性。

图2 7种温带树种P50水势对应达到P50或P80时的天数Figure 2 For 7 temperate tree species,P50 water potential corresponds to the number of days at P50 or P80
从表2可以看出:与CK相比,维持50%木质部导水率(P50)的中度干旱条件下,不同树种的气孔导度(gs)下降程度差异明显。鸡爪槭和辽东冷杉两种植物维持了对照组62%~75%水平,表现出明显的“维持气孔导度”非等水植物特点;相反,赤皮青冈、欧美黑杨、麻栎、北美红杉等gs深度下降达到对照水平的15%~30%,表现出等水植物调节行为特征,尤其是北美红杉;云杉介于上述两类型中间。同时,考察P50条件下的蒸腾速率(E)和净光合速率(A),非等水物种鸡爪槭同样维持了60%以上的水平;而另一个非等水植物辽东冷杉尽管维持了对照75%的气孔导度(gs),然而蒸腾速率(E)和净光合速率(A)和其他等水植物一样,仅为对照组的25%~15%。中间型树种云杉虽然维持了对照组40%~50%的气孔导度(gs)和蒸腾速率(E)水平,净光合速率(A)仍然深度下跌,导致其水分利用效率降低到对照的25%水平。而欧美黑杨和北美红杉提高了50%的水分利用效率(WUEi),其他树种保持了对照组水平。

表2 7种温带树种在不同栓塞条件下的气体交换特点Table 2 Gas exchange characteristics of 7 temperate tree species under different embolization conditions
P80重度干旱条件仅保持20%木质部导水率水平;欧美黑杨叶片凋落进入休眠,多数树种如鸡爪槭、赤皮青冈、麻栎、云杉进一步降低了gs、E,但深度降低了A,导致WUEi大幅度下降;表明生化限制等非气孔因素构成这些物种严重栓塞下的生存障碍。而北美红杉同步降低了gs、E、A,因此保持了对照水平WUEi。辽东冷杉进一步降低了蒸腾速率,却维持或提升了gs、A和WUEi水平。暗示这两种树种在极端栓塞条件下生化水平还没有受到显著影响。
至于胞间CO2浓度(Ci),在P50条件下,7个物种几乎都提高了13%~30%;P80条件下除了北美红杉提高了40%左右外,其他物种提高了70%~100%。
2.3 不同树种在不同栓塞条件下的光化学响应
由表3可知,在P50和P80栓塞水平下,植株的荧光相关参数发生显著变化。暗适应下PSII反应中心完全开放时的最大光化学效率(Fv/Fm)、PSII天线效率(Fv′/Fm′)表征的光照下开放的PSII反应中心激发能捕获效率出现明显下降,趋势十分相似,都随着栓塞程度的加深逐渐降低。Fv′/Fm′被建议用作考察植物在中重度干旱条件下植物生存的表观指标;然而研究中发现P80重度胁迫条件下除麻栎外其他树种与P50没有显著差异,P50与CK相比仅有3个树种有明显差异,显示该指标很难作为栓塞胁迫的预测指标。

表3 7种温带树种在不同栓塞表型下的光化学分析Table 3 Photochemical analysis of 7 temperate tree species under different embolic phenotypes
光照条件下PSII实际的光化学量子效率ΦPSII在4种阔叶植物(鸡爪槭、赤皮青冈、麻栎、欧美黑杨)中随导水率下降而梯度下降,而在3种松科/杉科针叶植物中没有显著变化。相反,所有物种非光化学淬灭系数(NPQ)呈现出随栓塞胁迫而上升;说明随着栓塞胁迫程度的加深,PSII反应中心中的热耗散逐渐增强。
3 讨论
有研究表明,植物木质部的抗栓塞能力与其抗旱能力有密切联系,抗旱能力更高的植物在低水势条件下,木质部仍然具有较高的导水能力[5]。但也有研究认为,这只是植物节约水分的一种耐旱策略,植物木质部栓塞脆弱性与抗旱性之间的相互关系并不明显[6]。研究对植物重度干旱下达到P80的时间点与木质部栓塞脆弱性抗性呈正相关,支持了前一种观点。
自Tardieu等[7]提出等水植物和非等水植物观点以来,对两类植物类型的划分一直存在争议。对等水植物和非等水植物的划分一般通过明确叶片水势和gs的关系来确定,研究同样遵循这个原则来确定,通过P0、P50、P80等3个时点来确定其属于等水还是非等水植物。Klein等[8]总结了70种树种气孔导度降低50%对应的叶水势(Ψgs50),认为植物水分调节对策是一个连续体,而非截然划分成两类,并发现导水率损失点P50水势与Ψgs50成线性正相关,提示可以采用P50条件分析植物气孔的水分调节行为,尽管Andereggl等[9]总结了34种植物发现等水植物的木质部导水率损失值达到50%时其气孔导度显著低于50%。因此,研究将处于P50条件下气孔导度降低幅度40%~60%称为中间型。通过归类已经发表的P50和气孔导度/蒸腾速率损失率关系,支持这个划分方法。
其中较特殊的物种是辽东冷杉,按照定义,本研究和Klein等[8]的报道均将辽东冷杉类植物归为非等水行为调节树种。然而,在P50情况下,尽管物种gs仅降低了25%,其E却降低了80%;同时,发现其他6种植物E与gs保持同步变化,而辽东冷杉的E/gs比例在对照条件下远高于其他树种,干旱处理后大幅度降低,表明该树种气孔调节能力远低于其他物种。这可能与该类冷杉属物种主要生长于光照较低的阴坡,喜欢阴冷湿润气候有关。有研究将中间型树种云杉归入非等水调节行为树种,该物种损失50%的木质部导水率情况下仍然保持了50%的E和40%的gs,然而该物种在P50干旱胁迫下A下降很快,导致其P50干旱条件下WUEi大幅度下降到对照的25%,P80条件下A仅对照的5%,表明该树种在中度和重度干旱情况下都受到严重的非气孔限制,该树种在P50条件下Fv′/Fm′的极显著下降证明了这一点。此外,该树种维持生存的导水率损失阈值PLC甚至弱于通常树种88%临界点,反映了这种在湿润条件下生长的树种对栓塞忍受能力很弱。
鸡爪槭在P50条件下同步维持了较高的gs、E、A,表明在导水率降低了50%条件下维持了碳最大化的冒险型水分调节对策。研究发现北美红杉导水率下降的情况同欧美黑杨一样能够快速响应外界干旱,大幅度降低gs和E,通过提高WUEi来维持生存,反映了北美红杉作为一种高达百米、树龄长至上千年的古老树种的保护型生存策略。同时,北美红杉和赤皮青冈在极端干旱下还具有很强的耐低导水率和耐强栓塞胁迫特点。作为环境胁迫信号指标,两种植物的Fv′/Fm′在P50/P80胁迫条件下变化较小,证实两个树种的叶绿体天线色素仍然保持了正常的光能捕获效率。赤皮青冈具有最强的栓塞脆弱性抗性,作为常绿树种,具有7种典型树种最高的光化学效率(ΦPSII)和较高的A,表明这种以优良材质著称的红木树种在生长和抗旱性方面具有优势,这也是其能千年生存的原因之一。
研究表明树种耐旱能力与其水分调节对策没有很强的相关性,Quero等[10]的研究也证实了这一点。即使同属植物如栎属,也具有耐旱性相反的树种(如赤皮青冈和麻栎)。研究表明麻栎对干旱非常敏感,具有较高的栓塞脆弱性,这可能是导致该树种树干上节较多的原因。
光化学参数中,最大光化学效率(Fv/Fm)反映了PSII的潜在光化学效率与实际光化学效率,ФPSII反映了PSII的电子捕获能力,这也是光合速率的另一种体现[11]。非光化学淬灭系数(NPQ)表征热耗散的大小,可以准确衡量PSII类囊体薄膜的捕光复合体所吸收的不能用于光化学反应而以热能形式耗散的那部分光能,这是为了避免光合器官灼伤的自我保护机制[12]。有研究表明Fv′/Fm′是判断植物生长状况的关键指标[13]。在本研究结果中,当植物处于干旱胁迫逆境后,7种植物的光下最大荧光(Fm′)逐渐减小,光能捕获效率(Fv′/Fm′)逐渐降低,与对照组形成差异。这可能是在干旱胁迫环境下,植物的PSII系统被破坏,开放程度降低,导致原初光能捕获效率下降、电子传递活性减小,从而导致光合效率的降低。前人认为Fv′/Fm′可以作为重度干旱濒死的预测指标,但是基于导水率损失值PLC的研究表明:即使在达到濒临生存危险的处理下其变化不大[14]。而胞间CO2浓度(Ci,非常用的Fv′/Fm′)对木质部栓塞程度具有一致的响应,致死条件下可作为干旱下栓塞预测指标[15]。胞间CO2浓度(Ci)可综合反映植物非气孔限制和光合生化障碍等,植物叶肉细胞光能力降低CO2利用能力降低,表现为Ci升高[16],刘英等[17]的研究在土壤含水量更低的情况下柠条叶片中Ci上升,与我们研究结果一致。考虑到测定栓塞和VC曲线比较繁复,Ci变化可以作为干旱下栓塞程度预测指标。
4 结论
研究表明树木的水分调节方式与其抗栓塞能力和抗旱性关系不大。基于PLC设计抗旱性试验能够清楚反映种间水分调节和抗旱性差异,有助于理解树木适应中度以上干旱强度的生存机制。种间分析表明:胞间CO2浓度(而非常用的Fv′/Fm′)对木质部栓塞程度具有一致的响应,可作为干旱下栓塞预测指标。建议在今后研究植物木质部栓塞脆弱性与抗旱性关系时,应进一步对导管结构和植物生长量、生长特性、生长速率以及外界条件等方面结合起来研究。
猜你喜欢
干旱地区农业研究(2022年1期)2022-01-28
干旱地区农业研究(2021年6期)2021-12-08
植物研究(2021年2期)2021-02-26
农村实用技术(2020年9期)2020-11-19
装备制造技术(2019年12期)2019-12-25
湖北林业科技(2019年4期)2019-09-16
科技创新与品牌(2019年12期)2019-02-06
天津农业科学(2015年11期)2015-12-03
中国果业信息(2015年9期)2015-01-22
浙江农林大学学报(2015年5期)2015-01-08
