
环割对马家柚蔗糖和柠檬酸含量及其相关基因表达的影响
2022-08-10 05:47:16赵连鑫牟德生郭艳兰史星雲韩登山姚元文
江西农业学报 2022年5期
赵连鑫,王 鑫,何 彩,牟德生,郭艳兰,史星雲,韩登山,姚元文
(1.武威市林业科学研究院,甘肃 武威 733000;2.华中农业大学 园艺林学学院,湖北 武汉 430070;3.湖州市农业科技发展中心/湖州市农业科学研究院,浙江 湖州 313000)
环割是果树栽培管理中常用的一种技术措施,具有保花保果和提高果实品质的作用。环割时间和圈数是影响环割效应的主要因素,笔者前期对不同环割时间、环割圈数对马家柚(Citrus grandisL.Osbeck)环割试验表明,不同环割处理对马家柚果实糖酸的影响显著[1]。柑橘果实中主要含有蔗糖和果糖等[2],其中,柚类果实中蔗糖含量最高,糖含量的变化与糖代谢途径中相关的酶有着紧密的联系,因而决定果实中糖含量的酶有很多。
20世纪90年代,在玉米和菠菜中对植物细胞质进行纯化得到了蔗糖磷酸化酶SPS[3],它可催化果糖转化为蔗糖,在香瓜中也证实了这一点[4]。蔗糖转运蛋白(SUC)是质膜上转运蔗糖的载体,与蔗糖的装卸有密切关系[5-6]。SUC是一个多成员的基因家族[7],根据不同的性质将其分成3个亚族:SUC1、SUC2、SUC3和SUC4,它们在植物的不同组织和器官中存在着不同的表达。有机酸的代谢是一个复杂的过程,有很多相关的酶参与其中,不同的酶在果实的有机酸代谢中起着不同的作用。柑橘果实存在着丰富的有机酸,主要有柠檬酸、苹果酸和乙酸等[8-9],其中柠檬酸是柑橘果实中含量最多的有机酸。文涛等[10]在脐橙果实酸代谢研究中发现,CS的表达与柠檬酸的代谢存在着正相关,但也有研究发现[11],CS的表达与果实柠檬酸的含量无关,说明在不同品种中相同的基因有着特异性的表达,这也决定了在不同的果树品种中研究每个基因的必要性。ACO又称乌头酸水合酶,研究发现在果实中它具有2种形态:线粒体ACO和胞质ACO[12],其表达量与柠檬酸密切相关[13],在甜来檬研究中发现,它与柠檬酸的含量呈负相关[14]。
为了解环割对柚子果实糖、酸含量变化的原因,本研究以马家柚为试材,设置不同环割时间和圈数处理,选择与糖酸相关的SUC1、SUC2、SUC3、SUC4、CS、ACO等6个基因,研究了环割对马家柚果实中糖、酸(蔗糖、柠檬酸)含量的变化及相关基因的表达情况,以期明确环割后相关基因的表达与糖酸含量变化的关系,从而分析了环割措施对提高果实品质的原因。
1 材料与方法
1.1 材料及处理
试验地概况:试验地设在江西省广丰区瀛洲农业公司基地,位于江西省上饶市广丰区,属亚热带季风气候,年平均无霜期266 d,年平均气温17.9℃,年平均降雨量1661.6 mm,土壤以红壤、黄壤为主。基地果园内只栽植马家柚,浇水、施肥和病虫害防治等统一管理。
试验材料选择树势、挂果量一致的10年生柚树,在第1次生理落果期(5月15日)和第2次生理落果期(6月15日)进行环割处理,设环割1、2、3圈处理,每株只环割3个粗度一致的主枝,在主枝基部处进行环割,环割刀口宽度1 mm,圈距2 cm,每个处理3株。果实成熟后(11月15日),采集环割枝上同一方位、同一大小、同一成熟度的果实,取果实中部果肉液氮处理后,用于糖酸含量和糖酸相关基因表达量的测定。
1.2 试验方法
1.2.1 糖酸含量的测定 果实中糖和酸采用气相色谱仪(Agilent 6890N)测定,具体方法参考文献[15];色谱条件:HP-5色谱柱,分流/不分流进样口温度270 ℃,检测器温度300 ℃;气体流速:高纯N2作载体,流量为45 mL/min;H2的流量为40 mL/min,空气流量为450 mL/min,柱头压12.00 psi,进样量1 μL,分流比30∶1,分流流速60.1 mL/min;在升温程序中,初温130 ℃,以8 ℃/min升至152 ℃,12℃/min升至176 ℃,16 ℃/min升至198 ℃,20 ℃/min升至238 ℃,24 ℃/min升至280 ℃,280 ℃保留4 min,最大温度不超过325 ℃。
1.2.2 糖酸相关基因筛选及引物设计 从NCBI数据库中,筛选出与柑橘糖、酸代谢相关基因序列7条,其中蔗糖磷酸化酶1条(SPS)、蔗糖转运因子3条(SUC1/SUC3/SUC4)、乌头酸水合酶1条(ACO)、柠檬酸合成酶1条(CS4),用Primer Express 3.0软件设计定量引物,以Actin为内参基因,其中内参基因及所选定量引物序列如表1所示。

表1 糖酸代谢相关基因引物用于Real-time PCR的引物
1.2.3 qRT-PCR测定 用中量法提取果实中的RNA,方法参考徐文欣[16]的研究,再经qRT-PCR测定,测出糖酸相关基因的表达量。
1.3 数据统计分析
采用Excel 2007、SAS软件(SAS System Release 8.1)进行数据统计和差异显著性分析,采用Primer Express 3.0软件设计糖酸相关基因引物。
2 结果与分析
2.1 果实蔗糖和柠檬酸的测定
由表2可知,不同环割处理果肉蔗糖含量均高于对照,蔗糖含量由高到低依次为:Ⅱ-1>Ⅰ-3>Ⅱ-3>Ⅱ-2>Ⅰ-1>Ⅰ-2>CK,Ⅱ-1、Ⅰ-3和Ⅱ-3处理的蔗糖含量较对照分别提高33.5%、13.7%、11.3%,差异显著。果实中柠檬酸的含量由高到低依次为:Ⅱ-1>Ⅰ-3>Ⅰ-1>CK>Ⅱ-2>Ⅰ-2>Ⅱ-3。与CK相比,Ⅱ-1处理的柠檬酸含量最高,比对照高44.1%,差异显著,Ⅱ-3处理的柠檬酸含量最低。
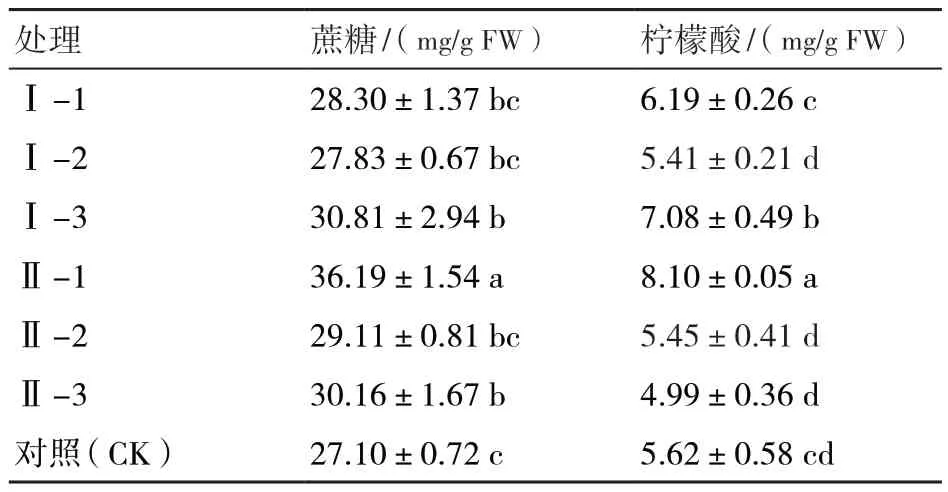
表2 环割处理后果实中可溶性糖、有机酸含量比较
2.2 对糖相关基因表达的影响
图1显示了马家柚果实成熟期蔗糖相关基因的表达情况,A、B、C、D分别为CsSPS(蔗糖磷酸化酶)、CsSUC1、CsSUC3和CsSUC4(蔗糖转运酶)。
图1A显示了各环割处理对马家柚果实成熟期(180 DAF)CsSPS表达的影响。与CK相比,不同环割处理对CsSPS的表达量有升高也有降低的作用,Ⅱ-3处理对CsSPS的表达量最大,比对照高54%;Ⅰ-2和Ⅱ-1处理的表达量最低,比对照低63%和30%,其他处理与对照差异不显著。各处理CsSPS的表达量与该处理下的蔗糖含量不一致。
图1B显示了各环割处理对马家柚果实成熟期(180 DAF)CsSUC1表达的影响。与CK相比,Ⅱ-3处理显著提高了CsSUC1的表达量,提高了144.1%,其他环割处理也提高了CsSUC1的表达量,与该处理下蔗糖的含量表现较为一致。
图1C显示了每种环割处理对马家柚果实成熟期(180 DAF)CsSUC3表达的影响。与CK相比,不同环割处理对CsSUC3的表达有升高也有降低的作用,与各处理下蔗糖的含量不一致。
图1D显示了各环割处理对马家柚果实成熟期(180 DAF)CsSUC4表达的影响。与CK相比,不同环割处理对CsSUC4的表达有升高也有降低的作用,各处理中CsSPS的表达量与该处理下蔗糖的含量不一致。
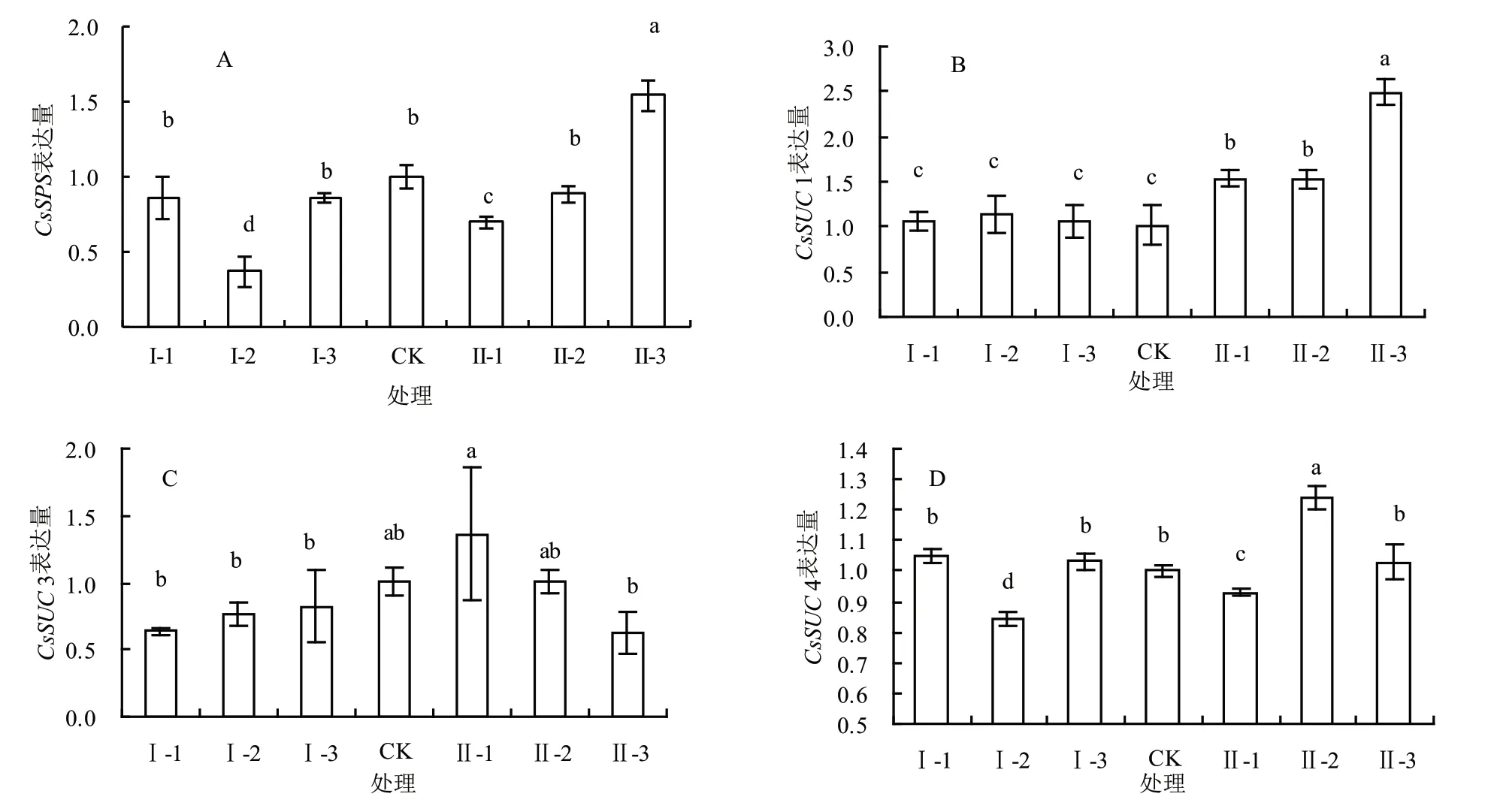
图1 不同环割处理下果实中蔗糖相关基因表达量的比较
2.3 对酸相关基因表达的影响
图2显示了马家柚果实成熟期(180DAF)柠檬酸代谢相关基因的表达情况,本课题组的前期研究表明:CsCS4(柠檬酸合成酶)和CsACO1(乌头酸水合酶)2个基因在柠檬酸代谢途径中具有重要作用。
图2A显示了环割处理对马家柚果实成熟期(180 DAF)CsCS4表达的影响。与CK相比,各环割处理对CsCS4表达有影响,Ⅰ-2处理下的CsCS4表达量显著高于其他处理和CK,与该处理下柠檬酸含量表现不一致。
图2B显示了环割处理对马家柚果实成熟期(180 DAF)CsACO1表达的影响。与CK相比,不同的环割处理对CsACO1的表达有影响,Ⅱ-3处理显著提高了CsACO1的表达量,高于对照26%,Ⅱ-1处理显著降低了CsACO1的表达量,低于对照13%,与该处理下果实中柠檬酸含量较一致;其他处理对CsACO1的表达量,与该处理下果实中柠檬酸含量表现也较为一致。
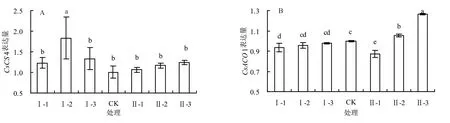
图2 不同环割处理下果实中柠檬酸相关基因表达量的比较
3 讨论与结论
环割作为一种常用的栽培技术手段,已广泛应用于生产,其对果实品质的影响主要表现在对糖、酸含量的变化上,相关的研究甚多[17-18],具有升糖、降酸的效果,且可溶性糖上升的效果最显著。本研究表明:不同的环割处理对马家柚果实中蔗糖和柠檬酸含量均有影响,各环割处理均提高了果实中蔗糖的含量,Ⅱ-1、Ⅰ-3 和Ⅱ-3 处理的蔗糖含量分别较CK 高33.5%、13.7%、11.3%,且差异显著。果实中柠檬酸含量中Ⅱ-1 处理的柠檬酸含量最高,比CK 高44.1%,且差异显著;Ⅱ-3 处理的柠檬酸含量最低,这与涂美艳等[17]的研究结果一致,即环割对可溶性糖与总酸含量的表现一致。
另外,本研究还表明,环割对马家柚蔗糖和柠檬酸相关基因表达的影响,与CK相比,环割处理后CsSPS、CsSUC1、CsSUC3和CsSUC4这4种基因的表达有升高也有降低的作用,其中,环割处理后CsSUC1的表达量与各处理下的蔗糖含量基本一致,其他3个蔗糖相关基因表达与蔗糖含量不一致。因此认为,环割处理可能对CsSUC1表达的影响较大,从而改变了果实中蔗糖的含量。环割对柠檬酸代谢相关基因的研究结果表明:环割对马家柚果实中柠檬酸相关基因表达有影响,其中,环割处理后CsACO1的表达量与各处理柠檬酸含量基本一致,因此认为,环割处理中可能对CsACO1表达的影响较大,从而改变了果实中柠檬酸的含量。
综上所述:环割处理果肉中蔗糖含量均高于对照,Ⅱ-1、Ⅰ-3和Ⅱ-3处理蔗糖含量较对照分别提高33.5%、13.7%、11.3%,差异显著。第2次生理落果期环割1圈果实中柠檬酸含量最高,高于对照44.1%,差异显著;第2次生理落果期环割3圈果实中柠檬酸的含量最低。环割处理对蔗糖和柠檬酸相关基因的表达影响显著,其中环割处理引起CsSUC1和CsACO1表达量的变化,可能与马家柚果实中蔗糖和柠檬酸含量变化密切相关。
猜你喜欢
科学大众(2022年12期)2022-02-18 11:17:38
河北果树(2021年4期)2021-12-02 01:14:38
江苏调味副食品(2021年1期)2021-04-01 12:51:12
房地产导刊(2020年9期)2020-10-28 08:37:24
中国果树(2020年2期)2020-07-25 02:14:18
浙江柑橘(2016年4期)2016-03-11 20:13:01
应用化工(2015年2期)2015-07-13 03:12:26
山西果树(2014年6期)2015-03-12 20:40:56
应用化工(2014年1期)2014-08-16 13:34:08
长江大学学报(自科版)(2014年4期)2014-03-20 13:20:35
