神经病理性疼痛小鼠背根神经节中SIRT3表达的变化
2022-08-10 06:28:38任秀花杨庆庆张晶晶张义丹臧卫东
郑州大学学报(医学版) 2022年4期
任秀花,杨庆庆,张晶晶,张义丹,臧卫东
郑州大学基础医学院人体解剖学系 郑州 450001
组蛋白去乙酰化酶Sirtuin3(SIRT3)作为一种烟酰胺腺苷二核苷酸(NAD+)依赖性去乙酰化酶,广泛表达于脑、心脏、肝脏等代谢旺盛的器官组织[1]。研究[2-3]显示,SIRT3可通过激活超氧化物歧化酶和过氧化氢酶,促进线粒体对活性氧的清除,保护线粒体免受氧化应激损伤,与线粒体代谢、细胞死亡、免疫和炎症等生物功能息息相关。背根神经节(dorsal root ganglion,DRG)为初级感觉神经元的胞体所在部位,其氧化应激水平在神经病理性疼痛的发展中起着重要作用[4]。本研究通过免疫印迹和免疫荧光标记法,对小鼠DRG中SIRT3的表达及其在神经病理性疼痛状态下的表达变化进行了探讨,现报道如下。
1 材料与方法
1.1 主要试剂和仪器SIRT3抗体购自Affinity公司,DRG大直径神经元标志物NF200、胶质细胞标志物GS、β-actin抗体以及HRP标记的山羊抗兔IgG抗体和山羊抗鼠IgG抗体购自美国Abcam公司,DRG小直径非肽能神经元标志物IB4和中小直径肽能神经元标志物CGRP抗体购自美国Sigma公司,Cy3标记的山羊抗兔IgG和488标记的山羊抗鼠IgG抗体购自美国Jackson ImmunoResearch公司。多聚甲醛、组织裂解液、蛋白浓度测定试剂盒、抗荧光淬灭剂购自北京索莱宝科技有限公司。Von-Frey丝购自法国Bioseb公司,体视显微镜、正置荧光显微镜购自日本Nikon公司,小动物气体麻醉机购自深圳瑞沃德公司,全自动超薄切片机购自德国Leica公司,生物分光光度计购自德国Thermo公司,电泳、转膜及化学发光成像系统购自美国Bio-Rad公司,低速瞬时离心机、磁力搅拌器购自武汉赛维尔生物科技有限公司。
1.2 实验动物及分组体重22~25 g的健康成年雄性C57BL/6小鼠购自河南省实验动物中心(合格证号DW2020070025)。小鼠购买后正常饲养至少7 d以适应环境(12 h/12 h光照周期,水食自取),采用随机数字表法分为正常组(3只)、假手术组(共12只)和坐骨神经慢性压迫性损伤(chronic constriction injury,CCI)组(12只)。所有动物实验均已获得郑州大学动物保护和使用委员会批准。
1.3 CCI模型的建立使用异氟烷麻醉(体积分数4%异氟烷用于诱导,体积分数2%异氟烷用于维持)健康成年雄性C57BL/6小鼠,剃除手术部位及周围毛发,随后使其处于俯卧位并固定左后肢,使用碘伏对去毛部位进行消毒,在股骨外侧上方纵行切开皮肤以及顺肌纹钝性分离股二头肌,暴露并分离坐骨神经。在坐骨神经主干的中部,用手术缝合线以1 mm 的间距进行3次结扎,结扎强度以引起腿部肌肉轻度颤动为宜。结扎完毕后逐层缝合伤口,伤口再次消毒。假手术组小鼠仅做皮肤切口和肌肉钝性分离,不做坐骨神经结扎。
1.4 各组小鼠机械痛行为学测试采用经典的Von-Frey丝测试法(0.07 g和0.40 g)进行机械痛行为学测试。假手术组和CCI组小鼠分别于术前1 d(-1 d)、术后7 d进行测试。每次测试前将小鼠随机放置于测试位适应30 min,安静(不来回走动,舔爪)后,先用0.07 g Von-Frey丝垂直作用于小鼠掌跖部无毛处(Von-Frey丝呈“S”形,作用时间6~8 s,相邻刺激间隔至少7 s,每只脚测量10次)。0.07 g丝测试完毕30 min后开始进行0.40 g Von-Frey丝测试。作用时间内,小鼠缩足或舔舐被刺激侧记为阳性反应。机械痛阈值为10次刺激中阳性反应的百分比,即机械缩足频率(paw withdrawal frequency,PWF)。
1.5 各组小鼠DRG中SIRT3蛋白表达的Western blot检测术后7 d使用体积分数4%异氟烷深度麻醉小鼠后快速断头处死并取出L3~5腰椎DRG。每组6只小鼠,每2只共6个术侧L3~5DRG合为一个测试样,用于后续检测(因单只动物组织量过小,无法检测,故合并标本进行检测)。向每个样本加入65 μL裂解液,使用低温研磨仪充分研磨,1 000 r/min 4 ℃离心10 min并取上清液,使用蛋白浓度检测试剂盒检测蛋白浓度;取30 μg总蛋白进行电泳,260 mA恒流转膜1.5 h,含50 g/L牛血清白蛋白的TBST溶液封闭2 h,随后使用SIRT3和β-actin一抗(均按1∶1 000稀释)4 ℃摇床孵育过夜;TBST洗膜3次后使用山羊抗兔二抗或山羊抗鼠二抗(均按1∶10 000稀释)孵育2 h;TBST洗膜3次;使用ECL发光试剂盒曝光成像。使用Image J软件分析,以目的条带灰度值/内参条带灰度值的比值为目的蛋白相对表达量。
1.6 SIRT3在DRG中神经元的定位使用体积分数4%异氟烷深度麻醉小鼠3 min,随后快速打开小鼠胸腔,暴露心脏。使用输注器于左心室进针,并于右心耳剪口用于液体流出,使用注射用生理盐水快速灌流至小鼠右心耳造口无血液流出,肝脏变为乳白色。随后使用预冷的40 g/L多聚甲醛溶液灌流20 min,灌流结束后取术侧L3~5DRG组织,置于40 g/L多聚甲醛中4 ℃过夜固定。固定后组织常规脱水及包埋,4 μm厚切片,每个蜡块连续切片6张,60 ℃烤片2 h备用。经常规脱蜡至水的切片使用ddH2O清洗3遍,使用柠檬酸缓冲液(pH 6.0)进行高温抗原修复,使用PBS清洗3遍,经体积分数10%的山羊血清封闭1 h后,分别用SIRT3(抗体按1∶200稀释)、SIRT3+NF200(抗体按1∶500稀释)、SIRT3+IB4(抗体按1∶500稀释)、SIRT3+CGRP(抗体按1∶500稀释)、SIRT3+GS(按1∶500稀释)进行免疫标记(4 ℃过夜)。随后加入对应的荧光二抗(Cy3标记的羊抗兔IgG或488标记的羊抗小鼠IgG抗体,均按1∶300稀释),避光室温孵育2 h,PBS漂洗3次,10 min/次。DAPI(按1∶200稀释)衬染胞核,使用抗荧光衰减封片剂封片,荧光显微镜下观察拍照。每张切片使用Imaging J软件进行阳性细胞数的统计。
1.7 统计学处理采用Graphpad Prism数据分析软件。应用重复测量数据的方差分析比较不同时间点各组小鼠机械缩足频率的差异,应用单因素方差分析比较各组小鼠SIRT3蛋白表达水平的差异,组间两两比较使用Sidak′s多因素检验。检验水准α=0.05。
2 结果
2.1 SIRT3在正常小鼠DRG神经元中的定位免疫荧光双标结果显示,在正常小鼠DRG中,SIRT3与NF200、CGRP和IB4均有共标(图1A~C),且定位于细胞质中。SIRT3与GS不共标(图1D)。SIRT3与各类神经元标志物共标率的统计结果显示:分别有95.7%的NF200阳性神经元(图1E)、79%的CGRP阳性神经元(图1F)和80.5%的IB4阳性神经元与SIRT3共标(图1G);在SIRT3阳性神经元中,分别有46.4%、51.9%和36%的神经元与NF200、CGRP和IB4共标(图1E~G)。该结果提示,SIRT3广泛分布于小鼠外周神经系统的各类神经元而非胶质细胞中。

A~D:SIRT3分别与NF200、CGRP、IB4和GS双标示例;蓝色为DAPI指示的细胞核;E~G:SIRT3和NF200、CGRP、IB4的免疫荧光阳性率以及共标率统计图
2.2 假手术和CCI组小鼠机械痛阈值的比较见表1。由表1可知,CCI组小鼠术后7 d机械缩足频率较术前升高,且高于假手术组,说明CCI手术导致小鼠术侧足底出现了显著的机械痛觉敏化,模型建立成功。

表1 假手术和CCI组小鼠机械痛阈值的比较
2.3 2组DRG中SIRT3蛋白表达的比较术后7 d,与假手术组(1.00±0.08)相比,CCI组小鼠DRG中SIRT3蛋白(0.46±0.18)表达量降低(t=3.882,P=0.018)。
2.4 2组不同类型DRG神经元中SIRT3的表达变化见表2和图2。神经损伤诱导的神经病理性疼痛小鼠DRG中SIRT3的下降主要发生于大直径神经元和小直径非肽能神经元中。

表2 假手术组与CCI组小鼠不同类型DRG神经元中SIRT3表达的比较 %
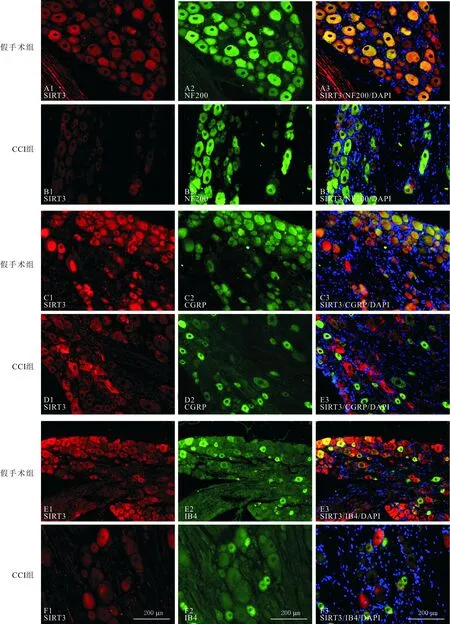
A~F:假手术组和CCI组小鼠DRG中SIRT3分别与NF200、CGRP以及IB4的双标结果
3 讨论
初级感觉神经元(伤害性感受器)在痛觉传导通路中负责感知外界伤害性刺激类型、强度、位置并将其转化为电信号,其功能是保证痛觉感知可正常进行的关键因素。神经损伤、炎症、化疗等刺激均可导致DRG神经元发生如离子通道表达、线粒体损伤、氧化应激等变化而使其处于过度兴奋状态,参与多种慢性疼痛的发生发展[5-8]。
DRG是初级感觉神经元的胞体聚集部位。SIRT3作为Sirtuin家族中主要成员之一,主要定位于线粒体,通过参与线粒体代谢,对多种细胞生理状态起着重要调控作用[9-12]。我们的研究结果显示SIRT3在DRG的NF200阳性的有髓神经元、CGRP阳性的肽能伤害性感受器以及IB4阳性的非肽能伤害性感受器中均有广泛表达;在CCI诱导的神经病理性疼痛状态下小鼠DRG中SIRT3表达减少,且SIRT3表达减少主要发生在大直径神经元和小直径非肽能神经元中,提示SIRT3可能通过影响这两类神经元的功能而参与神经病理性疼痛。
随着近年来针对SIRT3的研究逐渐深入,越来越多的研究成果[7,11]证明,SIRT3可通过调节能量代谢和氧化应激而参与如糖尿病、癌症、神经退行性疾病等多种疾病的发生发展,SIRT3在疾病发生过程中的作用机制也开始被深入探索。作为机体内功能活动最为旺盛、ATP需求量最大的细胞之一,神经元的生存与活动均极大地依赖于线粒体的正常活动。线粒体功能异常可导致神经元ROS生成增多,最终导致神经元凋亡[13]。有研究[14]表明,SIRT3可以通过抑制线粒体氧化应激来控制细胞存活、代谢和应激反应。SIRT3敲除可使线粒体出现严重的呼吸酶缺陷,导致细胞内ATP水平降低,提示SIRT3可通过去乙酰化激活呼吸酶而促进线粒体功能[15]。此外,SIRT3敲除小鼠表现出蛋白乙酰化水平增加,超氧化物歧化酶2活性降低,氧化还原稳态被破坏[15-16]。相反,SIRT3过表达可抑制氧化应激导致的线粒体Ca2+超载,保护神经元免受氧化应激损伤[17]。
在肾脏疾病中的研究[18]显示,近端肾小管SIRT3表达下降和线粒体功能障碍是肾小管损伤与应激的发生因素之一;在急性脑缺血过程中,SIRT3蛋白表达下降可通过破坏线粒体功能导致神经元损伤[19]。结合我们的研究结果,在CCI诱导的神经病理性疼痛状态下,大直径神经元和小直径非肽能神经元中SIRT3阳性细胞比例减少,我们高度假设SIRT3的减少可能参与了大直径神经元和小直径非肽能神经元的线粒体损伤,进而介导小鼠的痛觉敏化。
我们将进一步在不同的疼痛模型中上调或下调SIRT3,继而观察对疼痛的行为学影响,以明确SIRT3是否是参与疼痛的重要分子,以及其影响下游的分子机制。同时,我们也将进一步针对SIRT3在神经病理性疼痛的发生发展乃至治疗中的作用进行深入研究,以期为将SIRT3作为神经病理性疼痛的治疗靶点转化为安全可靠的临床治疗手段提供充分的依据。
猜你喜欢
海洋通报(2021年1期)2021-07-23 01:55:14
生物学通报(2021年4期)2021-03-16 05:41:26
世界科学技术-中医药现代化(2020年2期)2020-07-25 02:05:56
基层中医药(2018年11期)2019-01-31 05:27:00
科普童话·神秘大侦探(2017年4期)2017-04-06 20:37:47
西南军医(2016年6期)2016-01-23 02:21:19
汽车生活(2015年6期)2015-05-30 04:59:21
西南军医(2015年2期)2015-01-22 09:09:37
癌变·畸变·突变(2014年1期)2014-03-01 04:39:36
食品科学(2013年15期)2013-03-11 18:25:48