
Involvement of toll-like receptor 5 in mouse model of colonic hypersensitivity induced by neonatal maternal separation
2022-08-12 08:10GeoffroyMallaretAmandineLashermesMathieuMeleineLudivineBoudieuJulieBarbierYoussefAissouniAgatheGelotBenoitChassaingAndrewGewirtzDenisArdidFredericAntonioCarvalho
World Journal of Gastroenterology 2022年29期
Geoffroy Mallaret, Amandine Lashermes,Mathieu Meleine, Ludivine Boudieu, Julie Barbier,Youssef Aissouni,Agathe Gelot, Benoit Chassaing, Andrew T Gewirtz, Denis Ardid, Frederic Antonio Carvalho
Abstract
Key Words: Chronic abdominal pain; Colonic hypersensitivity; Toll-like receptors; Intestinal microbiota;Early life events
lNTRODUCTlON
Irritable bowel syndrome (IBS) is one of the major chronic gastrointestinal disorders, strongly related to stress. It is characterized by abdominal pain, changes in bowel habits and increased intestinal permeability without macroscopic organic alterations. Such changes has been hypothesized to trigger impairment of life’s quality and the development of comorbidities such as anxiety and depression[1]. A worldwide prevalence of 3%-5% has been reported and today, efficient pharmacological treatments are limited to relieve symptoms[2]. Colonic hypersensitivity (CHS), frequently associated with abdominal pain, has been described as the main cause of medical consultation in IBS patients with a prevalence ranging from 33% to 90%[3]. This symptom is defined by an altered sensation in response to colorectal stimuli and is clinically revealed by enhanced perception of mechanical triggers applied to the bowel.The common hypothesis is that CHS may result from colonic homeostasis changes and/or alterations of the brain-gut connection. In fact, the brain-gut axis has been shown to be impacted by inflammation and immunological factors, psychological factors, dysregulation of the hypothalamic-pituitary-adrenal(HPA) axis, abnormal activation of the vagus nerve and the enteric nervous system and intestinal dysbiosis[4].
Qualitative and/or quantitative alterations of the intestinal microbiota has been characterized in most of the functional gastrointestinal disorders including IBS[5]. Despite the numerous studies carried out,data on specific bacterial groups altered in IBS patients are still inconclusive. However,Enterobacteriaceaefamily,Lactobacillaceaefamily, andBacteroidesgenus seem to be increased in patients with IBS compared with controls, whereas unculturedClostridialesI,FaecalibacteriumandBifidobacteriumgenus were decreased in IBS patients[6]. Furthermore, it has been described that some IBS patients with chronic abdominal pain present specific intestinal microbiota dysbiosis, allowing considerations of the gut microbiota as a potential therapeutic target[7].
In healthy conditions, the interaction between gut microbiota and pattern recognition receptors(PRRs), especially local toll-like receptors (TLRs), allow maintenance of intestinal barrier in a homeostatic state. Indeed, TLRs, mostly present on the membrane of immune and epithelial cells,identify pathogen-associated molecular patterns (PAMPs) and induce intracellular signaling cascade resulting in the production of cytokines and chemokines important for colonic homeostasis. The mammalian TLRs family consists of 13 members (TLR1-10 in humans, TLR1-9 and TLR11-13 in mice)and each TLR responds to distinct PAMPs leading to the activation of specific signaling pathway. For example, TLR4 recognizes lipopolysaccharide (LPS) and TLR5, which is expressed in the basolateral membrane of the intestinal epithelium, detects flagellin (FliC)[8]. In a dysbiotic state, alterations in the signature of microbial molecules sensed by the host can lead to abnormal activation state of the immune system and induce a low-grade intestinal inflammation[9].
The breakdown of the symbiotic relationship between TLRs and gut microbiota could contribute to the development of various multifactorial intestinal diseases, such as IBS. Previous studies have reported modifications of TLRs expression and activation in intestinal biopsies of IBS patients[10-15].Furthermore, a preclinical study assessed the effect of neonatal maternal separation (NMS) in rats on TLRs expression, showing an upregulation of TLRs in colonic mucosa[16]. In this context, because of correlation between IBS and early life adverse events[17], this study investigated the impact of NMS paradigm on intestinal permeability, fecal microbiota composition and CHS development in mice as well as the association with TLRs expression.
MATERlALS AND METHODS
Animals
Seven-week-old wild type C57Bl/6J males and females mice were purchased from Janvier Laboratories(Le Genest Saint Isle, France). They were mated to obtain male pups for the NMS protocol. After birth,wild-type C57Bl/AJ pups were isolated from their mother from post-natal days P2 to P14, three hours per day (from 9:00 a.m. to 12:00 p.m.). These mice were named NMS mice. Pups were then left with their mothers up to weaning (P21) (Figure 1A). Control wild-type C57Bl/AJ pups were co-housed in the animal facility and were called non-handled (NH) mice. In addition, ten-week-old wild type C57Bl/6J males were purchased from Janvier Laboratories and used for FliC intrarectal instillation experiment.Animals were given access to food and waterad libitumand housed with a 12 h light-dark cycle. All experiments were performed on twelve-week-old male mice and were performed according to the ethical guidelines set out by the International Association for the Study of Pain (IASP), complied with the European Union regulation and were approved by ethics committees: The local committees C2EA-02 of Clermont-Ferrand (approvals CE110-12 and CE111-12).
Colorectal distension test
Colorectal distension test (CRD) was performed using the non-invasive manometric method described[18]. A miniaturized pressure transducer catheter (Mikro-Tip SPR-254; Millar Instruments, Houston, TX,United States) equipped with a custom-made balloon (length: 1.5 cm) prepared from a polyethylene plastic bag which avoid any colonic compliance effect. On the day of the experiment, the mice were accustomed to the holding device for 1 h before the CRD. Then, under mild anesthetic (2.5% isoflurane),the balloon was inserted into the rectum such that the distal end of the balloon was 5 mm from the anal margin. Subsequently, the animals were placed in the holding device and allowed to recover for 30 min prior to CRD. The balloon was connected to an electronic barostat (Distender Series II, G&J Electronics,Toronto, Canada) and a preamplifier (PCU-2000 Dual Channel Pressure Control Unit, Millar Instruments, Houston, TX, United States) connected to the PowerLab interface (AD Instruments,Dunedin, New Zealand). The barostat enabled the control of the balloon pressure. The distension protocol consisted of a set of increasing distension pressures (20, 40, 60, 80 and 100 mmHg), each of which was repeated twice, which was applied for 20 s with a 4 min inter-pressure interval. The signal was acquired and analyzed using LabChart 7 software (ADInstruments, Dunedin, New Zealand). After intracolonic pressure recording for each animals along the CRD protocols and signal treatment as previously described[18], intracolonic pressure variation (IPV), reflecting the colonic sensitivity, was calculated as previously described[19] for each distension pressure. Briefly, IPV was calculated by subtracting the integral (area under the curve) of the treated signal corresponding to the 20 s preceding the CRD from the integral (area under the curve) of the treated signal during the 20 s of CRD stimulation. Therefore, two groups of NMS mice were defined: NMS non-sensitized (NMS NS) and NMS sensitized (NMS S) mice. The NMS S animals are distinguished according to the area under the curve (AUC) value in response to the distention pressures from 60 to 100 mmHg during CRD procedure[20]. Briefly, if this value is higher than the average AUC of the NH control animals plus twice the SEM value (AUCNMSS≥ AUCNH+ 2 × SEMNH), this mouse is considered as hypersensitive and are placed in the NMS S group. Others are considered as NMS NS. For FliC intrarectal instillation experiment, the distension protocol was the same before intrarectal instillation and, only a set of distension pressure 60,80 and 100 mmHg was used 30 min, 60 min and 120 min after intrarectal instillation.
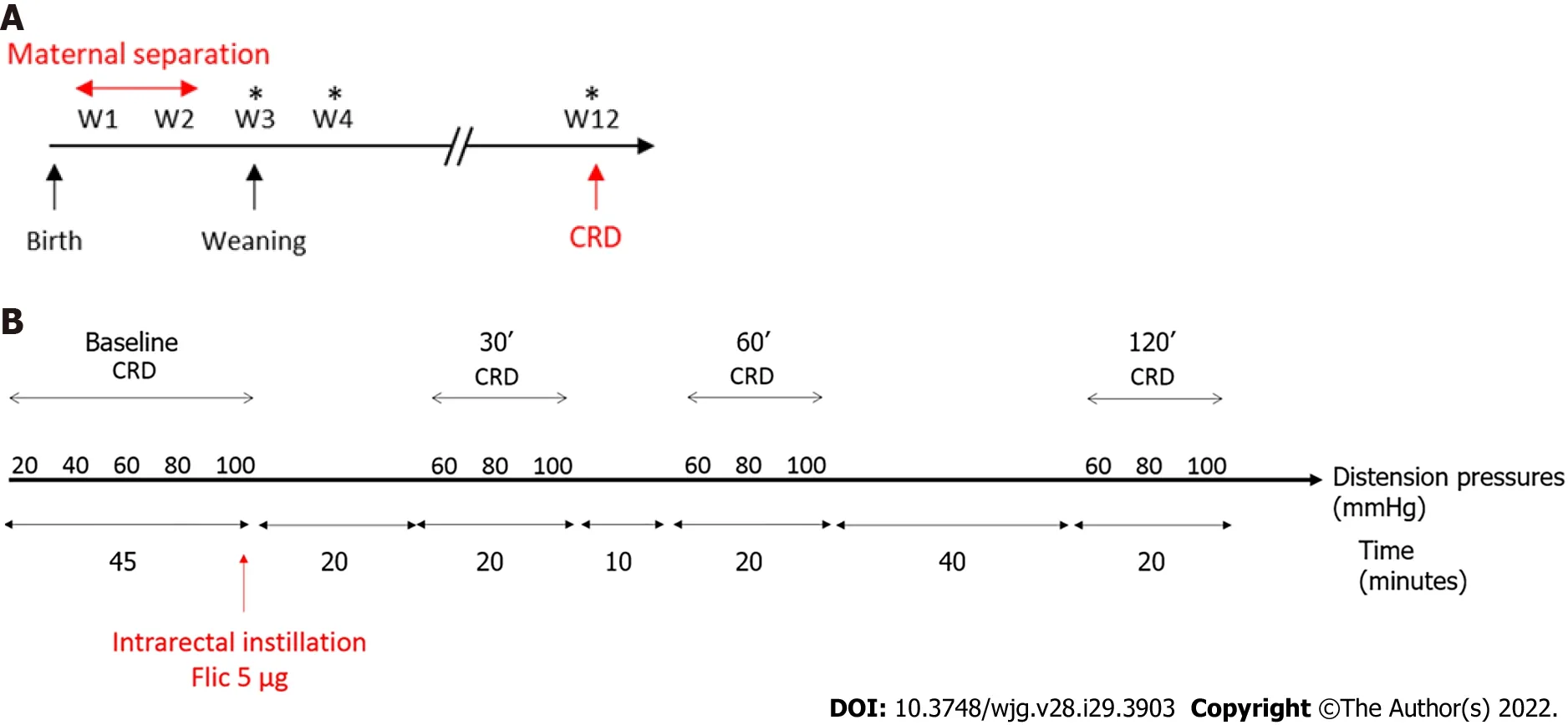
Figure 1 Time course protocols used in this study. A: Time course protocol for neonatal maternal separation experiment; B: Time course protocol for flagellin intrarectal instillation experiment. *Feces sample collection for Next Generation Sequencing; CRD: Colorectal distention test.
In vivo intestinal permeability
In vivointestinal permeability was assessed using fluorescein dextran (FITC- dextran 3000-5000 Da, TdB Consultancy AB, Uppsala, Sweden) as previously described[21]. Briefly, before CRD, NMS and NH mice were orally gavaged with 0.6 g/g body weight of FITC-dextran and blood samples were obtained from the retro-orbital venous plexus 3 h after this administration. Plasma FITC levels were determined by fluorometry at 488 nm using a microplate reader (Tecan, Lyon, France).
Fecal pellets collection, DNA extraction and microbiota sequencing
Fecal pellets were collected from mice at week 3, 4 and 12 and stored at -80 °C prior to DNA extraction.Bacterial DNA was extracted from fecal bacteria following the protocol of NucleoSpin®Soil kit(Macherey-Nagel, Düren, Germany). DNA concentrations and purity were then assessed using Take3 micro-volume plate and Epoch Microplate Spectrophotometer (BioTek, Winooski, VT, United States).The 16S rRNA gene V4 variable region polymerase chain reaction (PCR) primers 515/806 with barcode on the forward primer were used in a 30 cycles PCR using the HotStarTaq Plus Master Mix Kit(Qiagen®, Germantown, MD, United States). Next generation sequencing (NGS) was performed at Molecular Research DNA (MR DNA - Shallowater, TX, United States) on a MiSeq following the manufacturer’s guidelines. Sequences data analysis was performed using the quantitative insights into microbial ecology pipeline (QIIME)[22]. The analysis was carried out on the core microbiotai.e.the operational taxonomic units (OTUs) present in the fecal microbiota of 90% of the mice.
FliC intrarectal instillation
FliC from wildtypeSalmonella entericaserovar typhimurium (SL3201, fljB-) was provided by Pr. A.Gewirtz (Center for Inflammation, Georgia State University, Atlanta, GA, United States). Briefly, FliC was purified through sequential cation- and anion-exchange chromatography and purity was verified as described previously[8]. Intrarectal instillation was performed under mild anesthetic (2.5% isoflurane)using orogastric feeding tube and inserted 2.5 cm up the colon (Figure 1B). At this point, 50 μL of FliC diluted in PBS, corresponding to 5 μg was slowly administered over 30 s while pressure was applied to the anal area to prevent leakage. Following the injection of the solution, the tube was slowly removed and the rectal pressure was maintained for a further 30 s.
Colonocytes extraction
Following mice euthanasia, fragments of colon (3-4 cm) were flushed and opened longitudinally along the mesentery and homogenized in cold PBS to remove feces. Then, these fragments were incubated into HBSS containing EDTA solution (2 mmol/L) 30 min at 37 °C with strong agitation every 10 min.After HBSS/EDTA incubation, colons were removed and samples were centrifuged at 2000 g for 10 min.Then, HBSS/EDTA was removed and colonocytes were deep-frozen in liquid nitrogen and stored at -80°C for further analysis.
RNA extraction, reverse transcription and quantitative PCR
Total RNA from mice colonocytes was extracted using the RNeasy Plus Mini Kit (Qiagen®,Germantown, MD, United States) according to the manufacturer's protocol. After RNA extraction,reverse transcription was performed with the High Capacity cDNA RT Kit (Applied Biosystems, Foster City, CA, United States) with 500 ng of RNA, followed by a qPCR using LightCycler FastStart DNA Master SYBR Green Kit (Roche Applied Science, Penzberg, Germany). The primers used for TLRs expression analysis are described in Table 1. All results were normalized to theHPRTgene. Samples were tested in duplicate, and the average values were used for quantification by using 2-ΔΔCt method.
Fecal FliC and LPS load quantification
FliC and LPS were quantified using human embryonic kidney (HEK)-Blue-mTLR5 and HEK-BluemTLR4 cells, respectively (Invivogen, San Diego, California, United States). Fecal material was resuspended in PBS to a final concentration of 100 mg/mL and homogenized for 10 s using a Mini-Beadbeater-24 without the addition of beads to avoid bacteria disruption. The samples were then centrifuged at 8000 g for 2 min and the resulting supernatant was serially diluted and applied to mammalian cells. PurifiedEscherichia coliFliC and LPS (Sigma, St Louis, Missouri, United States) were used for standard curve determination using HEK-Blue-mTLR5 and HEK-Blue-mTLR4 cells,respectively. After 24 h of stimulation, cell culture supernatant was applied to QUANTI-Blue medium(Invivogen, San Diego, California, United States) and alkaline phosphatase activity was measured at 620 nm after 30 min.
Statistical analysis
Statistical analyses were performed with Prism 7 software (GraphPad, La Jolla, CA, United States). The Kolmogorov-Smirnov test has been used to check if data follow a normal distribution. One-way ANOVA, Kruskal-Wallis test or two-way ANOVA (more than two groups) were used for intergroupcomparisons with Tukey’s, Dunn’s and Dunnett’s test for the post-hoc analysis. Correlation was assessed using Pearson’s test. ANOSIM method followed by Monte-Carlo permutation test was performed to assess the significativity of beta-diversity analysis of fecal microbiota using the QIIME. APvalue ≤ 0.05 was considered statistically significant.
RESULTS
NMS paradigm induces CHS and intestinal permeability increase in a subset of mice
In order to evaluate colonic sensitivity, a CRD test was performed on twelve-week-old NH or NMS mice(Figure 1A). As previously described[23], among NMS mice only a subset developed CHS in comparison to NH mice Therefore, two groups of NMS mice were defined: NMS NS and NMS S mice.In fact, colorectal distension assessment revealed significant increase of IPV for the highest distension pressures 60, 80 and 100 mmHg in the NMS S group in comparison to NMS NS and NH groups(Figure 2A). Analysis of the areas under the curve (AUC) for each mouse confirmed this significant difference between NH, NMS NS and NMS S groups (Figure 2B). Intestinal permeability assessment revealed significant increase of FITC-Dextran plasma levels in the NMS S group compared to NH and NMS NS groups (Figure 2C).
Fecal microbiota dysbiosis is associated with CHS in neonatal maternal separated mice
Illumina sequencing of the 16S rRNA gene was performed on fecal pellets DNA extracts from NH, NMS NS and NMS S mice at W3, W4 and W12 (just before the CRD test) according to the time course protocol for NMS experiment (Figure 1A). Alpha-diversity analysis (number of observed OTUs) of the core fecal microbiota revealed no statistical difference between NH, NMS NS and NMS S animals at week 3,before weaning (Figure 3A-left panel). However, a significant decrease of species richness appeared at W4 in NMS S mice in comparison to NH or NMS NS animals and persisted at adulthood (W12, time point of CRD test), even if NMS NS and NMS S mice were co-housed in the same cage during all the experiment (Figure 3A-middle and right panels). In addition, a significant decrease of the observed OTUs number was present in NMS NS at adulthood (W12) in comparison to NH mice. Principal coordinates analysis based on unweighted UniFrac distances confirmed the alteration of the core fecal microbiota. It enabled to significantly (ANOSIM method followed by the Monte-Carlo permutation test,P< 0.05) identify the three animals’ groups from W3 to W12 (Figure 3B). The taxonomic analysis of the fecal core microbiota composition in the NMS S group revealed in twelve weeks old mice a decreased relative abundance of bacteria belonging to the phylumBacteroidetesand an increase inFirmicutesin comparison to the NMS NS group (Figure 3C). At lower taxonomic levels, NMS S mice were characterized by a decreased abundance of bacteria from the generaAllobaculumandBarnesiellacompared tocontrol NH mice, and a decreased abundance of bacteria from the generaBacteroidescompared to NMS NS mice. The relative abundances ofLachnoclostridium, ClostridiumandLactobacilluswere increased in these NMS animals with CHS in comparison to NMS mice without CHS. Surprisingly, the relative abundance ofLactobacilluswas decreased in NMS NS animals compared to NH group (Figure 3D).

Table 1 Primers used for toll-like receptors expression analysis

Figure 2 Neonatal maternal separation induces colonic hypersensitivity and increases intestinal permeability in mice. A: Intracolonic pressure variation (IPV) in response to colorectal distension in non-handled (NH), neonatal maternal separated non-sensitized (NMS NS) and neonatal maternal separated sensitized (NMS S) mice; B: Area under the curve (AUC) of the IPV relative to colorectal distension for each NH, NMS NS and NMS S mouse; C: FITCdextran 4 kDa plasmatic concentrations, 3 h after oral gavage with 15 mg of FITC-dextran of NH, NMS NS and NMS S mice. Values are expressed as a percentage of FITC-dextran per mL of plasma in comparison to the NH group mean. NH: n = 6; NMS NS: n = 6; NMS S: n = 8. aP < 0.05 and bP < 0.01 vs NH group; and dP <0.05, eP < 0.01 and fP < 0.001 vs NMS NS group. For IPV to CRD test, dots represent means and error bars represent SEM. For AUC and FITC-dextran, each dot represents one mouse and red lines represent means.
CHS induced by NMS exposure increased fecal level of FliC and is related to TLR5 overexpression in colonocytes
To understand potential mechanisms between fecal microbiota dysbiosis and CHS induced by NMS,quantification of two different PAMPs, FliC and LPS, was performed in feces from twelve-week-old NH, NM NS and NMS S mice. Exposure to NMS paradigm increased significantly fecal level of FliC(Figure 4A) rather than fecal LPS which is not significant better between different animal (Figure 4B).
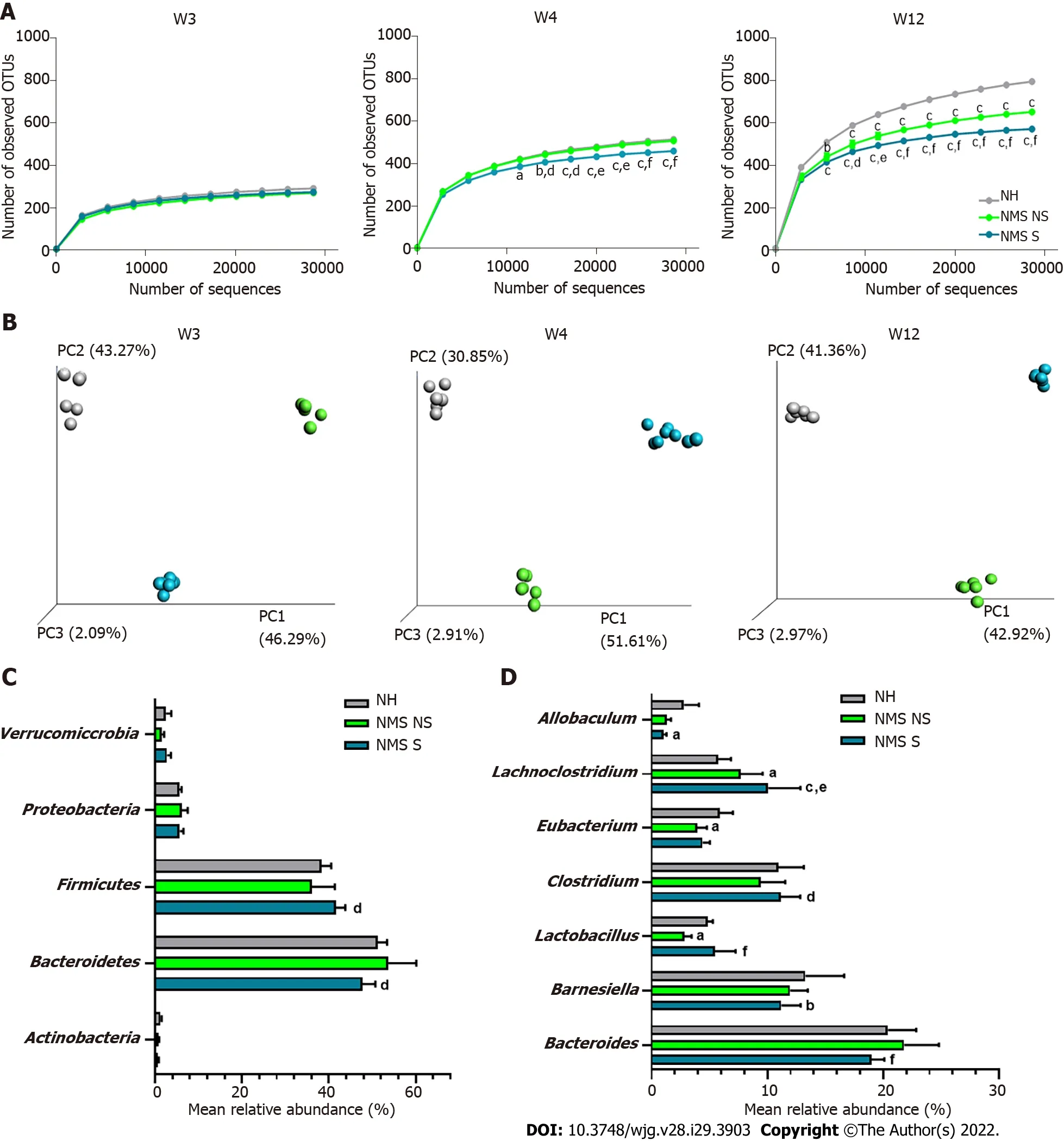
Figure 3 Neonatal maternal separation paradigm induces alterations of core fecal microbiota related to colonic hypersensitivity. A: Alphadiversity analysis of the core microbiota. Number of observed operational taxonomic units according to the number of sequences per samples of fecal samples from non-handled (NH), neonatal maternal separated non-sensitized (NMS NS) and neonatal maternal separated sensitized (NMS S) at week 3 (W3), week 4 (W4) and week 12 (W12); B: Beta-diversity analysis of the core microbiota. Principal coordinates analysis (PCoA) of unweighted UniFrac distances of NH, NMS NS and NMS S mice at W3, W4 and W12; C and D: Mean relative abundances of bacterial phyla (C) and genera (D) significantly altered by the NMS paradigm between NH, NMS NS and NMS S mice at W12. NH: n = 6; NMS NS: n = 6; NMS S: n = 8. aP < 0.05, bP < 0.01 and cP < 0.001 vs NH or dP < 0.05, eP < 0.01 and fP < 0.001 vs NMS NS groups respectively. For alpha-diversity analysis, dots represent means and error bars represent SEM. For PCoA analysis each dot represents one mouse.
As TLRs are the main receptors of PAMPs, the TLRs mRNA expression in colonocytes from NH,NMS NS and NMS S mice was quantified in adult (W12) mice. As previously described, three mouse groups were defined, based on the CHS (Supplementary Figure 1A and B). In those mouse groups, the TLR2, 3, 4 and 9 mRNA were not modified between NH, NMS NS and NMS S animals (Supplementary Figure 1C), whereas TLR5 mRNA expression is significantly increased only in NMS S subgroup(NH: 0.836 ± 0.200, NMS NS: 0.662 ± 0.120, NMS S: 1.925 ± 0.363,P< 0.05vsNMS NS) (Figure 4C). AUC corresponding to the IPV for highest colorectal distension pressures (60, 80 and 100 mmHg) significantly correlated with the mRNA expression level of TLR5 in colonocytes of NMS mice (P< 0.01) (Figure 4D).
FliC intrarectal instillation is associated with a transient increase of colonic sensitivity
Intrarectal instillation of FliC, agonist of the receptor TLR5, significantly increased IPV at the 60, 80 and 100 mmHg distension pressure 30 min and 60 min post-instillation (Figure 5A). The increase in the response to CRD test was transient and did not persist 120 min after FliC instillation. AUC confirmed this significant increase of IPV 30 min after intrarectal instillation of FliC (Figure 5B).
DlSCUSSlON
Abdominal pain, frequently associated with CHS, has been shown to be a common feature of IBS patients. It also strongly impacts on patient’s quality of life, leading to an important rate of consultation in Gastroenterology[24]. According to clinical studies, 33% to 90% of IBS patients exhibit CHS[3,25]. IBS presents a poorly first line treatment efficacy, especially regarding the treatment of abdominal pain[26].Thus, in accordance with the aim of our study, a better characterization of mechanisms associated with CHS is important for the establishment of new potential pharmacological targets.
The etiology of this condition, resulting in various symptoms, remains unclear even if biological,psychological and social factors seem to be involved. Indeed, several studies reported an increased risk of IBS associated with early adverse life events[17,27-29]. These events refer to traumatic experience during childhood such as physical, sexual or emotional abuse as well as discordant relationship with primary caretaker. Using NMS stress animal model[30], our study demonstrates the impact of early adverse life events on colonic sensitivity of adult mice. Interestingly, only a subset of NMS mice presented CHS, revealed by CRD test, compared to control non-handled mice. These results were consistent with data obtained in previous studies carried out in both rats and mice[23,31,32].
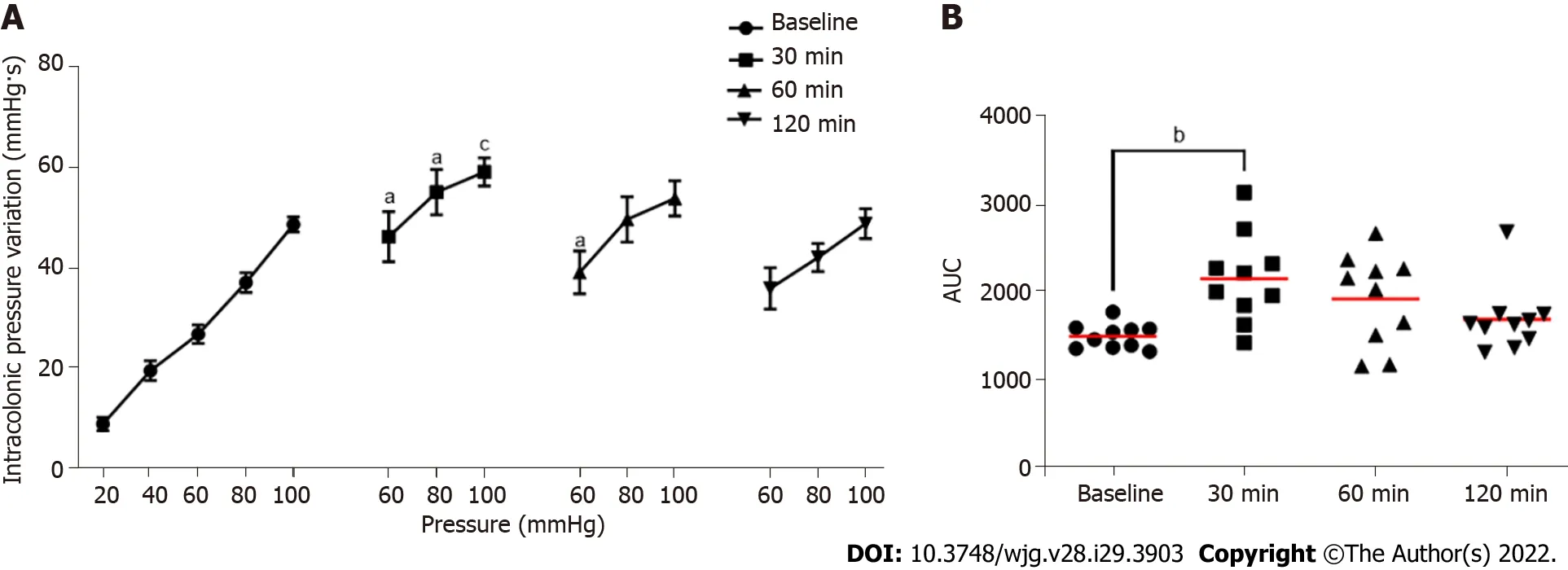
Figure 5 Evaluation of the impact of intrarectal instillation of flagellin on colonic sensitivity. A: Intracolonic pressure variation (IPV) in response to colorectal distension in males mice before (Baseline) and after (30, 60 and 120 min) intrarectal instillation of flagellin (5 μg); B: Area under the curve (AUC) of the IPV relative to highest colorectal distension pressures (60, 80 and 100 mmHg). For each mouse and each time point, n = 10. aP < 0.05, bP < 0.01 and cP < 0.001 respect to Baseline. For IPV to colorectal distension test, dots represent means and error bars represent SEM. For AUC, each dot represents one mouse and red lines represent means.
Many studies reported an association between activation of the HPA axis, the major neuroendocrine system regulating various bodily processes in response to psychological or physical stressors, and intestinal permeability increase[33,34]. Furthermore, alteration of the intestinal barrier is a key clinical feature of IBS and it has been related to CHS[35]. In our study, assessment of intestinal permeability was carried out by measurement of FITC-dextran plasma level. Only NMS animals with CHS exhibited high plasmatic levels of FITC-dextran, suggesting that NMS paradigm induced CHS is associated with altered intestinal barrier. This result is in accordance with previous reports showing increased intestinal permeability following NMS paradigm or chronic stress exposure[34,36,37]. In addition, the link between the weakness of the intestinal mucosa barrier and CHS has been demonstrated in a mouse model of post-infectious IBS[20].
Consistent studies reported intestinal dysbiosis in IBS patients[6]. The main distinguishing feature of IBS patients compared to heathy volunteers is on one hand the increased abundance of bacteria belonging to theFirmicutesphylum and on the other hand, the decreases abundance of bacteria belongs to theBacteroidetesphylum. Implication of the intestinal microbiota in CHS and associated chronic abdominal pain was also suggested[38]. In the present study, the characterization of the fecal microbiota composition using high-throughput sequencing of the 16S rRNA revealed the presence of a dysbiotic state making it possible to discriminate NH, NMS NS and NMS S mice. Indeed, the beta-diversity analyses showed that the composition of the fecal microbiota is different between NMS and NH control mice but also between NMS NS and NMS S mice while these animals came from the same litters and were co-housed. Changes in intestinal microbiota composition associated with NMS and CHS appeared very early, before weaning the animals (week 3) and persisted over time up to 12 wk. These alterations in the fecal microbiota composition were also characterized by a decreased bacterial richness in NMS S mice from week 4 to week 12. In general, this decrease was associated with a physiological disorder in the host, which seemed to be in agreement with the results obtained in this model[39]. A reduction in the bacterial diversity of the intestinal microbiota has notably been demonstrated in IBD and IBS patients but also in stress animal models[40-44].ClostridiumandLachnoclostridium, flagellated bacteria,are among the genera whose abundance was increased in NMS S mice, at W12, the time of colonic sensitivity assessment, compared to NMS mice without CHS. Studies carried out in animals subjected to stress during the neonatal period have also shown an increase in the relative abundance of theClostridiumgenus[43,45,46]. Furthermore and interestingly, Lunaet al[47] highlighted an increased relative abundance of different species ofClostridiumandLachnoclostridiumwithin the mucosaassociated microbiota in children with an autistic disorder associated with functional gastrointestinal disorders and in particular abdominal pain. These findings suggest an implication of the intestinal microbiota in the development of CHS in the NMS model.
In a dysbiotic state, particularly associated with an increase in intestinal permeability, alterations in the signature of microbial molecules sensed by the host can lead to a different activation state of the immune system[9]. Indeed, PAMPs, such as LPS or FliC, are sensed by PRRs including TLRs, which are expressed on the host cell surface or in the cytosolic compartment of numerous cell types. In this context, the aim of our study was to characterize the expression of different TLRs in colonocytes from our different animal subgroups after NMS paradigm. It is important to note that NMS paradigm is not associated with a modification of the intestinal inflammation status[23,36]. An increased TLR5 expression was observed only in animals presenting CHS after NMS paradigm, moreover, correlation between gene expression of TLR5 and AUC from 60 to 100 mmHg (corresponding to nociceptive stimulation) in NMS mice. These findings are in line with some reports showing upregulation of TLRs in IBS patient’s colonic biopsies[10-12,15]. An increased expression of some TLRs was also observed in NMS model but without association with visceral pain[11]. Few publications have indicated TLRs implication in animal pain model, especially inflammatory and neuropathic pain[48,49]. In visceral pain context, Tramullaset al[50] in 2014 demonstrated involvement of TLR4 in visceral sensitivity in a chronic stress model. Furthermore, Luczynskiet al[51] demonstrated increased colonic sensitivity to colorectal distention in germ free mice, associated with an increase of TLRs expression in spinal cord. Finally, in 2018, a study published by Zhouet al[52] established TLR4 implication in inflammatory visceral pain in animals with high-fat diet. Following the demonstration of FliC increase in NMS S mice fecal content and the upregulation of TLR5 expression in the NMS S mouse colonocytes, the effect of FliC was assessed on visceral sensitivity in naïve animals. We highlighted a transient increase of colonic sensitivity between 30 min and 60 min after FliC intra-rectal instillation. These results are the first to demonstrate potential FliC and TLR5 involvement in CHS in a non-inflammatory IBS-like animal model. Indeed, only Daset al[53] have shown that TLR5 signaling mediates hypersensitivity in a model of allodynia and that sensitivity was reversed by blocking TLR5 with a specific antagonist. Moreover,Dlugoszet al[54] has found a significantly higher serum level of antibodies to FliC patients with IBS.Our data, associated with the results of previous studies suggest that TLR5, through its activation by FliC, could play a key role in CHS induced by dysbiosis related to the NMS paradigm and more generally, in the pathophysiology of IBS.
CONCLUSlON
In conclusion, our results demonstrated the association of fecal dysbiosis, characterized especially by an increased abundance of flagellated bacteria, with impaired intestinal permeability, increased TLR5 expression and induced CHS. Taken together, TLR5 signaling upon recognition of FliC is relevant in visceral pain through both direct and indirect mechanisms, and application of TLR5-specific antagonists could potentially reversed CHS in non-inflammatory visceral pain context[23,36].
ARTlCLE HlGHLlGHTS


Research conclusions
Taken together, our results suggest a pathophysiological continuum between intestinal dysbiosis and CHS, with a role for TLR5.
Research perspectives
TLR5 signaling upon recognition of flagellin is relevant in visceral pain through both direct and indirect mechanisms, and application of TLR5-specific antagonists could potentially reversed CHS in noninflammatory visceral pain context.
ACKNOWLEDGEMENTS
The authors would like to acknowledge Abdelkrim Alloui (Animal facilities) for animal care.
FOOTNOTES
Author contributions:Mallaret G and Lashermes A contributed equally to this article; Study concept and design done by Mallaret G, Lashermes A, Ardid D and Carvalho FA; Acquisition of data done by Mallaret G, Lashermes A,Barbier J, Aissouni Y, Chassaing B; Analysis and interpretation of data done by Mallaret G, Lashermes A, Chassaing B, Gewirtz AT, Ardid D and Carvalho FA; Drafting of the manuscript done by Mallaret G, Lashermes A Meleine M,Boudieu L and Carvalho FA; Obtained funding done by Ardid D and Carvalho FA; Study supervision done by Gewirtz AT, Ardid D and Carvalho FA.
Supported bythe Region Auvergne-Rhône-Alpes and FEDER, No. Thématiques émergentes and Pack Ambition Recherche; the French Government IDEX-ISITE Initiative, No. 16-IDEX-0001-CAP 20-25; the Ministère de la Recherche et de la Technologie, INSERM and University of Clermont Auvergne, No. UMR1071.
lnstitutional animal care and use committee statement:All experiments were performed according to the ethical guidelines set out by the International Association for the Study of Pain, complied with the European Union regulation and were approved by ethics committees: the local committees C2EA-02 of Clermont-Ferrand (approvals CE110-12 and CE111-12).
Conflict-of-interest statement:All authors have nothing to disclose.
Data sharing statement:All sequencing raw data have been deposited in European Nucleotide Archive (ENA) under accession number PRJEB50651.
ARRlVE guidelines statement:The authors have read the ARRIVE guidelines, and the manuscript was prepared and revised according to the ARRIVE guidelines.
Open-Access:This article is an open-access article that was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution NonCommercial (CC BYNC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is noncommercial. See: https://creativecommons.org/Licenses/by-nc/4.0/
Country/Territory of origin:France
ORClD number:Geoffroy Mallaret 0000-0002-3658-3418; Amandine Lashermes 0000-0002-8241-464X; Mathieu Meleine 0000-0002-0925-087X; Ludivine Boudieu 0000-0002-4774-9313; Julie Barbier 0000-0001-7177-3236; Youssef Aissouni 0000-0002-9264-7690; Agathe Gelot 0000-0002-7884-6968; Benoit Chassaing 0000-0002-4285-769X; Andrew T Gewirtz 0000-0002-6338-7578; Denis Ardid 0000-0003-4320-7338; Frederic Antonio Carvalho 0000-0003-1592-2369.
S-Editor:Zhang H
L-Editor:A
P-Editor:Yu HG
 World Journal of Gastroenterology2022年29期
World Journal of Gastroenterology2022年29期
- World Journal of Gastroenterology的其它文章
- Mechanistic and functional extrapolation of SET and MYND domain-containing protein 2 to pancreatic cancer
- Clinical challenge for gastroenterologists-Gastrointestinal manifestations of systemic mastocytosis: A comprehensive review
- Structural changes of proteins in liver cirrhosis and consequential changes in their function
- Epidemiologic and socioeconomic factors impacting hepatitis B virus and related hepatocellular carcinoma
- Enhanced endoscopic ultrasound imaging for pancreatic lesions: The road to artificial intelligence
- Qingyi decoction attenuates intestinal epithelial cell injury via the calcineurin/nuclear factor of activated T-cells pathway