
栲树叶片中不同类型钙含量对海拔梯度和土壤因子变化的响应
2022-08-12 08:05柏雨萱刘丹萍邢郑瑜林仁忠廖晓丽靳少非郑德祥
植物资源与环境学报 2022年4期
柏雨萱, 刘丹萍, 邢郑瑜, 苏 仪, 林仁忠, 廖晓丽, 靳少非, 郑德祥,①
(1.福建农林大学林学院, 福建 福州 350002; 2.顺昌县国有林场, 福建 顺昌 353200; 3.闽江学院地理与海洋学院, 福建 福州 350108)
钙作为植物生长必需的营养元素之一,在植物生长发育和适应环境过程中发挥着至关重要的作用[1]。在植物细胞中,钙主要以水溶性钙、果胶酸钙、草酸钙、硝酸钙和氯化钙、磷酸钙和碳酸钙、硅酸钙6类形式存在。植物叶片是钙的存储器官之一,叶片对环境因子的变化较为敏感[2,3],导致叶片细胞中的钙在含量和类型上变化各异且具有特异性[4]。研究结果[5,6]表明:在不同种类的植物叶片细胞中钙的类型有明显差别;苏志孟等[7]认为,堇菜(ViolaverecundaA.Gray)叶片细胞中草酸钙晶体的密度与束缚水含量具有显著正相关关系,说明在干旱条件下堇菜叶片细胞可能通过调节草酸钙晶体的密度应对干旱胁迫;刘剑锋等[8]的研究结果表明:梨(Pyrussp.)的果实用质量体积分数6%CaCl2浸泡后,果实的磷酸钙和草酸钙含量均不同程度提高。可见,植物细胞中存储的不同类型的钙对植物适应环境异质性有重要作用,在不同逆境条件下植物叶片细胞中各类型的钙可通过调整其含量来适应生境的变化[4]。
在实际营林生产过程中,海拔是制约树种分布与造林培育的重要环境因子,海拔变化可引起气温、光照和水分等生境因子的改变,进而影响植物的生长和生理代谢过程[9,10]。植物叶片中的钙含量也可随海拔变化而改变,其中海拔差异引起的气温变化是影响植物钙吸收能力的重要原因之一[11]。除海拔因子外,土壤作为植物生长的基本载体,其养分含量及含水量等因子对植物的生长同样存在重要影响[12,13],其中,土壤理化性质变化对植物钙素含量也存在一定的影响[14,15],例如:土壤pH值降低可能会抑制植物对钙素的吸收;土壤有机碳、全氮、全钙和全磷含量与叶片钙含量存在显著正相关。由于钙在植物的生长发育和环境适应过程中具有重要作用,目前已有不少学者对植物叶片中钙含量及不同类型钙的比例及其环境影响因子进行了研究,但研究目标主要集中于喀斯特等高钙土壤环境地区[4,5,16]。然而,钙作为第二信使参与植物体诸多生理代谢过程[1],其与环境因子间的相互作用机制尚不明确,且在其他非钙质土壤中不同类型钙对植物的作用与高钙土壤是否相同,均有待深入研究。此外,酸化土壤对土壤系统中钙的平衡产生影响,进而影响植物的正常生长[17],使得亚热带森林土壤中的钙可能逐渐取代氮素成为限制该地区林木生长的关键因子[15,18],那么,在亚热带非钙质土壤环境中植物能否通过调整叶片中各类型钙的分配策略来适应酸性土壤环境尚需进一步的深入研究。
栲树(CastanopsisfargesiiFranch.)为中国亚热带常绿阔叶林的常见建群种之一,主要分布在长江以南各地。目前对栲树的研究主要集中在土壤微生物群落结构[19]、林分结构[20]、幼苗对氮素的响应[21]、种子性状变异[22]、叶片功能性状对增温和施氮的影响[23]及优势群落的结构稳定性[24]等方面,而关于栲树生长和代谢与钙元素吸收、积累的关系以及植物在不同环境下不同类型钙的分配策略尚不清楚。
为此,作者在福建郭岩山县级森林生态自然保护区内,按照海拔设置5个样地,通过分析不同样地中栲树叶片不同类型钙含量和土壤理化指标的变化特征,探究各类型钙含量与海拔和土壤理化指标的关系,揭示栲树叶片中各类型钙的积累水平对海拔和土壤因子变化的响应规律,探讨栲树叶片通过调整不同类型钙的分配策略适应非钙质土壤环境的作用机制,以期为中国亚热带地区山地造林以及林分施钙措施的制定提供理论依据。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于福建省顺昌县郭岩山县级森林生态自然保护区,地理坐标为东经118°00′~118°30′、北纬27°06′~27°18′,地处福建省南平市与三明市的交接地带,最高海拔1 383.7 m。该保护区地处亚热带地区,属海洋性季风气候;年均气温19 ℃,四季分明;年降水量1 600~1 800 mm,常年雨量充沛。主要土壤类型为黄红壤(海拔500~700和900~1 000 m)和黄棕壤(海拔700~900 m),土壤厚度多为0~20 cm。
研究区内植被类型丰富且具有明显的垂直分布特征。海拔500 m以下区域多为杉木〔Cunninghamialanceolata(Lamb.)Hook.〕人工林;海拔500~1 000 m区域以栲树、木荷(SchimasuperbaGardn.et Champ.)和马尾松(PinusmassonianaLamb.)混交林为主,其中栲树为主要建群种;海拔1 000 m以上区域多为肿节少穗竹〔Oligostachyumoedogonatum(Z.P.Wang et G.H.Ye)Q.F.Zhang et K.F.Huang〕及小乔木。
1.2 研究方法
1.2.1 实验设计和样品采集 在2020年11月,依据随机性和独立性的样地划分原则,在研究区内海拔500~900 m区域,以100 m为海拔梯度设置5个样地,各样地光照、坡度、地形和地貌条件等基本一致,坡向均为西南朝向。各样地基本概况见表1。
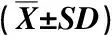
表1 供试栲树样地的基本概况
在每个样地中划分5个自然条件基本一致的标准样方,面积均为30 m×30 m。在每个标准样方中随机选取生长良好、树形基本一致的野生成年栲树5株,在各样株的树冠外围采集完全成熟、无病虫害、无损坏的健康叶片20~40枚,置于密封袋中并用便携式冰箱带回实验室,于-80 ℃超低温冰箱中保存,用于叶片钙含量测定。
在每株样株的树冠下,去除地表的凋落物,以树干基部为中心沿地表径流方向,在上坡位和下坡位各采集表层(0~20 cm)土壤300~500 g,同一样地采集的土壤样品混合均匀后带回实验室,用于土壤理化指标测定。
1.2.2 叶片中各类型钙的提取和含量测定 参照Ohta等[25]的方法,依次加入不同提取剂进行叶片中各类型钙的提取,每个样地设置5个重复样品,同时以空白样品(未放叶片)为对照进行结果校正。将叶片磨碎后分别称取约0.5 g,精密称量后加入7 mL提取剂,于25 ℃水浴中恒温振荡1 h,在4 ℃下4 000 r·min-1离心10 min,收集上清液,重复提取2次;合并上清液并使用定量滤纸过滤,用体积分数5%HCl溶液定容至25 mL,摇匀后即为供试样液,残渣备用。其中,硝酸钙和氯化钙以体积分数80%C2H5OH溶液作为提取剂;水溶性钙以蒸馏水作为提取剂;果胶酸钙以浓度1 mol·L-1NaCl溶液作为提取剂;磷酸钙和碳酸钙以体积分数2%CH3COOH溶液作为提取剂;草酸钙以体积分数0.6%HCl溶液作为提取剂。
提取后的残渣用电热板加热挥干溶剂,参照齐清文等[5]的提取步骤,用V(HNO3)∶V(HClO4)=4∶1混合液作为消化液,消化提取叶片中的残渣钙(硅酸钙),冷却后用体积分数0.2%HNO3溶液定容至25 mL,摇匀后即为残渣钙样品溶液。同时消煮空白样品(未放叶片)进行结果校正。
上述样品溶液用ZEEnit 700P原子吸收分光光度计(德国Analytik Jena公司)在波长422.7 nm下进行测定。
1.2.3 土壤理化指标测定 在采集土壤样品时用Model SK-250WP温度传感器(日本SATO公司)测定土壤温度;采用烘干法[26]测定土壤含水量。将自然风干的土壤研磨后过孔径2 mm筛,称取10 g土壤样品,加入25 mL超纯水并剧烈搅动,静置30 min后利用ST 2100玻璃电极pH计(美国OHAUS公司)测定土壤pH值。
将自然风干的土壤研磨后过孔径0.149 mm筛,称取0.2 g土壤样品,采用HNO3-HClO4法[27]消煮后,用PerkinICP-MS电感耦合等离子体发射光谱仪(美国PERKINELMER公司)测定土壤全磷、全钾和全钙含量。称取前述土壤样品0.2 g,用锡舟(4 mm×4 mm×11 mm,德国ELEMENTAR公司)包样后,用Vario MAX碳氮元素分析仪(德国ELEMENTAR公司)测定土壤全碳和全氮含量。每个样地设置5个重复样品,同时设置空白样品(未放叶片)进行结果校正。每份样品各指标均重复测定3次,结果取平均值。
1.3 数据处理和统计分析
用EXCEL 2010和SPSS 23.0软件对数据进行统计分析;采用单因素方差分析法(one-way ANOVA)分析不同样地间栲树叶片各类型钙含量的差异;采用Pearson法分析叶片各类型钙含量间的相关性。
以海拔和土壤基本理化性质为环境因子,以叶片各类型钙含量为物种因子,使用CANOCO 5软件进行RDA约束排序以分析叶片不同类型钙含量与海拔和土壤因子的关系,在RDA分析前需对参数进行标准化处理。在RDA排序图中,将每个环境因子箭头连线的长短视为环境因子对叶片各类型钙含量解释量的相对大小;环境因子与各类型钙含量的夹角角度(θ)能够反映二者的相关性,0°≤θ<90°,呈正相关;90°<θ≤180°,呈负相关;θ=90°,则无相关性[28]。
2 结果和分析
2.1 栲树叶片不同类型钙的含量及相关关系
2.1.1 叶片不同类型钙含量的比较 不同海拔样地间栲树叶片中不同类型钙含量的差异见表2。
由表2可以看出:随样地海拔的变化,栲树叶片中各类型钙含量呈现不同的变化规律。叶片中硝酸钙和氯化钙含量随样地海拔的升高总体呈现逐渐降低的趋势,以海拔500 m样地中最高,且显著(P<0.05)高于另4个样地;而另4个样地间的硝酸钙和氯化钙含量无显著差异。水溶性钙含量随样地海拔的升高总体呈现“降低—升高”的趋势,海拔800和900 m样地中均较高,且显著高于另3个样地;水溶性钙含量在海拔700 m样地中最低,且显著低于其他4个样地。果胶酸钙及草酸钙含量随样地海拔的升高总体呈现逐渐升高的趋势,均在海拔900 m样地中最高,且显著高于另4个样地;而另4个样地间的果胶酸钙及草酸钙含量无显著差异。磷酸钙和碳酸钙含量随样地海拔的升高总体呈现“升高—小幅降低”的趋势,在海拔700 m样地中最高、海拔500 m样地中最低,在海拔700~900 m样地间无显著差异,但均显著高于海拔500和600 m样地。硅酸钙含量随样地海拔的升高呈波动变化趋势,以海拔500 m样地中最高,且显著高于海拔600和900 m样地,而在海拔600~900 m样地间无显著差异。
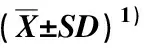
表2 不同海拔样地间栲树叶片不同类型钙含量的比较
在同一海拔的样地中,栲树叶片中不同类型钙含量的排序规律一致,从低到高依次为硝酸钙和氯化钙、水溶性钙、果胶酸钙、磷酸钙和碳酸钙、草酸钙、硅酸钙,均值依次为54.06、208.57、820.56、1 122.54、1 891.58、6 962.60 mg·kg-1。各样地的硝酸钙和氯化钙与水溶性钙的含量无显著差异,但这二者含量均显著低于其他4类钙含量;除海拔900 m样地外,其他样地的硅酸钙含量显著高于另5类钙含量;除海拔700和900 m的样地外,其他3个样地的果胶酸钙、磷酸钙和碳酸钙、草酸钙的含量无显著差异。
总体上看,在海拔500~900 m的5个样地间,叶片的总钙含量依次为12 350.43、8 653.85、11 752.14、11 382.13、11 160.97 mg·kg-1,其中,海拔700~900 m样地间总钙含量差异不大,而海拔500和600 m样地间总钙含量差异较大,与前者硅酸钙含量最高、后者草酸钙及硅酸钙含量均最低有关。
2.1.2 叶片不同类型钙含量间的相关性 栲树叶片中不同类型钙含量间的相关系数见表3。由表3可以看出:栲树叶片6个类型钙含量间存在不同程度的相关性。其中,硝酸钙和氯化钙含量与磷酸钙和碳酸钙含量呈极显著(P<0.01)负相关,与硅酸钙含量呈显著正相关;果胶酸钙含量与草酸钙含量呈极显著正相关;磷酸钙和碳酸钙含量与草酸钙含量呈极显著正相关。表明栲树叶片中的硝酸钙和氯化钙与磷酸钙和碳酸钙之间可能互相转化,且硝酸钙和氯化钙可与硅酸钙同步增减;栲树叶片中钙含量更倾向于果胶酸钙及磷酸钙和碳酸钙与草酸钙协同增减的分配策略。

表3 栲树叶片中不同类型钙含量间的相关系数1)
2.2 栲树叶片不同类型钙含量与海拔和土壤因子的关系
2.2.1 不同样地土壤理化指标的差异 不同海拔样地土壤理化指标的差异见表4。结果显示:供试样地的土壤温度随海拔升高逐渐降低,在海拔500 m样地最高、海拔900 m样地最低,总体上差异显著(P<0.05)。土壤含水量在海拔900 m样地最高,与另4个样地差异显著;而另4个样地间无显著差异。土壤pH值在海拔500 m样地最高,与另4个样地差异显著;而另4个样地间无显著差异。土壤全碳含量在海拔500 m样地最低,与另4个样地差异显著;而另4个样地的土壤全碳含量总体随海拔升高波动变化,但均无显著差异,其中海拔900 m样地的土壤全碳含量最高。土壤全氮含量随海拔升高逐渐提高,在海拔500 m样地最低、海拔900 m样地最高,总体上差异显著。供试样地土壤全磷含量均无显著差异,在海拔700 m样地最低、海拔600 m样地最高。土壤全钾含量随海拔升高波动变化,在海拔600 m样地最高且显著高于其他样地,在海拔500 m样地最低。土壤全钙含量在海拔500和600 m样地显著高于海拔700~900 m样地,在海拔700 m样地最低,并与其他样地差异显著。
表4 不同海拔样地土壤理化指标的比较
总体上看,在供试的5个样地中,在海拔相对较低(500和600 m)的样地内,土壤的温度、含水量和pH值以及全磷和全钙含量均较高,但土壤的全氮含量则较低。
2.2.2 栲树叶片不同类型钙含量与海拔和土壤理化指标的相关性 对栲树叶片中不同类型钙含量与样地海拔和土壤理化指标进行RDA约束排序分析,结果见表5和图1。
由表5可见:从单一因子看,海拔对栲树叶片中各类型钙含量的解释率最高,达28.8%,贡献率高达48.6%;8个土壤理化指标的总解释率达30.5%,总贡献率达51.4%,其中,土壤全钙含量的解释率和贡献率均最高(分别为9.0%和15.1%),表明海拔和土壤全钙含量对栲树叶片各类型钙含量的影响较大。

表5 海拔和土壤理化指标对栲树叶片中不同类型钙含量变化的解释率和贡献率1)
第1和第2排序轴的特征值分别为0.338和0.145,前2个RDA排序轴包括90.84%栲树叶片各类型钙含量指标以及海拔和土壤理化指标的信息。由图1可见:叶片硝酸钙和氯化钙含量与土壤的pH值、温度、全磷含量和全氮含量呈正相关,与土壤的全碳含量、全钙含量、全钾含量和含水量及海拔呈负相关;叶片水溶性钙含量与土壤的全钙含量、全钾含量、全碳含量和含水量及海拔呈正相关,与土壤的温度、pH值和全氮含量呈负相关,但与土壤全磷含量无相关关系;叶片果胶酸钙含量及草酸钙含量与土壤的全钾含量、全钙含量和含水量及海拔呈正相关,与土壤的全磷含量、全氮含量、温度和pH值呈负相关,与土壤全碳含量无相关关系;叶片磷酸钙和碳酸钙含量与土壤的全碳含量、全钙含量、全钾含量和含水量及海拔呈正相关,与土壤的全磷含量、全氮含量、pH值和温度呈负相关;叶片硅酸钙含量仅与土壤的全氮含量和pH值呈正相关,与其他因子呈负相关。

C1: 硝酸钙和氯化钙含量Content of calcium nitrate and calcium chloride; C2: 水溶性钙含量Content of water soluble calcium; C3: 果胶酸钙含量Content of calcium pectate; C4: 磷酸钙和碳酸钙含量Content of calcium phosphate and calcium carbonate; C5: 草酸钙含量Content of calcium oxalate; C6: 硅酸钙含量Content of calcium silicate.Alt: 海拔Altitude; pH: 土壤pH值pH value of soil; t: 土壤温度Temperature of soil; WC: 土壤含水量Water content in soil; TCa: 土壤全钙含量Content of total calcium in soil; TK: 土壤全钾含量Content of total potassium in soil; TN: 土壤全氮含量Content of total nitrogen in soil; TP: 土壤全磷含量Content of total phosphorus in soil; TC: 土壤全碳含量Content of total carbon in soil.
3 讨论和结论
3.1 栲树叶片不同类型钙含量差异的生态学意义及其对海拔变化的响应
植物体内钙含量约为0.1%~5.0%,钙离子浓度的改变可影响植物细胞中钙的存在形式,各类型钙相互调节并维持平衡,辅助植物适应环境异质性[29]。本研究中,栲树叶片含有的不同类型钙中,硝酸钙和氯化钙含量最低,且与磷酸钙和碳酸钙含量呈极显著负相关,这是因为硝酸钙和氯化钙为活性钙,在植物体内代谢较快并可迅速转化成其他类型的钙[4],而磷酸钙具有解毒作用并能提高植物抗性[30],因而,随着海拔上升、气温下降,细胞中的硝酸钙和氯化钙向磷酸钙转化,以提高栲树抵御寒冷环境的能力。
在海拔800和900 m样地中,栲树叶片的水溶性钙含量显著增加,其原因可能与这2个样地中的生境有关。许瑶瑶等[31]的调查结果显示:在郭岩山海拔800~900 m处,栲树林分的密度和郁闭度较小,光合能力较强。植物叶片所含的水溶性钙有利于植物体内钙离子的吸收利用[32],钙离子含量的增加可有效缓解低温依赖性光损伤及叶片中非结构性碳水化合物的过量积累,从而提高植物的光合作用并缓解低温冻害[33]。因此,在海拔800~900 m样地中,栲树叶片水溶性钙含量增加,可在提高栲树耐寒性的同时增强其光合作用,促进栲树个体的生长发育。
在寒冷环境下,植物体内水溶性果胶含量的增加,能有效提高植物抗寒性及微生物侵染能力[34]。本研究中,在海拔900 m样地中栲树叶片的果胶酸钙含量显著增加,其原因是果胶酸钙作为植物细胞壁的主要组成成分之一,对提高细胞壁的刚性和维持细胞膜的稳定性具有重要作用;且在海拔900 m处气温较低,栲树通过增加体内的果胶酸钙含量增强其对低温的抗性,确保植株正常生长。
在海拔700~900 m样地中,栲树叶片的磷酸钙和碳酸钙含量显著高于海拔500~600 m样地,这一现象可能与磷酸钙在植物体中的功能有关。磷酸钙具有较强的解毒作用,细胞中钙离子激活H+-ATP酶调节叶片细胞的pH值[17],并能在酸性环境下保持细胞中钙离子的稳定性[30],有利于增强植物抗逆能力[35]。因此,在供试样地的高海拔及酸性土壤环境下,栲树叶片通过磷酸钙的积累缓解酸性环境对植株的毒害作用并提高其抗性。
随着海拔上升、气温下降,栲树叶片通过增加对草酸钙的吸收,在细胞间隙沉积并形成各类晶体,以减轻有机酸积累过量对植物体产生的毒害[4,36],提高低温胁迫下植株的抗逆性并维持自身生长[37],因此,海拔900 m样地的栲树叶片中草酸钙含量显著增加。根据侯学煜[38]对植物生长与土壤钙质的依赖关系分类可知,栲树应为嫌钙或亚嫌钙植物;且栲树叶片中积累的钙主要以草酸钙和硅酸钙为主,这与曹建华等[4]对嫌钙植物叶片各类钙含量的研究结果类似。草酸钙和硅酸钙为非活性钙,其在植物体内难以被吸收、利用和转移[36],因而,在栲树叶片中草酸钙和硅酸钙的含量均高于其他4类钙。
本研究中,栲树叶片中果胶酸钙、磷酸钙和碳酸钙及草酸钙含量与海拔均呈正相关,这3类钙的含量两两之间也呈正相关,其原因可能为草酸钙的抗逆性[4]与磷酸钙的解毒性[30]具有协同作用,果胶酸钙能维持和增强组织的稳定性[34],这3类钙的含量随海拔升高协同增加,以应对气温变化对植物的胁迫作用。由此可知,栲树叶片各类钙含量随海拔变化而改变,这或许是栲树植株为适应因海拔变化而引起的气温变化所采用的调节策略之一,以实现对各类钙的再分配,使植株达到最适生长状态。在低温条件下,钙离子能够参与调控植物体内一些抗寒基因的表达,且钙含量较低的植物易受冻害[39,40]。因此,在实际营林过程中,当处于低温或较高海拔环境时,应适当考虑施用钙肥以调整栲树体内各类钙的分配策略,从而提高栲树植株对外部环境变化的适应性。
3.2 土壤理化性质对栲树叶片不同类型钙含量的影响效应
前述的研究结果表明:栲树不同海拔样地的土壤理化指标存在较大差异,不同土壤理化指标对叶片各类钙含量的解释率各异;由RDA排序结果可知,在8项土壤理化指标中,土壤的全钙含量、pH值和温度对栲树叶片中各类钙含量影响较大,其中,土壤全钙含量是影响栲树叶片各类钙含量变化的最重要的土壤因子(解释率为9.0%),且磷酸钙和碳酸钙含量与土壤全钙含量呈正相关。说明土壤钙是植物叶片钙的主要来源,土壤钙供应不足将直接导致植物生长受阻,提高土壤钙含量是增加植物抗性的有效措施之一[41]。土壤pH值也是影响栲树叶片中各类钙含量变化的重要因子之一,土壤的pH值与磷酸钙和碳酸钙含量呈负相关,这是因为在土壤酸性条件下,钙的吸收有利于维持细胞膜的正常结构和增强细胞的抗酸能力[17],故随着土壤pH值下降,栲树需要调整体内钙的分配策略,从土壤中吸收更多的钙并以磷酸钙的形式储存在叶片内,以此来提高植株的抗酸性。
研究结果表明:土壤温度与栲树叶片中水溶性钙、果胶酸钙及草酸钙的含量均呈负相关,产生此结果的原因可能与这3类钙的特性有关。随着土壤温度降低,栲树叶片可能将储存的钙更多地转化为水溶性钙、果胶酸钙及草酸钙,以共同抵御环境胁迫,但具体原因尚待深入研究。土壤全钾含量与叶片水溶性钙及果胶酸钙的含量呈正相关,这是由于土壤钾离子在一定程度上可增强植株对土壤钙的吸收,并提高叶片中钙的积累能力[42];而水溶性钙和果胶酸钙含量的增加则有利于钙离子的转移、吸收和利用。因此,土壤全钾含量提高则对栲树叶片钙的吸收有促进作用,可进一步提升栲树对环境的适应能力,这与刘进等[11]的研究结果类似。
在本研究中,土壤的全碳、全氮和全磷含量对栲树叶片中各类钙含量的影响效应较小,具体原因有待进一步研究。在中国南方地区植物易受冻害的原因主要是酸性土壤的钙供应水平较低[17]。因此,在中国南方进行栲树等树种的营林过程中,应考虑在传统施肥的基础上增施钙肥,不仅能有效提高土壤供钙能力,提供植物生长的必需元素,还能在一定程度上提高植株对各种逆境的抗性,维持其稳定生长。
3.3 结论
综上所述,在栲树叶片含有的6类钙中,除硝酸钙和氯化钙以及硅酸钙的含量与海拔呈负相关外,其余4类钙的含量均与海拔呈正相关,并通过协同增加草酸钙、磷酸钙和碳酸钙以及果胶酸钙含量的策略来应对海拔引起的生境变化。而在土壤理化指标中,土壤的全钙含量和pH值是影响栲树叶片中各类钙分配的重要因子,说明土壤钙是植物叶片钙的主要来源。在亚热带造林树种实际营林过程中,对种植在南方非钙质土壤或处于低温或高海拔环境下的林木树种,应考虑在传统施肥方式的基础上增施钙肥,以此改善土壤的酸化问题并降低林木遭受冻害的可能性,提高林木整体抗逆性,从而提高森林生产力。
猜你喜欢
军事文摘(2022年14期)2022-08-26
贵州畜牧兽医(2022年3期)2022-06-28
散文诗(2021年22期)2022-01-12
现代园艺(2021年23期)2021-12-01
西北大学学报(自然科学版)(2021年3期)2021-06-28
新农业(2020年18期)2021-01-07
现代园艺(2020年19期)2020-10-02
中国保健营养(2019年7期)2019-10-21
环球时报(2019-05-23)2019-05-23
现代养生·上半月(2015年11期)2015-12-08
