
酸模对Cd、Cu、Pb 复合污染农田的修复潜力
2022-08-11 14:02赵怡阳陶祥运张易旻王燕蒋位青
浙江农业科学 2022年8期
赵怡阳 ,陶祥运 ,张易旻∗ ,王燕 ,蒋位青
(1.浙江博世华环保科技有限公司,浙江 杭州 310015;2.诸暨奕禾环保科技有限公司,浙江 诸暨 311825)
酸模 (Rumexacetosa) 是蓼科 (Polygonaceae)酸模属中一种常见的多年生草本植物,广泛分布于我国各地山坡、路边、荒地或沟谷溪边,是长江中下游铜矿区的优势植物。酸模根系可深达1.5~2.0 m;在冬季结冰条件下仍可保持较强的生命力。Tang 等[1]报道生长在长江流域铜矿区的酸模对Cu 的平均积累量达到601 mg·kg-1。
关于酸模属植物在污染修复中的应用研究屡见不鲜,例如杂交酸模对盐渍化土壤的开发和改良[2],矿区废弃地酸模对重金属的吸收[3]及其在Pb、Zn 胁迫下的耐性机理[4],以及酸模对Cu 的积累特性研究[5]等。Barrutia 等[3]研究施肥对矿区和非矿区酸模生长特性的影响,结果表明,施肥对矿区酸模的生长有很好的促进作用,体现了酸模对重金属的耐性。李红艳等[5]研究酸模对铜的耐性和积累特性,结果表明,酸模地上部对Cu 的积累量可达到1 500 mg·kg-1以上,可用于Cu 污染土壤的修复。Babalonas 等[6]研究表明,小酸模是生长在Cu、Zn 等重金属污染土壤上最重要的物种之一。Hulina 等[7]对克罗地亚Mahovo 地区的几种野草中重金属含量进行了调查,结果表明,皱叶酸模(Rumexcrispus) 对Cu、Zn 具有很强的吸收能力。王小平等[8]研究了酸模在碱性土壤上修复Cd、Pb污染的土壤,结果表明,酸模对镉的修复系数和转运系数均较为优秀。
然而,酸模对多种重金属复合污染土壤修复的相关研究鲜见报道,本文将采用田间试验的研究方法探究酸模在Cd、Cu、Pb 复合污染的情况下对重金属的富集及植物的生长情况,为今后酸模用于大面积农田土壤修复提供了基础。
1 材料与方法
1.1 材料
供试植物:酸模,种子采集于进贤当地田间。供试土壤:采用田间原位试验,在江西省南昌市进贤县某农田选择不同重金属污染程度地块A、地块B,地块土壤理化性状见表1。

表1 供试土壤的理化性质
实验用主要仪器包括:原子吸收光谱仪(ZEEnit700P,德国耶拿分析仪器有限公司)、研磨仪 (ZM200,赛默飞世尔科技公司)、恒温水浴锅 (HH-8,国华电器有限公司)、超声波清洗器(KQ5200,昆山市超声波仪器有限公司)、微波消解仪 (MARS XPRESS,北京安南科技有限公司)电子天平 (FA2104N,上海民侨精密科学仪器有限公司)、电热恒温鼓风干燥箱 (DHG-9243BS-Ⅲ,上海新苗医疗器械制造有限公司)、高速冷冻离心机 (ST16R,赛默飞世尔科技公司) 和超低温冰箱(赛默飞世尔科技公司)。
1.2 方法
1.2.1 酸模种植处理设计
育苗。酸模种子用1%NaClO 溶液灭菌10 min,先用自来水冲洗数次,再用去离子水冲洗3 次,然后放入初始温度为45 ℃的蒸馏水中浸泡,24 h 后取出放入培养皿中,加蒸馏水浸泡催芽。将露白的种子放入盛有供试土壤的穴盘中,待幼苗长出3~4片真叶,选择均匀一致的幼苗移栽。
种植。选择长势均匀的酸模幼苗分别移栽到两块不同程度重金属污染的土壤中。每个地块按照36 株·m-2种植;每个地块设20 穴,每穴种植2株,每一生育期设4 个重复,即每块地需种植80株苗。种植前向土壤中施入底肥,生长到第2 个月时再次施肥,每次施用化肥用量约为尿素0.4 kg,磷酸二铵0.3 kg,硫酸钾0.4 kg,N∶P∶K 比为2∶1∶1.2。
1.2.2 样品的采集和处理
酸模分别于移栽后30、45、60、75 和90 d 各取样一次,同时采集各时期的土壤样品进行指标测定。将植物样分成茎叶和根两部分,剪碎放入烘箱,在105 ℃下烘0.5 h,再在75 ℃下烘至恒重,将植物干样品粉碎,过2 mm 筛待测。称取0.500 0(±0.000 5) g 植物样品,放入150 mL 锥形瓶中。设3 个平行,加空白。用HNO3∶HClO4体积比4∶1 的混酸加盖消解至溶液呈透明或浅黄色。用0.2% HNO3定容至25 mL 容量瓶,用原子吸收光谱仪测定样品中重金属含量。土壤中重金属含量的测定采用欧共体物质标准局提出的BCR 逐级提取法。
1.2.3 指标计算公式
富集系数=植物各组织中重金属含量/土壤中重金属含量;
转移系数=植物地上部分重金属含量/植物地下部分重金属含量。
1.2.4 数据处理
所有处理均设3 个平行,实验所得数据使用计算机软件Microsoft Excel 2007 和Origin 8.0 进行数据处理,并用SPSS 17.0 进行统计分析和显著性检验。
2 结果与分析
2.1 不同生育期酸模生物量的变化
A、B 中酸模生物量分别为2.11~15.61 g 和1.88~13.57 g;在75 d 时,地块A、B 中酸膜生物量分别从15.61 g 和13.57 g 跃增至115.65 g 和99.87 g,至90 d 时,达到生物量最大值136.61 g和117.23 g。从整个生育期比较A、B 地块中酸膜的生物量,B 地块中酸模的生物量明显低于A 地块;两者的茎叶部从最初相差0.22 g 到90 d 相差15.67 g,根部从最初相差0.01 g 到90 d 相差3.71 g;在这期间,两者生物量之间的差距逐渐加大。因此,酸模生长所受到的抑制作用与重金属污染程度呈正相关。采用F 检验法对A、B 地块中植物不同生长期的生物量进行显著性差异分析,置信度按95%计 (自由度=19,F表取2.18),结果 (表2)表明,A、B 地块中植物根的生物量在30、60 和75 d 时存在显著性差异;植物茎在45、60 d 时存在显著性差异;在90 d 时,两个地块植株生物量不存在显著性差异,说明本研究中的土壤重金属含量差距不足以严重影响酸模的生长。

表2 不同重金属复合污染水平下酸模不同生长时期的生物量
2.2 不同生育期酸模茎叶部和根部对Cd、Cu、Pb 的积累和分布
从图1 可看出,在整个生育期内,A 地块中酸模茎叶部对Cu、Pb 的富集量均表现出先升后降再升的趋势,在生长的45 d 分别达到最高富集量13.31 mg·kg-1和9.68 mg·kg-1;在75 d 之后,酸模对Cu、Pb 的富集量迅速降低,分别比45 d 下降了69.9%和73.6%。酸模根部对Cu、Pb 的富集量要大于茎叶部,在45 d 分别达到最高富集量30.11 mg·kg-1和35.72 mg·kg-1;在75 d,酸模对Cu 和Pb 的富集量又迅速降低。在B 地块中,酸模茎叶部和根部对Cu、Pb 的富集量在整个生育期内表现出与A 地块相同的富集趋势。B 地块中酸模茎叶部在5 个生育期对Cu 的富集量分别是A 地块的1.69 倍、1.70 倍、1.15 倍、2.33 倍、1.26倍,根部是其1.10 倍、1.82 倍、0.91 倍、1.60倍、1.45 倍;茎叶部对Pb 的富集量分别是其2.45倍、1.43 倍、1.52 倍、0.55 倍、1.61 倍,根部是其2.27 倍、1.77 倍、1.15 倍、0.49 倍、1.18 倍。因此,除75 d 酸模的根部外,随着土壤中Cu、Pb污染程度增加,酸模茎叶部和根部对Cu、Pb 的富集量都有显著增加,并且在整个生育期表现出相同的积累态势。
从图1 可见,虽然在整个生育期内,酸模茎叶部对Cd 的富集量表现出与对Cu 富集相同的富集趋势,但不同的是,酸模对Cd 的富集在60 d 才达到对Cd 最大富集量;酸模根部对Cd 的吸收与酸模茎叶部对Cd 的吸收在整个生育期内具有相同的富集趋势。在75 d,酸模对Cd 的富集量显著降低,但在90 d,酸模茎叶部和根部对Cd 的富集量有所回升;当土壤中Cd 的含量从0.70 mg·kg-1增加到1.34 mg·kg-1时,酸模茎叶部与根部对Cd的富集量都有所增加,并且根部对Cd 的富集量要高于茎叶部。由此可见,酸模在不同生育期对不同重金属的吸收具有偏好性。

图1 酸模不同部位在各个生长期对Cd、Cu、Pb 的富集量
综上分析可知,酸模对Cd、Cu、Pb 富集量随时间的变化分为3 个阶段,0~45 d 是酸模吸收Cd、Cu、Pb 的敏感期。此时酸模根系快速生长,对土壤中的重金属有一个快速积累的过程,富集量显著升高。45~75 d,酸模对土壤中Cd、Cu、Pb的富集量降低,此时期酸模茎叶部和根部在生长代谢旺盛,使得重金属通过土壤和根不断向茎叶部转移;随着根部化学性状的变化和茎叶部代谢机制减弱,茎叶部和根部吸收的重金属量逐渐减少[9-10]。此时酸模的生物量急剧增大,说明当酸模进入旺盛生长后,茎叶部快速生长,酸模叶片迅速增大,前期吸收的Cd、Cu、Pb 在酸模体内得到稀释,符合作物生长的稀释效应[11]。75~90 d,酸模中Cd、Cu、Pb 含量再次增加,原因是随着酸模从营养生长到生殖生长的转变,根系分泌物的种类发生变化,更有利于活化被土壤吸附的Cd、Cu、Pb,从而促进酸模对Cd、Cu、Pb 的吸收;其次,酸模根系发育第二个高峰期的到来,根系活力增加和吸收区域扩大,能够吸收到更多的重金属。尽管在整个生育期内酸模对土壤中Cd、Cu、Pb 的富集量随着复合污染程度的增加而上升,但总的来说,重金属主要富集于酸模的根部,特别在酸模生长的45~60 d,其根部重金属的含量远远高于茎叶部。
2.3 不同生育期酸模对农田中Cd、Cu、Pb 的富集系数和转移系数
2.3.1 富集系数
从表3 可知,在Cd、Cu、Pb 复合污染条件下,酸模不同部位对各重金属的富集能力各不相同,在生长的45~60 d,酸模对各重金属的富集系数达到最大值。随着土壤重金属污染加剧,酸模对Cu 和Pb 的富集能力改变较少,但对Cd 的富集能力有较大程度的下降。除此之外,酸模对重金属Cu 和Pb 的富集能力较差,富集系数都很低,而对Cd 的富集系数超过了1,最高可达5.77,由此可见,酸模在30~90 d 内对各重金属富集能力表现为Cd>Cu>Pb。
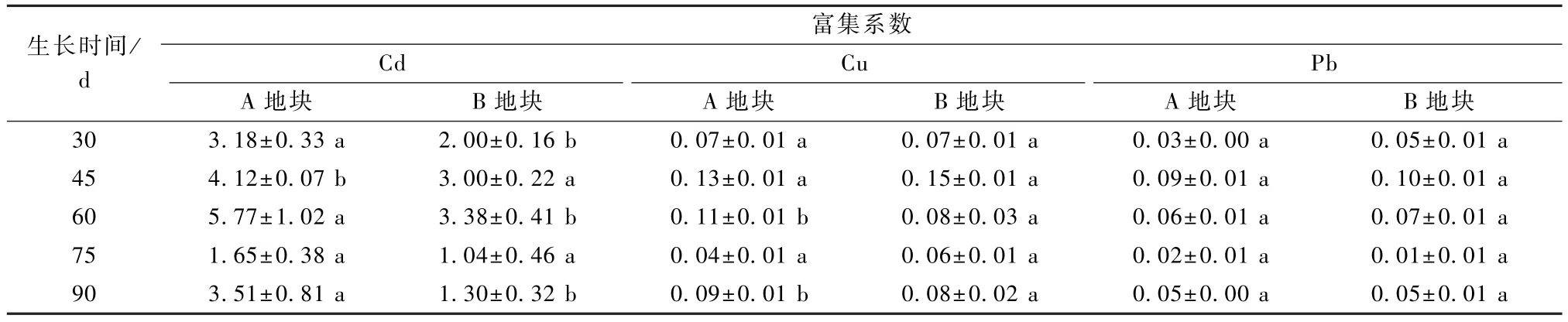
表3 酸模对Cd、Cu、Pb 富集系数
2.3.2 转运系数
从表4 可知,酸模在不同生育期,对不同重金属表现出不同的转移能力。A 地块内,除30 d和75 d 酸模对各重金属的转移能力规律为Cd>Pb>Cu 之外;其余3 个时期酸模对各重金属的转移能力表现的各不相同。B 地块内,酸模生长的45 d 和75 d,对各重金属的转移能力相同,为Cd>Cu>Pb;酸模生长的第60 天和第90 天,转移能力亦相同,为Cu>Pb>Cd。从整个生育期看,酸模对Cd、Cu、Pb 的转移系数最大值都出现在75~90 d,并且酸模对Cu、Cd 的转移能力总体表现出持续上升的趋势,只有在复合污染浓度较高时,酸模生长的90 d 对Cu、Cd 的转移系数有所下降。

表4 酸模对Cd、Cu、Pb 转移系数
2.3.3 综合分析
从表3 和表4 可以看出,酸模对土壤中Cu、Pb 的富集及转运效果不佳,这与之前报道的酸模对Cu 和Pb 具有超量的富集能力不符[3,5,12],并且不符合Cu、Pb 超积累植物的要求,但酸模对于Cd的富集能力大大超过了Cu 和Pb,其茎叶部和根部对Cd 的富集系数均超过了1。相比其他富集植物的研究结果,如龙玉梅等[13]研究表明,龙葵在模拟镉污染的不同种类土壤中转运系数为0.99~2.53,富集系数为2.13~10.68;魏树和[14]研究龙葵发现,对Cd 的富集系数为2.68,可推断酸模在实际Cd、Cu、Pb 复合污染农田中对Cd 仍具有富集的潜力。
3 讨论
本研究采用农田试验对酸模进行培养的方式,研究酸模的富集能力和转移能力,结果表明,在Cd、Cu、Pb 复合污 染农田 中,酸模对Cd、Cu、Pb 的富集系数最高分别达到5.77、0.15 和0.10,转移系数最高分别为1.19、0.89、0.68,富集能力为Cd>Cu>Pb。总体来说酸模在Cd、Cu、Pb 复合污染农田中对Cd 有足够的修复潜力,但是针对Cu 和Pb 修复存在较大的短板,在后续的研究中可尝试其他富集植物与酸模间作、套作等方式开展修复,以解决含Cd 重金属复合污染农田修复的问题。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
农业工程学报(2022年6期)2022-06-27
中国烟草科学(2022年2期)2022-05-27
福建林业科技(2020年4期)2020-12-31
今日农业(2020年19期)2020-12-14
西藏文学(2020年5期)2020-09-21
安徽农学通报(2020年7期)2020-05-26
快乐语文(2019年35期)2020-01-06
中国诗歌(2019年6期)2019-11-15
湖北农业科学(2018年18期)2018-12-11
