
桃小食心虫发育过程中蜕皮激素滴度动态及甲氧虫酰肼干扰滞育的作用
2022-08-13 08:02李永丽闫作炳尹新明雷振山
昆虫学报 2022年7期
李永丽, 闫作炳, 尹新明, 雷振山,*, 周 洲,*
(1. 信阳农林学院农学院, 河南信阳 464000; 2. 河南农业大学植物保护学院, 郑州 450002)
桃小食心虫Carposinasasakii属鳞翅目(Lepidoptera)果蛀蛾科(Carposinidae),在国内24个省市均有分布,是为害面积最大、发生最普遍的果树食心害虫(王鹏等, 2011)。桃小食心虫卵孵化后,幼虫会很快钻蛀果实,以桃、苹果、枣、梨和山楂为寄主的桃小食心虫形成了兼性滞育,滞育受光周期的影响;而以杏为寄主的桃小食心虫形成了严格的专性滞育,滞育基本不受光周期影响(花保祯等, 1998)。杏生物型在发生时间(季节)上与发生期较晚的(桃、苹果、枣、梨和山楂)生物型之间出现了非常大的遗传分化,在生殖上已经出现了明显的时间隔离(候月利, 2003)。光周期是调控其滞育发生的关键因子(黄可训等, 1976),在长光照(光周期14L∶10D)条件下脱果的老熟幼虫在土壤中结长茧,随后在长茧内化蛹,蛹期结束后,桃小食心虫蛹移动至长茧前端羽化成虫;在短光照(光周期12L∶12D)条件下脱果的老熟幼虫在土壤中结圆茧滞育,圆茧内一直保持幼虫状态,到滞育解除幼虫才咬破圆茧出土,在土中重新结成长茧化蛹,羽化后繁殖下一代(张乃鑫等, 1977; 李锐等, 2014)。
蜕皮激素(20-hydroxyecdysone, 20E)是昆虫发育过程中最重要的激素之一,在昆虫滞育的调节过程中发挥关键作用(Guoetal., 2021)。研究表明,昆虫种类不同其滞育调节过程20E滴度变化模式亦不尽相同。例如,麦红吸浆虫Sitodiplosismosellana在滞育前20E有较高滴度,进入滞育期显著降低维持在低水平,滞育期后期显著升高出现两个明显高峰(成卫宁等, 2009);舞毒蛾Lymantriadispar滞育诱导和维持过程中20E的作用是必要的,滞育解除期间20E的滴度降低(Denlinger and Lee, 1997)。20E在桃小食心虫滞育调控中的作用尙待明确。本研究首先对桃小食心虫滞育与非滞育两种发育过程中20E滴度变化进行测定,明确20E滴度在两种发育过程中的关键差异;然后使用蜕皮激素类似物甲氧虫酰肼(methoxyfenozide)影响桃小食心虫体内20E滴度,以期达到干扰桃小食心虫滞育的目的,为桃小食心虫防治新技术的开发奠定基础。
1 材料与方法
1.1 供试虫源
桃小食心虫采自河南省洛阳市洛宁县罗岭乡山楂园(34°37′19″N, 111°39′61″E,海拔650 m)。试虫于人工气候箱中使用山楂果连续饲养12代,温度25±1℃,相对湿度70%±5%,长光照条件光周期为15L∶9D,短光照条件光周期为12L∶12D。
1.2 主要试剂及仪器
98%甲氧虫酰肼原药(江苏昌瑞药业),20E(上海生工生物工程有限公司),昆虫20E ELISA试剂盒(江苏酶免MM-133301);其他分析纯试剂(天津四友)。超声波破碎仪(宁波新芝650E),MultiskanTMFC酶标仪(Thermo Scientific)。
1.3 虫体收集处理与20E滴度测定
收集不同发育阶段和不同滞育状态的虫体,包括长(短)光照幼龄到初脱果幼虫;长(短)光照脱果第1, 2, 3, 4, 5, 6, 7和8天蛹和幼虫;保存在8℃沙土中的短光照脱果滞育第10, 40, 70, 90, 120, 140及160天滞育圆茧内的幼虫,通过液氮速冻后置于-80℃超低温冰箱冻存备用。
取保存幼虫置于1.5 mL离心管中称重,脱果老熟幼虫3头合并称重,2龄及3龄幼虫每组10~12头合并称重。在每支离心管中按单组虫重的10倍加入蒸馏水稀释,使用玻璃研棒快速初步研磨虫体。装载初步研磨过虫体的离心管置于冰中保持0℃低温,超声波细胞破碎仪变幅杆伸入离心管液面,选择功率50 W、振幅50 Hz、每工作5 s间隔暂停3 s,超声破碎5 min;在4℃下2 500 r/min离心20 min,取10 μL上清液按照试剂盒说明书测定20E浓度。
1.4 甲氧虫酰肼处理脱果幼虫及结茧统计
甲氧虫酰肼原药先用二甲基亚砜溶解,涡旋震荡助溶均匀后再稀释使用。点滴法处理短光照脱果幼虫试验中,用丙酮(按H2O∶丙酮=1∶9, v/v)稀释甲氧虫酰肼至7个浓度梯度(0.5, 1, 2, 5, 7.5, 10和15 mg/mL),现配现用;喷沙法处理短光照脱果幼虫试验中,再用H2O(按二甲基亚砜∶H2O=1∶9, v/v)稀释甲氧虫酰肼至7个浓度梯度(0.05, 0.1, 1, 2, 5, 7.5和10 mg/mL),现配现用;仅点滴丙酮(按H2O∶丙酮=1∶9, v/v)作为对照。点滴法中,试虫于冰袋上冷冻麻痹,用移液枪将溶液点在前胸背板上,每头点滴1 μL,每组10头试虫,3组重复;试虫置于1 000 mL容量的一次性快餐盒中,铺1.5 cm厚沙子,含水量5%。正常情况下,试虫脱果2 d后均能够完成结茧,因此试验中每2 d筛沙子统计结茧结果,对结茧结果不会产生影响。喷沙法中,1 000 mL容量的塑料盒中铺1.5 cm厚沙子,含水量5%;用喷壶在餐盒的沙子表面均匀喷洒甲氧虫酰肼水溶液,单组喷药量控制在2 mL;脱果试虫每组10头,3组重复;仅喷清水组作为对照,每2 d筛沙子统计结茧结果。
1.5 甲氧虫酰肼和20E喷沙处理后桃小食心虫体内20E滴度的测定
选择5 mg/mL甲氧虫酰肼喷沙组与0.5和1 mg/mL 20E喷沙组,两处理组结茧比例处于同一水平,分别测定这两组中圆茧、畸形茧和不结茧试虫虫体的20E滴度。清水喷沙组作为对照。
1.6 甲氧虫酰肼喷沙处理对桃小食心虫圆茧滞育解除的影响观测
收集1.5节甲氧虫酰肼低浓度0.1 mg/mL喷沙处理组中所结的圆茧44个,甲氧虫酰肼高浓度5 mg/mL喷沙处理组中的圆茧19个,同期对照组中的圆茧60个,圆茧储存于6±2℃低温下120 d。然后取出开始升温过程,依次置于12±1℃, 16±1℃和20±1℃ 各2 d,最终保持在温度25±1℃,相对湿度70%±5%,光周期15L∶9D条件下。圆茧中幼虫经上述滞育解除处理后,经结长茧后羽化出成虫,统计各组羽化时间和羽化比例,羽化时间从取出升温日期开始计算。
1.7 数据分析
不结茧和结畸形茧都判定不能正常进入滞育,记录为死亡。 实验数据采用Excel 2016和SPSS 19.0 等软件进行统计学分析,组间差异采用Duncan氏多重比较。
2 结果
2.1 滞育和非滞育发育过程中桃小食心虫体内20E滴度动态变化
桃小食心虫在短光照(光周期12L∶12D)和长光照(光周期15L∶9D)条件下分别进行的是滞育和非滞育发育过程。两种发育过程中,幼虫期20E滴度均是从幼龄到5龄脱果每龄逐渐降低,脱果时达到最低,但是滞育发育脱果幼虫20E滴度显著低于非滞育发育脱果幼虫滴度(P<0.05)。随后虫体20E滴度升高,1 d后非滞育发育幼虫结成长茧,滞育发育幼虫结成圆茧,但是非滞育发育的幼虫20E滴度极显著高于同期滞育发育幼虫(P<0.01)(图1)。长光照下,幼虫20E滴度从2龄到脱果逐渐降低,从2.402 ng/g降低到0.473 ng/g;随着进入蛹期20E滴度迅速上升,长茧1 d达到0.922 ng/g,在蛹期持续保持较高滴度(0.652~1.035 ng/g),并且在7 d后蛹末期继续升高(1.217 ng/g)。短光照下,幼虫的20E滴度从2龄2.757 ng/g到脱果逐渐降低到0.254 ng/g;在进入滞育圆茧(如图1实线所示)的前几天,虫体20E滴度缓慢上升,第4天达到较高滴度0.656 ng/g,第6天滴度达到低值0.318 ng/g,第8天再次达到较高滴度0.790 ng/g;10 d后进入滞育稳定阶段,随着滞育时间延长缓慢降低,70 d以后达到滞育结束状态,90 d达到最低滴度0.424 ng/g,圆茧转移到25℃条件下很快可以出土活动;随后20E滴度逐渐上升,160 d 20E滴度升高到1.221 ng/g。
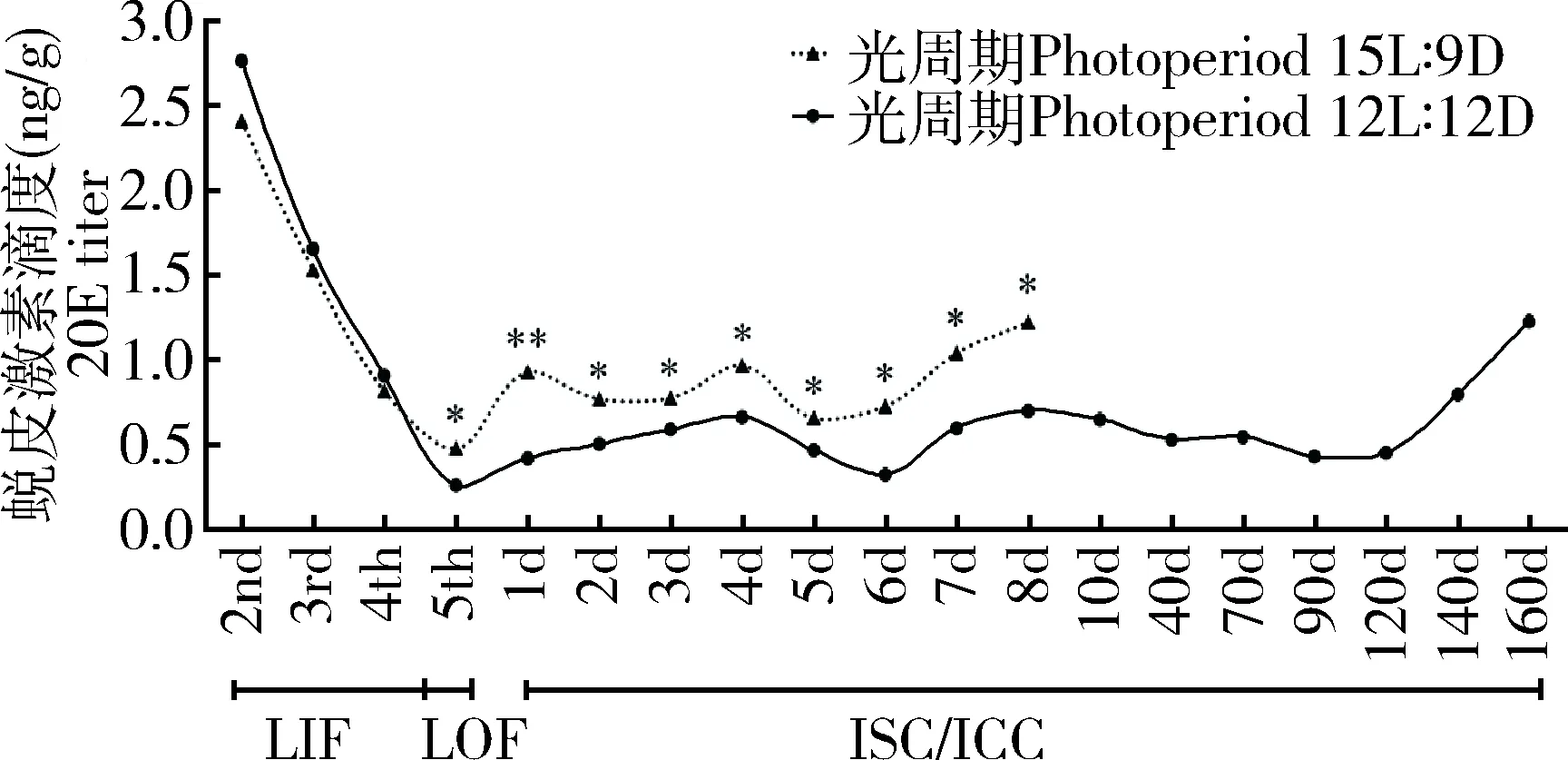
图1 桃小食心虫滞育和非滞育幼虫发育过程中20E滴度的动态Fig. 1 Dynamics of the 20E titer in Carposina sasakii larvaeduring diapause and non-diapause developmentLIF: 蛀果期幼虫Larvae in fruit; LOF: 脱果幼虫Larvae escaping off fruit; ISC: 幼虫进入长茧后Larvae entering long cocoon; ICC: 幼虫进入圆茧后Larvae entering round cocoon; 2nd: 2龄幼虫2nd instar larva; 3rd: 3龄幼虫3rd instar larva; 4th: 4龄幼虫4th instar larva; 5th: 5龄幼虫5th instar larva; 1d-160 d: 分别为进入滞育后1-160 d (1-160 d, respectively, after entering diapause). 图中数据为平均值±标准误; 下同。星号和双星号分别表示同一时间点的滞育和非滞育发育幼虫体内20E滴度差异显著(P<0.05)和极显著(P<0.01)(t检验)。Data in the figure are mean±SE. The same below. The asterisk and double asterisk indicate significant difference (P<0.05) and extremely significant difference (P<0.01)(t-test), respectively, in the 20E titer of diapause and non-diapause developing larvae at the same time point.
2.2 甲氧虫酰肼点滴处理对桃小食心虫幼虫发育和结茧的影响
用甲氧虫酰肼通过点滴法处理短光照脱果桃小食心虫幼虫,发现试虫出现的表型与甲氧虫酰肼点滴剂量相关。低剂量处理后结畸形茧(茧外形不圆、包裹松散,或包裹不完全)(图2: A)比例较少,随着点滴剂量的增加出现畸形茧比例增高,并且出现反复结茧现象,6 d后结茧稳定下来,最终统计结畸形茧和不能结茧(图2: A)的比例。畸形茧和不能结茧14 d时均自然死亡,在第6天时统计死亡率时将畸形茧和不能结茧记作死亡。点滴法测定甲氧虫酰肼对桃小食心虫脱果幼虫第6天的毒力学回归方程为y=0.2826x+3.0163,R2=0.9559,LD50=7.039 μg/头。
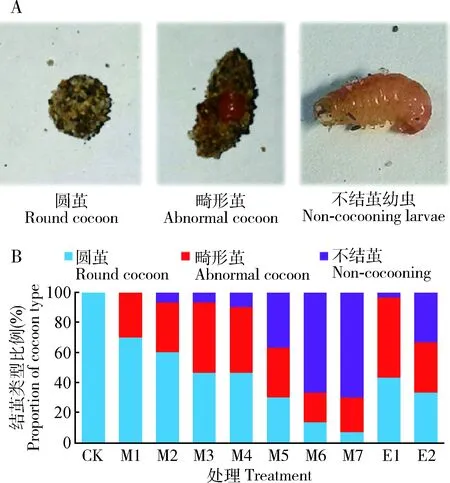
图2 甲氧虫酰肼和20E喷沙处理对桃小食心虫结茧的影响Fig. 2 Influence of methoxyfenozide and 20E sprayed onthe sand surface on the cocoon formationof Carposina sasakiiA: 典型圆茧、畸形茧和不结茧幼虫Typical round cocoon, abnormal cocoon and non-cocooning larvae; B: 结茧类型比例Proportion of cocoon type. CK: 为清水喷沙 Pure water sprayed on the sand surface; M1-7: 分别为0.05, 0.1, 1, 2, 5, 7.5和10 mg/L甲氧虫酰肼喷沙0.05, 0.1, 1, 2, 5, 7.5 and 10 mg/mL methoxyfenozide sprayed on the sand surface, respectively; E1-2: 分别为0.5和1 mg/mL 20E喷沙0.5 and 1 mg/mL 20E sprayed on the sand surface, respectively.
2.3 甲氧虫酰肼喷沙处理对桃小食心虫结茧的影响
由于脱果幼虫在沙表面有长时间爬行和咀嚼沙子结茧的行为,试验尝试了喷沙法干扰桃小食心虫短光照脱果幼虫滞育结茧行为,同时对比了0.5 mg/mL和1 mg/mL 20E喷沙效果。统计了处理第6天的圆茧、畸形茧及不能结茧的数量,结果见图2所示。喷沙组甲氧虫酰肼和20E均随施用浓度的升高,结正常圆茧比例(图2: B)明显减少,结畸形茧(图2: B)和不结茧(图2: B)的试虫数量明显增多。随着药剂浓度的增加,畸形茧比例先升高后降低,1 mg/mL甲氧虫酰肼处理组畸形茧比例最高;浓度超过1 mg/mL后,随浓度提高不结茧比例增多。这说明结畸形茧是结茧受影响较轻的表型,不结茧是结茧受影响较重的表型。相同施用量的甲氧虫酰肼和20E喷沙组比较,20E造成不结茧的比例稍高,这说明20E对桃小食心虫的活性更高, 5 mg/mL甲氧虫酰肼喷沙处理组和 1 mg/mL 20E喷沙处理组不能正常结茧的比例处于同一水平。
2.4 外源甲氧虫酰肼和20E对桃小食心虫体内20E滴度的影响
试验测定了5 mg/mL甲氧虫酰肼和 1 mg/mL 20E两喷沙处理组中虫体20E的滴度,结果统计见图3,不能结圆茧的比例分别为70.0%和66.7%,其中结畸形茧的幼虫体内20E滴度分别显著升高16.3%和143.0%,不能结茧的分别显著升高149.3%和278.6%。与对照组正常圆茧内虫体相比,甲氧虫酰肼和20E处理组中结正常圆茧虫体处于同一水平,结畸形茧和不能结茧试虫的20E滴度处理组显著高于对照组(P<0.05),结果说明外源甲氧虫酰肼和20E造成了虫体内20E的滴度的异常升高,这可能是引起结茧异常,甚至是不能结茧的一个原因。同时试验还发现,20E喷沙组中结畸形茧和不能结茧试虫的20E滴度显著高于甲氧虫酰肼喷沙组相应虫体(结果未展示)。

图3 外源甲氧虫酰肼和20E对桃小食心虫幼虫体内20E滴度的影响Fig. 3 Influence of exogenous methoxyfenozide and20E on the 20E titer in Carposina sasakii larvae喷清水的作为对照组(CK);两处理组分别为5 mg/mL甲氧虫酰肼喷沙处理组和 1 mg/mL 20E喷沙处理组。星号表示同对照圆茧内虫体20E滴度差异显著(P<0.05, t检验)。In the control group (CK) pure water was sprayed on the sand surface, and in the two treatment groups 5 mg/mL methoxyfenozide and 1 mg/mL 20E were sprayed on the sand surface, respectively. The asterisk indicates significant difference from the 20E titer in the control round cocoon (P<0.05, t-test).
2.5 甲氧虫酰肼处理对桃小食心虫滞育解除的影响
对照组和甲氧虫酰肼处理组中的圆茧,经历相同低温存储和同样升温过程,最终保持在温度25±1℃统计滞育解除成虫羽化时间。对照组中第1头成虫羽化时间需要29 d,46 d之后不再有成虫羽化出现,共羽化成虫49头,成功解除滞育羽化率为81.7%。低浓度(0.1 mg/mL)甲氧虫酰肼喷沙处理组圆茧经解除滞育,第1头成虫羽化时间是57 d,80 d解除滞育试虫的羽化率仅为20.8%,后续清理发现一半试虫已于圆茧内死亡;高浓度甲氧虫酰肼5 mg/mL喷沙处理组中圆茧没有出现成虫羽化,试虫全部于圆茧内死亡(表1)。这说明桃小食心虫幼虫接触到甲氧虫酰肼后,虽有一定比例能够结圆茧滞育,但相当多的幼虫后续的滞育发育依然受到了影响,能够成功完成滞育的比例减少。

表1 不同浓度甲氧虫酰肼喷沙处理对桃小食心虫滞育解除的影响Table 1 Effect of different concentrations of methoxyfenozide sprayed on the sand surfaceon the diapause termination of Carposina sasakii
3 讨论
桃小食心虫幼虫经历了长光照和短光照后,脱果老熟幼虫分别进行非滞育和滞育发育。两种发育过程幼虫脱果之前,各龄期20E滴度没有差异,变化趋势均随着龄期增大逐渐降低;但脱果之后,长光照脱果非滞育老熟幼虫20E滴度(0.473 ng/g)显著高于短光照脱果注定滞育的老熟幼虫(0.254 ng/g)。 非滞育老熟幼虫结长茧随后进入蛹期,虫体20E的滴度显著升高,在蛹期保持较高滴度(0.652~1.217 ng/g);注定滞育老熟幼虫随后结圆茧在进入滞育初期,最初4 d 20E滴度缓慢上升, 第4天达到第1个峰值0.656 ng/g,随后降低后再升高,在第8天达到第2个峰值0.790 ng/g;进入圆茧10 d后,逐渐进入稳定滞育阶段,20E滴度逐渐降低保持在0.552 ng/g左右低水平;70 d达到滞育能够解除的时间,直到第90天达到滞育期间最低滴度(0.424 ng/g),圆茧转移到25℃条件下很快可以出土活动;随后20E滴度逐渐上升,160 d 20E滴度升高到1.221 ng/g(图1)。本研究说明桃小食心虫进入滞育圆茧时需要低水平的20E滴度,在滞育过程中 20E滴度维持在低水平,在生殖变态发育阶段需要较高滴度,20E滴度变化模式与蜕皮和变态相吻合。
在滞育期里20E滴度保持低水平,这可能是滞育昆虫的共性,但不同昆虫滞育过程中20E滴度变化模式不尽相同。竹螟Omphisafuscidentalis(Singtripopetal., 2000)和麦红吸浆虫(成卫宁等, 2009)在滞育期体内的20E滴度也保持较低水平,这与桃小食心虫在此过程中保持20E滴度的模式相似。桃小食心虫在滞育前调控其体内20E滴度处于极低的水平,在滞育末期调控20E滴度迅速上升,施加外源20E类似物甲氧虫酰肼可阻止进入圆茧滞育(表1);这些结果说明,脱果时低水平的20E滴度是桃小食心虫进入滞育的条件之一,避免进入变态发育程序,高滴度的20E是桃小食心虫解除滞育的条件之一。与此类似,对柑橘大实蝇Bactroceraminax的研究发现,高滴度的20E可以终止其蛹滞育(Chenetal., 2016)。但是, Denlinger和Lee(1997)的研究表明,一定浓度的20E对舞毒蛾滞育诱导和维持是必要的,并且解除滞育期间20E的滴度降低,与桃小食心虫解除滞育有所不同;这其中原因可能是舞毒蛾是1龄幼虫滞育,滞育解除后需继续生长,但桃小食心虫是老熟幼虫滞育,解除滞育后将进行化蛹变态发育。Sun等(2015)研究者通过浸叶的方式使小菜蛾Plutellaxylostella摄入一定量的20E,发现对小菜蛾的取食、发育和繁殖均有不利影响,摄入20E可能会干扰综合信号通路,从而导致异常的生物合成或代谢。本研究中甲氧虫酰肼点滴桃小食心虫脱果幼虫能够干扰滞育圆茧的形成,甲氧虫酰肼对桃小食心虫幼虫的LD50值为7.039 μg/头;甲氧虫酰肼和20E喷沙处理均可有效干扰滞育圆茧的形成,相同浓度的20E比甲氧虫酰肼表现出的干扰活性更高(图2)。不能结滞育圆茧的比例随着甲氧虫酰肼点滴量的增加而增多,表现出结非滞育的长茧、结畸形茧、反复结茧、分泌红色体液、失去结茧能力、虫体快速缩小以及死亡等症状(图2),激素分析表明不能结滞育圆茧与虫体20E滴度显著升高相关(图3)。甲氧虫酰肼处理后虽有一定比例能够结圆茧,观察这些圆茧后续滞育解除,发现相当多的幼虫后续的发育依然受到了影响,能够成功完成滞育的比例减少(表1);一些虽然能羽化出成虫,但其发育速度与对照组野生型相比也有较大差异,这一定程度上将影响其匹配自然寄主植物的物候期。但在本研究中,仅测定了外源甲氧虫酰肼和20E处理后内源20E滴度的变化,20E之外的其他生化指标还有待进一步的试验来明确,有望解释干扰作用的内在原因。
目前,桃小食心虫的化学防治主要以树上防治为主,地下防治为辅(蔡明等, 2014)。树上防治以20%毒死蜱微囊悬浮剂500倍液的应用效果最优(张元臣等, 2018),使用4%高氯·甲维盐微乳剂(高越等, 2017)及氯虫苯甲酰胺水分散粒剂等(赵楠等, 2014)药剂对桃小食心虫进行防控也有很好的防控效果。树下防治应用毒死蜱乳油或氯氰-毒死蜱乳油,能够很好地抑制桃小食心虫老熟幼虫破冬茧出土(黄鹏等, 2013)。成虫性诱剂的使用和果实套袋也是较有效的防治方法,但是成虫性诱集的时效性和成本较高是一个突出问题(赵君瑾等, 2008; 薛艳花等, 2010)。本研究通过对比桃小食心虫滞育与非滞育幼虫体内的20E滴度差异,选择用环境友好、对人畜低毒无害的甲氧虫酰肼在室内成功地对桃小食心虫滞育进行了干扰,尤其是甲氧虫酰肼喷沙处理只需极低的使用量就达到较高的致死率和发育进程改变。本研究与之前树下防治的创新点是防治时间提前到老熟幼虫脱果时期,且药剂安全性高,为下一步田间应用甲氧虫酰肼干扰桃小食心虫滞育,达到控制其种群数量奠定了良好基础。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
动漫界·幼教365(大班)(2021年10期)2021-10-13
华侨大学学报(自然科学版)(2021年4期)2021-07-30
中国医药科学(2021年7期)2021-05-11
西北园艺·果树(2021年1期)2021-02-21
意林·少年版(2020年14期)2020-08-02
河北果树(2020年2期)2020-05-25
作文评点报·低幼版(2016年26期)2016-08-17
浙江柑橘(2016年2期)2016-03-11
奥秘(2014年10期)2014-10-17
