
内生木霉对樟子松幼苗生长及抗病能力的影响*
2022-08-06 00:49魏金龙宋小双刘艳红姜瑞凤平晓帆王新莹遇文婧
林业科技 2022年4期
魏金龙 宋小双 周 琦 邓 勋 刘艳红 姜瑞凤 平晓帆 王新莹 遇文婧**
(1.黑龙江省森工农化有限公司,黑龙江 哈尔滨 150324;2.黑龙江省林业科学院森林保护研究所,黑龙江 哈尔滨 150040;3.黑龙江省森林植物园,黑龙江 哈尔滨 150040)
樟子松(Pinus sylvestris var.mongolica)是三北地区治沙造林最多的针叶树,截至“九五”末期,我国三北地区樟子松人工林已达50~60 万hm2[1]。但随着人工林面积逐年扩大,樟子松病害危害程度日趋严重,最严重的樟子松枯梢病(Sphaeropsis sapinea)发病面积已达1 300 hm2,发病率100%,感病指数44.8,使樟子松的胸径、材积和树高等逐年下降,人工林生产量下降40%,减少商品材积65%,造成巨大的经济损失[2]。樟子松成林后,植株高大,山高林密,大大增加了防治工作的难度。因此,在苗圃期对其进行品质改良,提高其抗逆性和生长优势,不仅能提高苗木产量,还将为营造健康林地环境起到非常重要的作用。
近年来,木霉(Trichoderma spp.)作为一种新兴的微生物肥料越来越受到重视。木霉不仅能通过抗生、重寄生、营养竞争等机制对抗病原真菌,而且能分泌一些小分子蛋白诱导植物产生系统抗病性(Induced Systemic Resistance,ISR)[3],从而促进生长。木霉分为外生木霉和内生木霉,外生木霉是典型的土壤及根际菌,不能定植在植物组织内,作为肥料施用简单,适合在农业上应用;内生木霉定植在植物组织内,受外界环境因素影响较小,可长期发挥促生长、增强抗逆性的作用[4]。因此,利用内生木霉在林木苗期施用,可提高寄主抗逆性,不仅避免成林后林木病害防治难度大的问题,也避免连年施用化学药剂导致土壤肥力下降、环境污染等问题,达到持续、有效控制病害的目的。
本研究从黑龙江省牡丹江市江山娇实验林场健康樟子松组织中分离获得一株内生哈茨木霉(T.harzianum)Th30 菌株,并对樟子松出土苗根施互作,通过研究内生木霉Th30 菌株对樟子松幼苗生长生理指标、抗病指数、抗病基因等影响,探讨内生木霉Th30 菌株对樟子松幼苗促生抗病能力,旨在为樟子松苗圃提供微生物菌肥提供理论依据和物质基础。
1 材料与方法
1.1 试验材料
内生哈茨木霉Th30菌株分离自黑龙江省牡丹江市江山娇实验林场健康樟子松组织。樟子松苗期立枯病病原菌-立枯丝核菌(Rhizoctonia solani,Rs)和樟子松种苗由黑龙江省森林保护研究所病理实验室提供。
1.2 育苗及根施处理
将种子置于铺上湿纱布的育苗盘中,湿纱布覆盖,25 ℃恒温培养箱中培养,每日浇水,发芽后移至装有灭菌土的花盆中,生长2 个月后再移苗。将移栽苗划分为处理组和对照组,处理组分为3 个重复组,每组至少有樟子松出土苗500 株。在处理组,分别选择浓度为1×103个/mL(T1)、1×105个/mL(T2)和1×107个/mL(T3)孢子悬浮液,用200 mL/株水稀释混匀,纱布过滤后,对出土苗进行根施,每隔10 天在根部施用1 次,共3 次。施用时将根围表土移开3 cm 厚,菌肥悬液浇于植物根部,将移开的土壤重新覆盖。对照组用清水浇灌(CK)。
1.3 生长生理指标测定
分别在0、15、30、60 天时测量幼苗的株高、地径、根系、叶鲜重和干重,并随机采集生长状态一致的樟子松针叶,常规方法测定针叶中生理生化指标:蛋白质含量(考马斯亮蓝法),脯氨酸含量(茚三酮显色法),保护酶超氧化物歧化酶SOD(NBT光照化学还原法)和防御酶苯丙氨酸解氨酶PAL(催化L-苯丙氨酸裂解为反式肉桂酸和氨法)活性。并对不同处理的樟子松1 年生和2 年生幼苗的生长进行观测。
1.4 抗病指数调查
对生长3 个月的幼苗采用轻微伤根法接种立枯丝核菌,60 天后调查樟子松各种病害发病率和发病指数。以上实验每个时间点3 个重复,每个重复测定30 株苗。发病率(%)=针叶病变植株总数/调查植株总数×100。根据针叶变色长度占针叶长度的比例(x)将病情指数分为1~5 级(表1)。病情指数=∑(该级代表数值×该病级株数)/(调查总株数×最高级的代表数值)×100。

表1 樟子松幼苗病害分级标准
1.5 RT-qPCR 分析樟子松水杨酸和茉莉酸信号中功能基因的转录水平
设置对照组CK(清水浇灌),处理组T(105个/mL内生菌孢子悬浮液)诱导樟子松幼苗15 天,起苗将根洗净,采用TRIzol 方法提取根部总RNA,进行转录组测序。通过GO 功能富集和KEGG 通路等对差异表达基因进行功能注释,筛选到水杨酸转导路径中的病程相关基因非表达子基因(NPR1)、茉莉酸转导路径中的茉莉酸抗性基因(JAR1)和内参基因(Tubulin alpha),在NCBI 网站上进行BlastX序列对比确定基因序列,利用Primer 6.0 引物设计软件进行引物设计,由上海生工生物技术服务有限公司合成(表2)。在内生木霉诱导0、5、15、30、60 天后选取生长状态一致的樟子松幼苗,每个时间点取3 个重复,每个重复30 株苗。收集根,用液N 速冻后,利用CTAB 方法分别提取根RNA,反转录,以稀释10 倍后的不同cDNA 为模板,用上述3 组引物进行实时定量PCR(RT-qPCR),每组扩增均设3个重复。并采用2-ΔΔCt计算法对数据进行析处理并绘制成曲线图,以此来检测樟子松功能基因在内生木霉定殖后的表达模式。
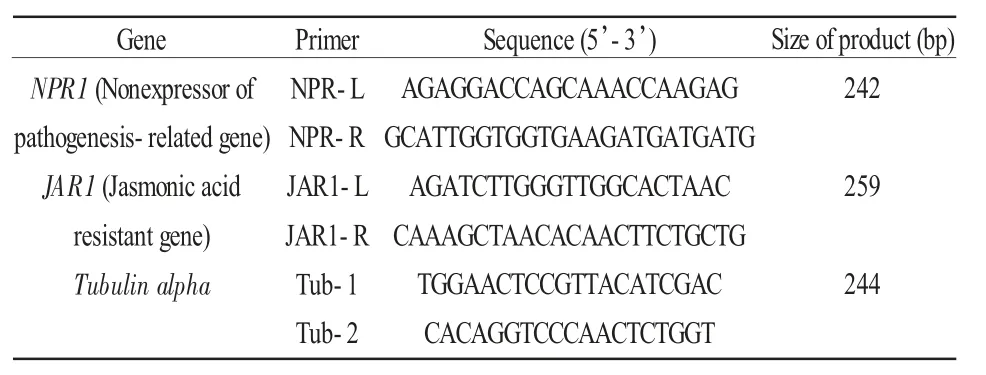
表2 RT-qPCR 引物
2 结果与分析
2.1 内生木霉对樟子松生长指标的影响
根施内生木霉Th30 菌株60 天后,T3(1×105个/mLTh30 孢子悬浮液)处理组的樟子松幼苗的株高、地茎、根长、鲜重和干重均显著高于对照(P<0.05),分别为对照的56.42%、33.33%、26.78%、83.23%和50.60%(表3)。T3 处理组的樟子松1 年生和2 年生植株均明显高于其他处理植株(图1)。因此,内生木霉Th30 菌株能够提高樟子松幼苗的生物量,并显著促进其生长。

图1 内生木霉Th30 对樟子松幼苗生长的影响
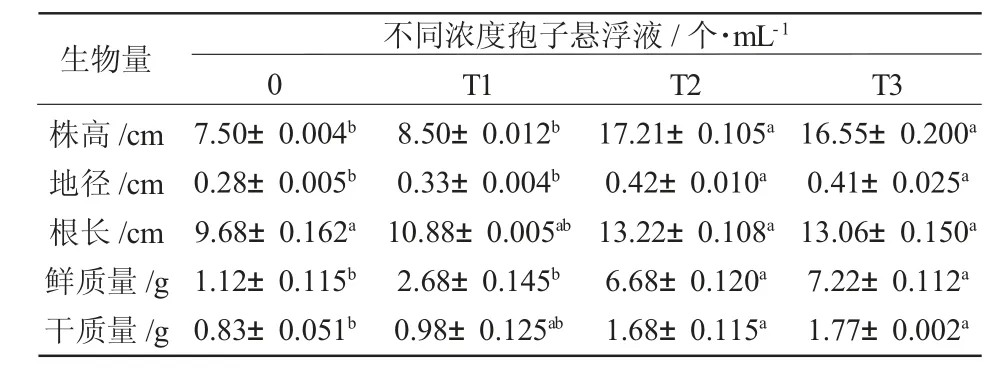
表3 内生木霉菌Th30 对樟子松幼苗生物量的影响
2.2 内生木霉对樟子松生理指标的影响
根施内生木霉Th30 菌株60 天后,樟子松幼苗的不同生理指标(蛋白质含量、脯氨酸含量、SOD和PAL 活性)随着时间的变化而不同(图2)。与对照相比,T2 和T3 处理组的可溶性蛋白质含量变化较显著,在15 天时达到峰值,分别为对照组的8.48和3.71 倍;与对照相比,T1、T2 和T3 处理组的脯氨酸含量均显著升高,在30 天时达到峰值,分别为对照组的2.82、4.29 和5.64 倍;T1 和T3 处理组的SOD 活性在15 天时达到峰值,分别为对照的2.19和1.33 倍,而T2 处理组的SOD 活性在30 天时达到峰值,为对照组的3.69 倍;T1、T2 和T3 处理组的PAL 活性都在30 天时达到峰值,分别高于对照的0.87、1.89 和1.37 倍。因此,内生木霉Th30 菌株在初期能够显著提高樟子松幼苗的可溶性蛋白质含量、脯氨酸含量、以及SOD 和PAL 活性。

图2 内生木霉对Th30 对樟子松生理指标的影响
2.3 内生木霉对樟子松抗病指数的影响
随着时间的推移,不同处理下植株病情指数均上升(表4),但发病程度不同(图3)。接种初期,病情指数不显著(P<0.05),但是接种10 天的CK 组针叶开始出现轻微病斑,而T1 组针叶无明显变化;接种30 天,CK 组与T3 组之间的病情指数差异显著(P<0.05),CK 组病情指数稳定上升且针叶基本变黄,而T3 组的病情指数基本平稳,且针叶发病程度明显滞缓。接种60 天时,CK 组的发病率显著高于处理组,其中T3 组的发病率最低,且CK组针叶全部枯黄,而T3 组的针叶无明显变化。说明接种内生木霉可以提高樟子松的抗病能力。
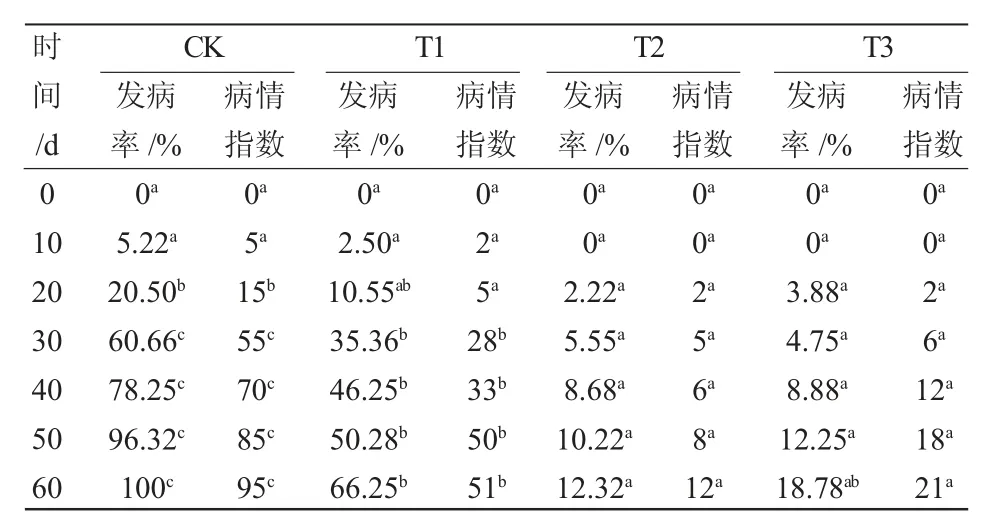
表4 樟子松发病率及病情指数
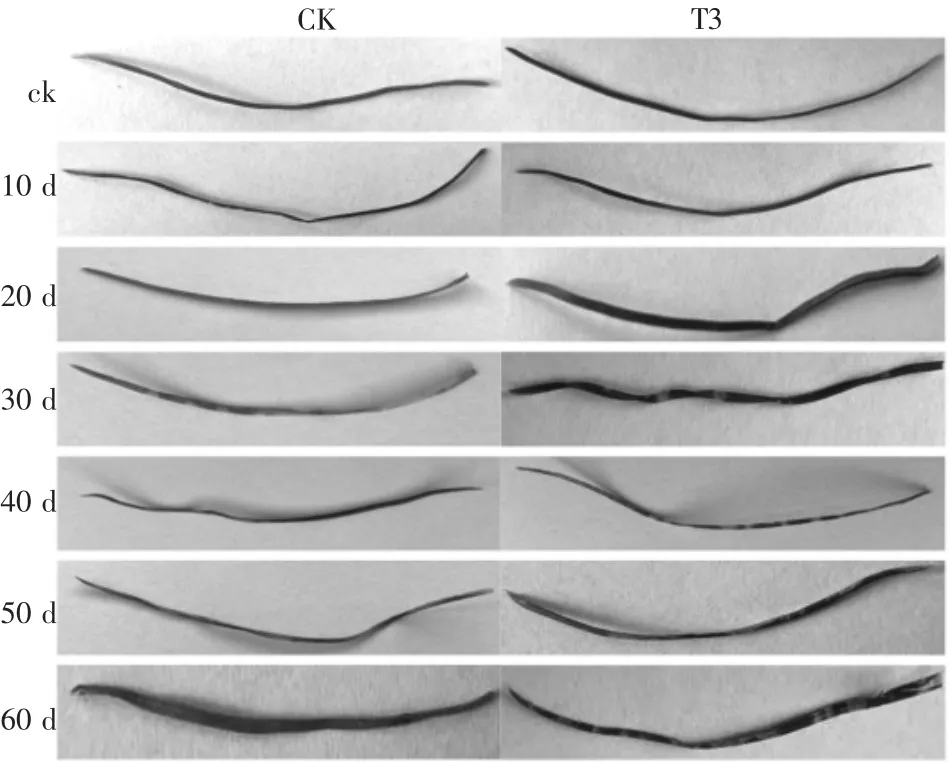
图3 接种病原菌后针叶感病变化
2.4 内生木霉对樟子松抗性基因的影响
经RT-qPCR 分析,在内生木霉Th30 菌株定植过程中,樟子松根系中病程相关基因非表达子基因(NPR1)和茉莉酸抗性基因(JAR1)表达水平如图4和图5。结果显示,CK 组根系中NPR1 无明显变化,T3 组根系中NPR1 在诱导5 天呈负调控,低于对照4 389.98(212.10)倍,之后一直呈正调控,15 天时为表达高峰,为对照的1 772.16(210.75)倍(图4)。CK 组根系中JAR1 无明显变化,T3 组根系中JAR1呈正调控,在5 天后显著增高,在15 天时达到高峰,为对照的5 007.93(212.29)倍(图5)。这说明内生木霉Th30 菌株在定植樟子松幼苗初期能够诱导樟子的NPR1 基因和JAR1 基因的表达。
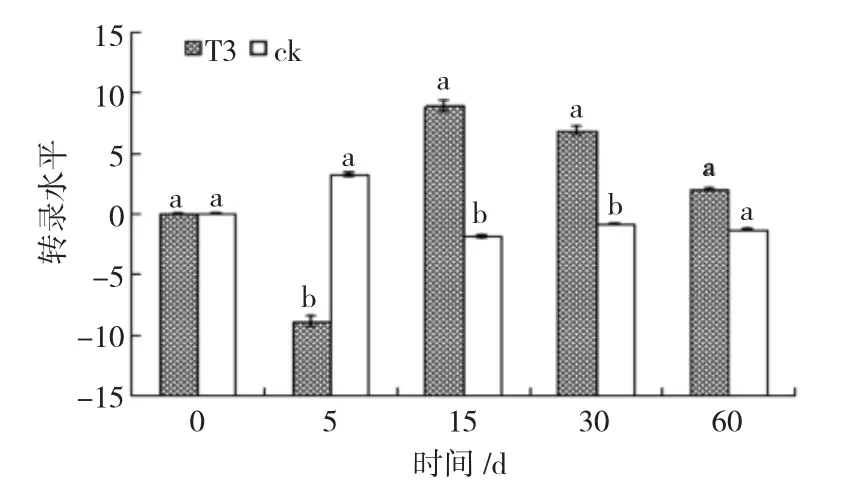
图4 内生木霉Th30 菌株诱导樟子松根系NPR1 基因的表达水平
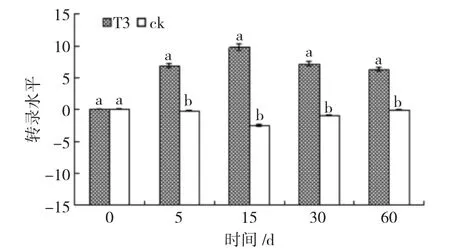
图5 内生木霉Th30 菌株诱导樟子松根系JAR1 基因的表达水平
3 结论与讨论
木霉菌被认为是一种可以广泛应用的新型促生剂[5],刘畅等[6]利用绿色木霉和哈茨木霉混合菌液灌根处理黄瓜幼苗后,发现混合菌液能够显著促进黄瓜幼苗生长,并且效果优于施加单种菌液。邓俊杰等[7]将哈茨木霉T6 施于月季盆栽土壤中,通过测定月季的相关生理指标表明哈茨木霉T6 具有促进月季生长和提高抗性的作用。李瑞敏等[8]也发现木霉菌对辣椒种苗的生物量、壮苗指数以及种苗移栽后株高、茎粗、单株果实重及根际木霉菌数量有显著影响。在本研究中,根施内生木霉Th30 菌株60 天后,T3(1×105个/mL Th30 孢子悬浮液)处理组的樟子松幼苗的株高、地茎、根长、鲜重和干重均显著高于对照(P<0.05),1 年生和2 年生植株均明显高于其他处理植株,因此,内生木霉Th30 菌株能够提高樟子松幼苗的生物量,并促进其生长。
抗逆性强的植株具有更高的蛋白质[9]和游离脯氨酸含量[10]。SOD 和PAL 等生理酶在植物正常生长发育和抵御病原菌侵害中也起着重要作用[10]。研究表明,棘孢木霉(T.asperellum)、深绿木霉(T.atroviride)、绿色木霉(T.viride)、长枝木霉(T.longibrachiatum)等均能通过提高植物的抗性生理指标来提高植物的抗病能力[10-13]。在本研究中,在内生木霉Th30处理下的樟子松幼苗中可溶性蛋白质含量、脯氨酸含量、SOD 和PAL 活性均显著升高,而T3 处理组的发病率最低,表明Th30 菌株对提高樟子松的抗病能力起到重要作用。
此外植物激素,特别是水杨酸(SA,salicylic acid)和茉莉酸(JA,Jasmonic acid),被广泛证实能够响应外界病原微生物的侵染[14]。在SA信号转导通路中,病程相关基因非表达子基因(NPR)在先天免疫和系统获得性抗性(System acquired resistance,SAR)中均发挥重要作用,NPR1 也是病程相关基因PR1(Pathogenesis-relatedproteingene)表达的关键成分,其过表达并未导致拟南芥(Arabidopsis thaliana)PR1的表达,从而提高其抗病能力[15]。JAR1 不仅影响JA 类化合物的合成,并且其作为一个信号中间体,参与茉莉酸信号途径的调控[16]。在本研究中,内生木霉Th30 定植樟子松幼苗过程中,根系中NPR1 基因和JAR1 基因在均在诱导初期显著上调表达,表明NPR1 基因和JAR1 基因能够参与Th30 菌株和樟子松幼苗的互作过程,从分子水平上再次证明内生木霉Th30 菌株能够提高樟子松的抗病能力。
综上所述,内生哈茨木霉Th30 菌株不仅能够促进樟子松幼苗的生长,还能通过提高樟子松幼苗蛋白质含量、脯氨酸含量、SOD 和PAL 活性、以及诱导樟子松幼苗根系中病程相关基因非表达子基因(NPR1)和茉莉酸抗性基因(JAR1)上调表达,从而提高樟子松幼苗的抗病能力。因此,在后续研究中对内生哈茨木霉Th30 菌株进行生物菌肥的开发和推广,对樟子松苗圃绿色发展具有重要意义。
猜你喜欢
今日农业(2022年4期)2022-06-01
广西植物(2021年9期)2021-10-16
翠苑(2019年4期)2019-11-11
发明与创新·大科技(2019年5期)2019-07-31
农民致富之友(2019年8期)2019-05-22
作文周刊·小学四年级版(2019年8期)2019-04-23
农民致富之友(2019年4期)2019-03-13
国际木业(2018年3期)2018-09-10
北极光(2018年12期)2018-03-07
现代农业研究(2017年8期)2017-09-22
