
基于转录组的糙苏MYB 转录因子的挖掘及分析
2022-08-11 14:02熊勇赵春艳李晓艳
浙江农业科学 2022年8期
熊勇 ,赵春艳 ,李晓艳
(1.云南民族大学 民族医药学院,云南 昆明 650500;2.中华全国供销合作总社昆明食用菌研究所,云南 昆明 650221)
糙苏 (PhlomisumbrosaTurcz.) 是唇形科(Labiatae) 糙苏属 (Phlomis) 的多年生直立草本植物,糙苏的根和全草均可入药,对肠胃炎、肺炎、咳嗽感冒具有疗效,也可以用于治辽牛马肺痈等[1]。糙苏作为一种民间常用草药,在其临床及中成药中的应用也在不断扩大,糙苏在化学成分、药理作用及生药学研究方面不断深入,糙苏中含有苯乙醇苷类、环烯醚萜类 (根)、黄酮类 (茎和叶)、萜类 (根和种子)、木质素 (地上部分) 及挥发油 (叶、花和籽) 等化学成分[2],但在其分子遗传及基因组方面的研究报道不多。
转录因子 (transcription factors,TFs) 通过与目的基因的远端和局部顺式元件结合来调控基因表达,是在植物发育、细胞周期、细胞信号和逆境反应中发挥着重要作用的调节因子。典型的转录因子包括转录调控区、核定位信号区、寡聚反应区和DNA 结合区,这些功能区域决定了转录因子的结构和特征[3]。DNA 结合区的氨基酸序列高度保守,这些保守的氨基酸决定了转录因子与顺式作用元件识别及结合的特异性。MYB 转录因子是植物中最大的转录因子家族之一,在植物生长发育、次生代谢调控、胁迫应答等生命活动中起着调控作用。MYB 转录因子是指以含有MYB 结构域为共同特征的一类转录因子,每个MYB 结构域都包含50~53 个氨基酸残基高度保守的重复序列[4]。根据MYB 转录因子N 端保守的DNA 结构域的重复个数,把植物MYB 转录因子家族分为4 个亚类,1R-MYB (related-MYB)、R2R3-MYB、3R-MYB 和4R-MYB[5]。1R-MYB 亚类成员只含有一个MYB 结构域,可能是一类重要的端粒结合蛋白,在维持染色体结构的完整性和调节基因转录上起着重要作用。R2R3-MYB 亚类成员N 端含有两个MYB 结构域,是成员最多的一个亚类,主要参与初级或次生代谢的调节,发育过程及对生物或非生物胁迫的响应[6-8]。3R-MYB 亚类成员的第一个保守色氨酸残基 (W) 会被苯丙氨酸 (F)、异亮氨酸(I)或亮氨酸(L)所取代[9]。4R-MYB 亚类成员的研究较少,只在少数的植物中进行表达[10]。
本研究以高通量测序技术构建转录组数据库,利用生物信息学方法对糙苏MYB 转录因子的理化性质、氨基酸序列及保守基序、高级结构、系统进化树进行系统分析,并通过GO 注释、蛋白质功能预测和表达模式进行深入分析,为进一步探究糙苏MYB 转录因子家族基因的潜在功能研究奠定基础。
1 材料与方法
1.1 MYB 序列鉴定与理化性质分析
基于前期构建的糙苏转录组测序数据库,获得糙苏MYB 转录因子基因家族氨基酸序列,通过NCBI 的ORF finder 预测各个MYB 基因开放阅读框,获得完整的编码蛋白序列,应用HMMER、Smart 和NCBI BLAST 与相应的蛋白数据进行蛋白相似性比对分析鉴定。
利用ExPASy-ProtParam 对糙苏MYB 蛋白进行理化性质分析[11]。利用ProtComp 和SignalP 在线软件对糙苏MYB 蛋白进行亚细胞定位和信号肽进行预测。
1.2 MYB 序列保守结构域及系统发育树分析
利用DNAMAN 8.0 对糙苏MYB 转录因子进行氨基酸序列对比;利用Motif 分析工具MEME 数据库对糙苏MYB 进行保守基序分析。利用二级结构分析软件SOPMA 对糙苏MYB 转录因子编码蛋白进行二级结构预测,并利用SWISS-MODEL 对不同亚类糙苏MYB 转录因子进行三级结构建模。
从PlantTFDB 数据库中下载拟南芥MYB 转录因子序列作为参考序列,利用MEGA 6.06 软件内的Clustal W 程序,与糙苏MYB 转录因子氨基酸序列进行对比分析,采用邻接法 (Neighbor-joining method) 构建系统发育树,参数设置P-distance、Bootstrap method 重复1 000 次,其他参数为默认值。
1.3 MYB 序列的功能分析和表达分析
通过糙苏的高通量测序数据获得基因的GO 注释号,利用 QuickGO (https://www.ebi.ac.uk/QuickGO/) 进行GO 注释信息查询,分析糙苏MYB 转录因子蛋白GO 富集的功能注释。以拟南芥作为参考,利用 STRING (https://string-db.org/) 的Muitipie sequence 对糙苏MYB 转录因子蛋白进行蛋白质功能预测。依据MYB 基因家族根和叶的fpkm 值,通过Tbtools 软件内置HeatMap 程序对糙苏MYB 转录因子进行聚类分析并绘制热图。
2 结果与分析
2.1 MYB 序列鉴定与理化性质
从转录组数据库中获得68 条糙苏MYB 碱基序列,通过HMMER、SMART 和NCBI BLAST 结构预测,去除重复序列及冗余转录本后,最终得到61 条具有MYB 保守结构域的序列,命名为PuMYB1~61,其中含11 个1R-MYB (related-MYB) 类转录因子,49 个 R2R3-MYB 类转录因子,1 个 4R-MYB(PuMYB38) 类转录因子。
ProtParam 对糙苏MYB 转录因子家族一级结构的理化性质分析结果 (表1) 显示,61 个糙苏蛋白的氨基酸数目在98~1 756,平均分子量为38 608.20 u;61 个糙苏蛋白中有25 个糙苏蛋白的理论等电点大于7 (偏碱性),而有36 个小于7 (偏酸性),最小值为4.82 (PuMYB54),最大值为9.92 (PuMYB11),平均值为6.97;从不稳定系数来看,仅有2 个糙苏蛋白 (PuMYB38、PuMYB51)的不稳定系数小于40 (稳定蛋白),而其他59 个均大于40 (不稳定蛋白),说明糙苏MYB 转录因子家族整体是不稳定蛋白;糙苏MYB 转录因子家族均为亲水性蛋白,且热稳定性较高,因为MYB家族蛋白的脂肪系数和总平均亲水性GRAVY 均小于0。
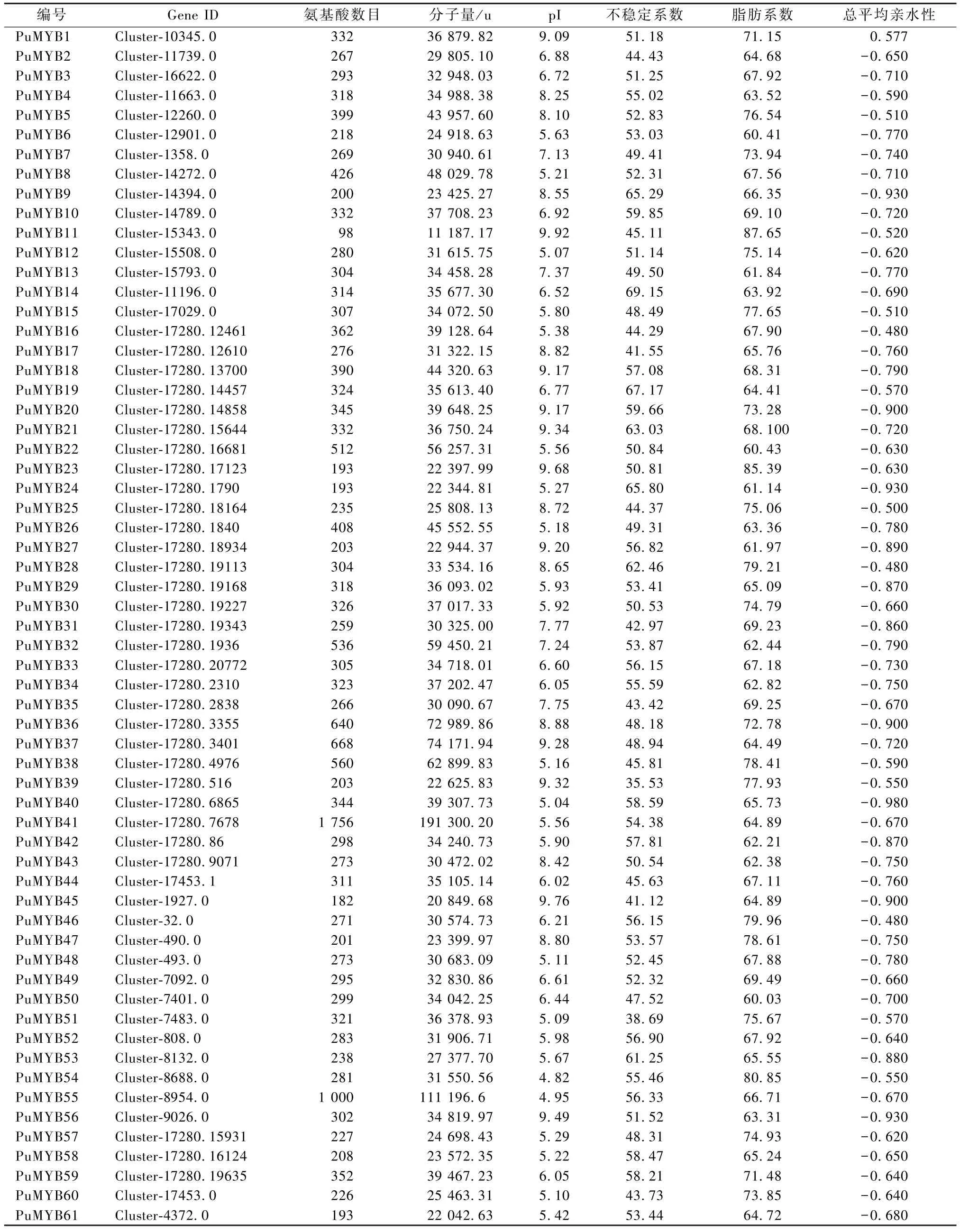
表1 糙苏MYB 转录因子家族蛋白理化性质
ProtComp 分析显示,61 个糙苏MYB 蛋白质中共有50 个定位在细胞核中,其余11 个MYB 成员定位在细胞核外。采用SignalP 对信号肽进行预测,61 个糙苏MYB 蛋白质中,有5 个存在信号肽(PuMYB57、PuMYB58、PuMYB59、PuMYB60、PuMYB61),为分泌蛋白。
2.2 MYB 基因家族的结构
2.2.1 MYB 结构域
DNAMAN 对糙苏MYB 转录因子基因进行氨基酸对比,并利用Jalview 软件绘制R2、R3 结构域的氨基酸序列图 (图1)。有49 个糙苏MYB 蛋白中有两个保守的结构域R2 和R3,R2 保守基序类型为-[W] -X (19) -[W] -X (19) -[W] -,含有3 个高度保守的色氨酸残基 (W),在这3 个色氨酸残基之间分别间隔着包括相对保守的精氨酸(R)、脯氨酸 (P)、赖氨酸 (K)、谷氨酸 (E)、半胱氨酸 (C)、亮氨酸 (L) 等19 个氨基酸残基序列。R3 保守基序类型为-[F] -X (18) -[W] -X (18) -[W] -,R3-MYB 的第一个保守色氨酸残基 (W) 被苯丙氨酸 (F)、异亮氨酸(I) 或亮氨酸 (L) 等酸性氨基酸替代。以上结果表明,PuMYB 转录因子结构域具有高度保守性,转录因子N 端的重复序列可以很好地维持H (螺旋) -T (转角) -H (螺旋) 结构,使转录因子与DNA 分子进行结合,控制糙苏基因的转录水平,从而发挥特定功能。

图1 糙苏MYB 氨基酸序列的对比
2.2.2 MYB 转录因子的保守基序预测
使用MEME 预测61 个糙苏MYB 家族成员的蛋白质的保守基序 (图2)。E 值最低的预测保守基序motif 1 长度为50 个氨基酸,且有50 个PuMYB 家族成员有该基序;motif 2 长度为20 个氨基酸,有51 个PuMYB 家族成员有该基序;motif 3长度为21 个氨基酸,56 个PuMYB 家族成员都有该基序;motif 4 长度为11 个氨基酸,有32 个PuMYB 家族成员有该基序;motif 5 长度为11 个氨基酸,拥有该基序的PuMYB 家族成员有21 个;motif 6 长度为11 个氨基酸,拥有该基序的PuMYB家族成员有26 个;motif 7 长度为50 个氨基酸,有3 个PuMYB 家族成员有该基序;motif 8 长度为11个氨基酸,有13 个MYB 家族成员有该基序;motif 9 长度为33 个氨基酸,有4 个PuMYB 家族成员有该基序;motif 10 长度为23 个氨基酸,有6 个PuMYB 家族成员有该基序。另外motif 7 中具有3个极度保守的色氨酸 (W) 位点,2 个位点之间分别间隔19、18 个氨基 酸,在motif 1、motif 3、motif 4、motif 6、motif 10 中也都有W 位点。其中有26 个PuMYB 的基序排列为motif 3、motif 4、motif 1、motif 2。结合图3,61 个糙苏MYB 家族蛋白中都含有W 位点,说明PuMYB 转录因子基因结构域具有高度保守性。

图2 糙苏MYB 转录因子蛋白的保守基序预测

图3 部分糙苏MYB 转录因子家族成员的保守基序排列情况
2.3 MYB 转录因子家族高级结构
通过SOPMA 对糙苏MYB 转录因子蛋白进行分析,结果为61 个PuMYB 中除PuMYB58 只有α-螺旋和无规则卷曲外,其余60 个PuMYB 都含有α-螺旋 (33.89%)、β-折叠 (7.45%)、β-转角(4.60%) 和无规 则卷曲 (54.25%)。选取PuMYB1、PuMYB31、PuMYB38、PuMYB58 进 行展示 (图4),说明PuMYB 以无规则卷曲构成二级结构的主要元件,α-螺旋为次要元件,而β-折叠和β-转角所占的百分比较少。

图4 糙苏MYB 转录因子家族蛋白二级结构预测
选取3 个亚类中具有代表性的糙苏MYB 转录因子蛋白进行同源建模 (Swiss-Model),得出PuMYB1、PuMYB31 是 SEQ 值为 59.09%、54.55%的R2R3 型MYB 的转录因子识别靶DNA的结构 (图5),可知α-螺旋、β-折叠、β-转角和无规则卷曲共同构成了糙苏MYB 蛋白的空间结构,与二级结构的预测结果相符。并且R2R3-PuMYB 类蛋白三级结构相似性较高,R2、R3 结构域中各有3 个α-螺旋,可以看到有HTH 结构的存在[12]。
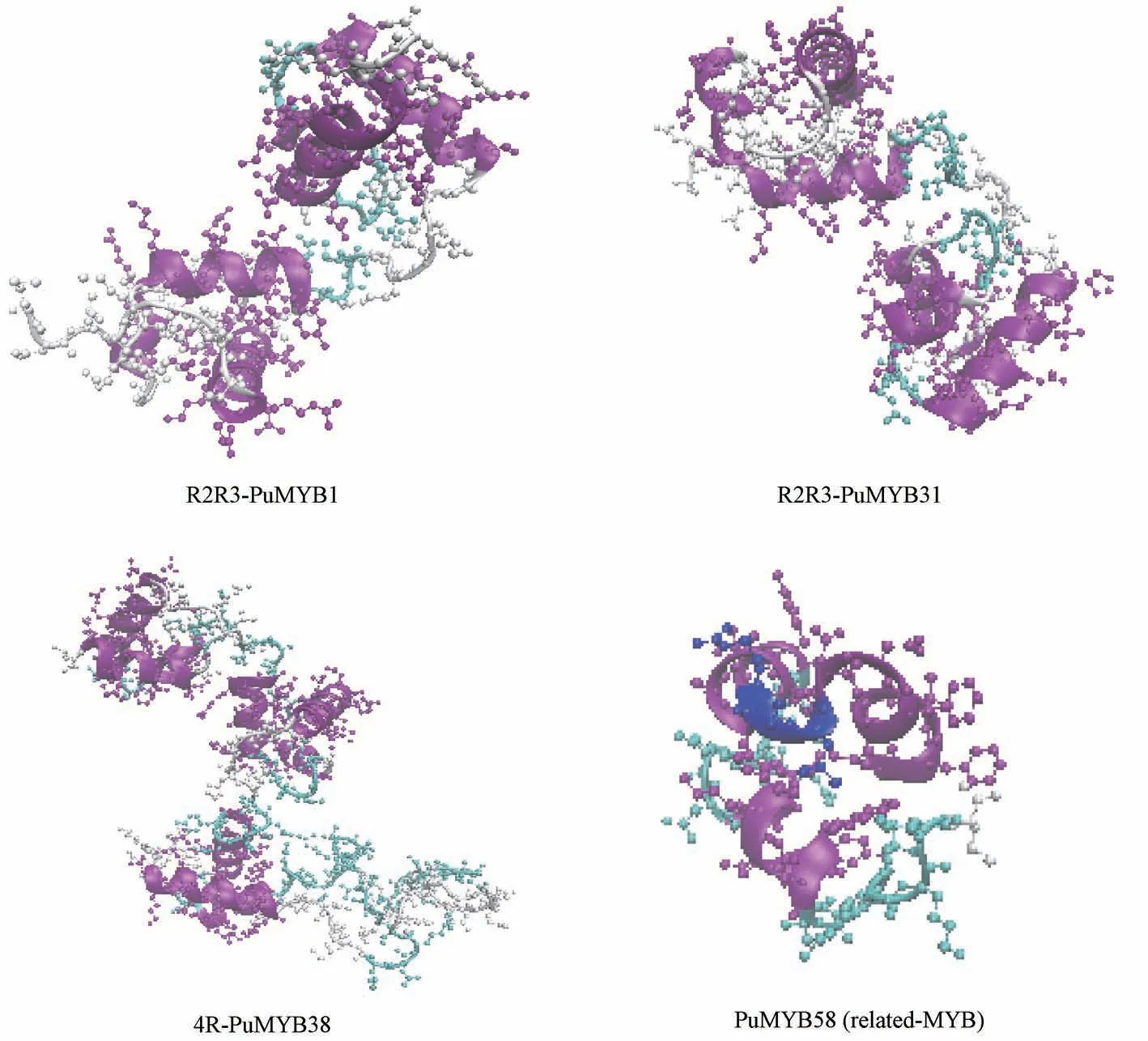
图5 PuMYB 蛋白三级结构的预测
2.4 MYB 基因家族的系统发育树
基于MYB 家族蛋白共分为25 个亚组 (S1~25),其中S15、S5、S6、S10、S16、S12、S17 这7 个亚组中未发现PuMYB 家族蛋白,其余18 个亚组中均有PuMYB 家族蛋白分布。拟南芥MYB 家族蛋白中,S4、S5、S6、S7 亚家族参与花青素和黄酮类化合物的合成途径,其中S4、S7 亚家族中的 ATMYB7、ATMYB11、ATMYB12、ATMYB111可以调控类黄酮类物质的生物合成[13]。所以,同这些蛋白处于同一分支的PuMYB40、PuMYB43 转录因子基因可能在黄酮类物质的生物合成中发挥重要调控作用[14]。
在植物抗逆性方面,拟南芥S1、S22 亚家族在响应非生物胁迫中起着一定的作用,与干旱胁迫和抗逆性有关[15-16],因此,与这些蛋白处于同一分支的PuMYB3、PuMYB44、PuMYB45、PuMYB57、PuMYB60、PuMYB61、PuMYB4、PuMYB19、PuMYB21、PuMYB27、PuMYB28 转录因子可能在响应非生物胁迫中起着一定作用[17]。拟南芥S2 亚家族ATMYB14、ATMYB15、ATMYB13 具有响 应冷胁迫和盐胁迫的作用[18],与这些蛋白处于同一分支的PuMYB6、PuMYB53 转录因子可能具有类似功能。S11 亚家族的ATMYB102、ATMYB41 和ATMYB74 分别有助于植物对食菜青虫产生抗性,并在盐胁迫下起作用,参与不同细胞过程对渗透胁迫的反应,在同一分支下的PuMYB30 转录因子可能有类似功能。S20 亚家族的ATMYB2 控制ABA诱导盐和脱水响应基因,ATMYB108 在生物和非生物胁迫中起作用,推测与这些蛋白处于同一分支的PuMYB7、PuMYB31、PuMYB42 转录因子也具有相似功能 (图6)。综上可知,PuMYB 转录因子基因可能参与糙苏对生物与非生物胁迫的响应过程。

图6 糙苏与拟南芥MYB 家族蛋白的聚类分析
拟南芥S3 亚家族在拟南芥纤维和导管中激活木质素的生物合成,S21 亚家族能够促进纤维细胞中细胞 壁的增 厚[19]。由此推 测,PuMYB48、PuMYB51、PuMYB9、PuMYB17、PuMYB56 可 能参与糙苏木质素的生物合成以及细胞壁的增厚等次生生长过程。S14 亚家族是腋生分枝的部分冗余调节因子,且ATMYB38 在花序发育过程中影响附属侧芽的形成;ATMYB68 是根生长的特异性调节因子,在不利条件下影响植物的整体发育[20]。因此,与拟南芥S14 亚家族在同一分支的 PuMYB2、PuMYB29、PuMYB33、PuMYB50 可能会影响糙苏附属侧芽和植物整体发育过程。
2.5 MYB 转录因子功能
2.5.1 MYB 转录因子的GO 注释
通过QuickGO 在线查询GO 注释信息,糙苏MYB 转录因子序列中分别有59 条、135 条和49 条注释信息被分别注释到生物学过程 (BP)、分子功能 (MF) 和细胞组分 (CC),共富集到16 个功能类别中,在生物学过程中,涉及细胞过程 (44)、生物调节 (7)、定位 (3)、发育过程 (1)、代谢过程 (1)、再合成 (1)、免疫系统过程 (1)、胁迫响应 (1) 等8 个功能类别,主要富集在细胞过程;在分子功能中,主要涉及结合 (83)、催化活性 (45)、转录因子活性 (5) 和分子转导活性(2) 等4 个功能类别,主要富集在结合和催化活性上 (图7);细胞组分中涉及细胞核 (39)、细胞膜 (2)、含蛋白复合物 (5)、病毒成分 (1)、胞外区 (1) 等5 个功能类别,主要表现在细胞核中富集。糙苏MYB 转录因子的结合过程占比最大,MYB 转录因子主要通过结合到DNA 区域发挥相应的调控功能,主要涉及糙苏的细胞过程和生物调节过程,以控制植物的生长过程[21]。

图7 糙苏转录因子的功能注释
2.5.2 次生代谢相关MYB 转录因子功能
以拟南芥MYB 转录因子作为参考,通过STRING 数据库的Muitipie sequence 分析预测糙苏MYB 转录因子的蛋白功能,若同源性越高,功能相同或越相似。结果如表 2 所示,PuMYB7、PuMYB42 编码蛋白与拟南芥转录因子AT3G06490.1 的同源性较高,推测它们参与了茉莉酸信号对雄蕊成熟和雄性育性的调节;PuMYB10、PuMYB34 转录因子推测是C4H 基因的转录抑制因子,参与了辛酸酯的生物合成,与拟南芥转录因子AT5G26660.1 同源性较高;推测与拟南芥转录因子AT5G49330.1 高度同源的PuMYB40转录因子参与黄酮醇的生物合成,主要控制黄酮醇在子叶和叶片中的生物合成;PuMYB43 转录因子与拟南芥转录因子AT4G38620.1 高度同源,与AT1G22640.1、AT2G16720.1 同源性较高,推 测PuMYB43 具有抑制肉桂酸4-羟化酶基因表达和抑制苯丙烷类生物合成基因表达的作用,并参与了黄酮醇生物合成的负调控,可见糙苏MYB 转录因子参与了植物萜类代谢生物途径[22]。

表2 糙苏MYB 转录因子同源性较高的拟南芥MYB 转录因子蛋白功能
2.6 MYB 转录因子表达
利用Tbtools 软件的HeatMap 功能对糙苏MYB转录因子进行聚类分析并绘制热图(图8),其中57个基因在根中表达,有15 个基因的表达量较高;56个基因在叶中表达,有10 个基因的表达量较高;有5 个基因在根和叶中的表达量都很高 (PuMYB18、PuMYB19、PuMYB21、PuMYB36、PuMYB43),44个基因在根中的表达量上调,17 个基因在叶中表达量上调;糙苏黄酮类化合物主要在根中表达,糙苏为多年宿生植物,民间用根入药。PuMYB48、PuMYB51、PuMYB9、PuMYB17、PuMYB56 在以上系统进化树分析中可能参与木质素的生物合成以及细胞壁的增厚等次生生长过程,且在根的表达量中上调,根据表达模式推测,这5 个基因可能参与糙苏根木质素的生物合成和次生生长等过程。
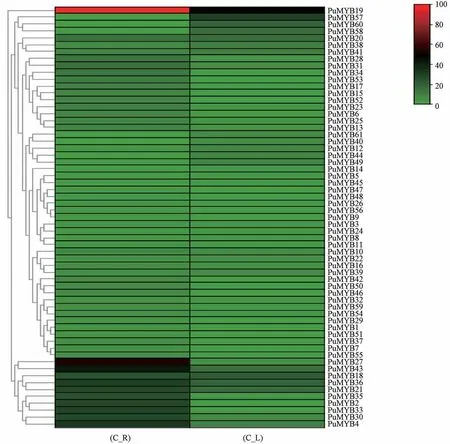
图8 糙苏MYB 转录因子基因的表达热图
3 讨论
MYB 类转录因子成员之间存在着机构和功能的相关性,根据这些相关性,可以在一定程度上预测一个MYB 类新成员的生物学功能。作为一类广泛参与植物生命活动过程的转录因子,MYB 转录因子在次生代谢、非生物胁迫与植物激素应答、环境胁迫响应等方面发挥重要作用[27]。在拟南芥[28]、花生[29]、水稻、黑果枸杞[30]等植物中均已经鉴定出MYB 转录因子蛋白,并在基因水平上进行了深入分析。对糙苏MYB 转录组进行研究,共筛选鉴定出61 个MYB 转录因子,氨基酸数目在98~1 756,可能是由于可变剪切导致了不同基因编码区和非编码区在基因全长的比例呈现多态性。糙苏MYB 转录因子基因的高级结构预测表明,无规则卷曲构成糙苏MYB 家族的主要元件,α-螺旋为次要元件。系统进化分析结果表明,在拟南芥的18 个亚组中发现糙苏MYB 蛋白分布,PuMYB40、PuMYB43 可能参与糙苏黄酮类物质的生物合成,有17 个PuMYB 转录因子可能在糙苏抗逆性胁迫中发挥其作用,PuMYB48、PuMYB51、PuMYB9、PuMYB17、PuMYB56、PuMYB2、PuMYB29、PuMYB33、PuMYB50可能参与糙苏的生长发育过程。
根据GO 注释分析表明,糙苏MYB 转录因子主要通过结合到DNA 区域发挥相应的调控功能,主要涉及糙苏的细胞过程和生物调节过程,控制植物生长过程。糙苏次生代谢相关MYB 转录因子功能分析表明,PuMYB7、PuMYB42 编码蛋白与拟南芥转录因子AT3G06490.1 的同源性较高,推测有对茉莉酸信号响应的作用,可对雄蕊成熟和雄性育性进行调节;推测与拟南芥转录因子AT5G49330.1 高度同源的PuMYB40 转录因子参与黄酮醇的生物合成,主要控制黄酮醇在子叶和叶片中的生物合成;PuMYB43 转录因子可能具有抑制肉桂酸4-羟化酶基因表达和抑制苯丙烷类生物合成基因表达的作用,并参与黄酮醇生物合成的负调控。糙苏MYB 转录因子表达分析显示,PuMYB48、PuMYB51、PuMYB9、PuMYB17、PuMYB56 可 能参与糙苏根的木质素的生物合成和次生生长等过程。
猜你喜欢
福建农林大学学报(自然科学版)(2022年4期)2022-11-01
湖北农业科学(2022年11期)2022-07-18
南方医科大学学报(2022年3期)2022-04-13
园艺与种苗(2020年12期)2021-01-08
实用肿瘤学杂志(2020年4期)2020-12-08
三农资讯半月报(2020年15期)2020-08-25
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
医学综述(2011年12期)2011-12-09
亚热带农业研究(2011年3期)2011-09-29
