
毛果杨CNGC家族全基因组鉴定及胁迫响应分析
2022-08-04 01:59:58王雪莹王瑞琪夏德安魏志刚
植物研究 2022年4期
王雪莹 王瑞琪 张 洋 刘 聪 夏德安 魏志刚
(1. 林木遗传育种国家重点实验室,东北林业大学,哈尔滨 150040;2. 国家林业与草原局盐碱地研究中心,中国林业科学研究院,北京 100091)
环核苷酸门控离子通道(cyclic nucleotide-gat‑ed channels,CNGC)是非选择性阳离子通道,其C端的环核苷酸结合域(cyclic nucleotide binding do‑main,CNBD)可以与环磷酸腺苷(cAMP)或环磷酸鸟苷(cGMP)这两种信号分子结合,从而介导单价或双价阳离子的转运,故因此得名为CNGC。在植物界中,CNGC 可将钙离子、钾离子导入细胞质基质中,从而影响植物的生长发育及非生物胁迫响应。1998年,植物中的环核苷酸门控离子通道首次在大麦()中被确认。此后,家族在拟南芥(、烟草()、水稻()、玉米()、番茄()和 梨()等植物中陆续被发现。根据拟南芥家族系统发育关系,可将20 个家族成员分为4 个亚群(Ⅰ、Ⅱ、Ⅲ和Ⅳ),其中Ⅳ亚群包括Ⅳa亚组和Ⅳb亚组。植物CNGC的核心结构由6 个跨膜结构域(S1~S6)组成,其中S5 与S6 之间有1 个由20~30 个氨基酸组成的孔状结构(P-loop,P 环),其肽链的C 端具有环核苷酸结合域及钙调素结合域(calmodulin binding domain,CaMBD),并且两者有部分重叠,在植物家族中,CNBD 最保守的区域是磷酸盐结合域(phos‑phate binding cassette,PBC)和铰链区(hinge region,HR),它们是植物家族成员的重要标志。
研究发现,植物家族在植物的生长发育及应对多种生物与非生物胁迫响应中发挥着重要作用。例如,过表达使烟草的花粉管变宽、变短;水稻能够促进植株花粉管通道内的细胞程序性死亡从而促进花粉管的生长。此外,经研究发现拟南芥基因参与植物抗病;烟草基因可以抵抗烟草青枯病菌的侵染,这表明家族在促进植物免疫中也起到重要作用。此外,蓖麻()在盐胁迫处理下时,起到重要的信号传导作用。由此可见,家族广泛存在于植物中,参与植物的生长发育,对植物的逆境胁迫响应也有着重要作用,在植物的生命周期中不可或缺。
2006 年,毛果杨()被基因组测序,可作为木本研究的模式植物。目前有关毛果杨基因的信息较为缺乏。因此,在全基因组水平上对毛果杨家族进行鉴定与分类具有重要意义。本研究利用生物信息学手段在毛果杨中鉴定出19个家族成员并对其进行分析,包括其基因结构、蛋白基本理化性质、系统进化关系、蛋白保守基序分布以及分析了有关植物激素响应和非生物胁迫相关的顺式作用元件,并利用qRT-PCR 分析毛果杨在不同组织中家族的表达模式,目的是为研究该家族在渗透胁迫过程中的作用提供参考。为进一步了解毛果杨家族的进化关系及生物学功能打下基础。同时,为后续深入开展植物家族的渗透胁迫研究提供了参考依据。
1 材料与方法
1.1试验材料
本研究所用的毛果杨来自中国科学院分子植物科学卓越创新中心,利用植物组织培养技术将毛果杨进行扩繁,选取生长1 个月大小、且长势一致的毛果杨组培苗移栽到土壤中,在温室环境为(25±2)℃,长日照(光照16 h/黑暗8 h)下培养3 个月,分别采集毛果杨的根、茎和叶,液氮速冻并置于-80 ℃保存备用。此外,将温室中培养1 个月、大小且长势一致的毛果杨组培苗分别进行100 mmol•LNaCI和8%PEG-6000处理。每种胁迫分别处理6、12、24、48、72 h,同时用水处理作为对照组,每个时间点设置3 个生物学重复,分别采集各处理组植株材料的根、茎和叶,液氮速冻并置于-80 ℃保存备用。
1.2毛果杨CNGC家族的鉴定及理化性质分析
利用已报道的拟南芥家族的蛋白序列比对Phytozome 数据库(https://phytozome.jgi.doe.gov)中的毛果杨数据库,初步获得毛果杨家族的候选基因。将候选基因提交至Pfam(http://pfam.xfam.org/)、SMART(http://smart.embl-heidel‑berg.de/)、CDD(https://www.ncbi.nlm.nih.gov/Struc‑ture/cdd/wrpsb. cgi)、PROSITE(https://prosite. ex‑pasy.org/)在线蛋白数据库分析其结构域,以含有CNBD 结构域及同时含有离子转运蛋白(Ion trans‑porter,ITP)结构域或跨膜结构域(Transmembrance domain,TM)为条件进行筛选,同时利用BioEdit 软件进行多序列比对,以具有CNBD 结构域内的PBC 和HR 为标准,最终鉴定出毛果杨家族的全部成员。
利用Phytozome 数据库获得家族成员的染色体位置和基因序列等信息,根据基因所在的染色体号及位置对C 家族成员进行命名;利用在线软件ExPASy(https://web.expasy.org/protparam/)预测PtrCNGC蛋白理化性质,包括氨基酸数、蛋白分子量和等电点等;使用TBtools软件对染色体位置信息进行可视化处理;通过YLoc 网站(https://abi-services.informatik.uni-tuebingen.de/yloc/webloc.cgi)在线预测家族成员的亚细胞定位信息。
1.3毛果杨CNGC家族系统发育分析
使用MEGA 6.0软件中的ClustaW 程序将鉴定出的毛果杨家族成员的蛋白序列与拟南芥、水稻和梨的家族成员的蛋白序列进行多重比对。采用最大似然法(Maximum likelihood,ML)构建系统进化树,Bootstrap 参数设置为1 000。将得到的系统进化树数据经iTOL 网站(https://itol.embl.de/)进行可视化处理。
1.4毛果杨CNGC 家族同源基因的Ka/Ks比值分析
利用NCBI中的BLAST在线网站(https://blast.ncbi.nlm.nih.gov/Blast.cgi),比对毛果杨家族成员之间的CDS 序列,其序列的相似性超过80%则认为具有同源性。利用TBtools 软件计算同源基因之间的同义替换率(synonymous substitu‑tions rates,)、非同义替换率(nonsynonymous sub‑stitution rates,)以及/比值。当/>1,认为有正选择效应;/=1,认为存在中性选择;/<1,则认为有纯化选择作用。
1.5毛果杨CNGC 家族基因结构及蛋白保守基序分析
利用TBtools软件将的外显子—内含子信息进行可视化。基因保守基序(Motif)由在线软件MEME(http://meme-suite.org/tools/meme)分析获得,使用默认参数。并利用在线网站WebLogo(http://weblogo.threeplusone.com/create.cgi)对Motif进行可视化处理。最后利用BioEdit 软件对家族的蛋白序列进行多序列比对,对其保守结构域进行分析。
1.6毛果杨CNGC 基因家族启动子区顺式作用元件分析
在Phytozome 网站上下载家族基因起始密码子上游2 000 bp 的序列作为启动子序列信息,利用在线网站PlantCare(http://bioinformat‑ics.psb.ugent.be/webtools/plantcare/html)进行顺式作用元件在线预测分析,最后利用TBtools 软件对其位置进行可视化。
1.7毛果杨CNGC 基因家族组织表达特异性及盐胁迫和PEG胁迫响应特异性分析
使用植物总RNA 提取试剂盒(MiniBEST,Ta‑KaRa)提取总RNA,然后利用PrimeScriptRT re‑agent Kit(Perfect Real Time,TaKaRa)试剂盒反转录获得cDNA 用于qRT-PCR。根据荧光定量引物设计原则,设计家族基因定量引物,以为内参基因(见表1)。使用赛默飞ABI 7500实时荧光定量PCR仪进行试验,体系如下:2×TransStart TOP/Tip Green qPCR Supermix 10.0 μL、上下游混合引物(10 μmol·L)0.4 μL、cDNA 1.5 μL,Passive Reference Dye(50×)0.4 μL,加ddHO至20.0 μL。反应条件如下:94 ℃30 s;94 ℃5 s,60 ℃15 s,72 ℃35 s,循环40 次;95 ℃15 s,60 ℃1 min,95 ℃30 s。每组处理重复3 次,最后采用2法计算相对表达量并利用TBtools软件进行可视化分析。

表1 毛果杨CNGC家族基因qRTPCR引物Table 1 Primers for qRT-PCR of PtrCNGC gene family
2 结果与分析
2.1毛果杨CNGC家族成员的基因鉴定
CNBD 是植物家族的典型结构域,尤其是存在于CNBD 区域内的PBC 和HR。为了确认CNBD 结 构 域,使 用5 个 蛋 白 数 据 库(Pfam、SMART、CDD、PROSITE和SurperFamily)对家族的结构域进行分析(见表2)。同时使用多序列对比方法检查各序列CNBD 中的PBC 和HR(见图3)。最终在毛果杨中鉴定出19 个。家族除了具有主要的结构域外,其中、和还具有含IQ 基序(IQxxxRGxxxR,Pfam00612)的钙调素蛋白结构域。
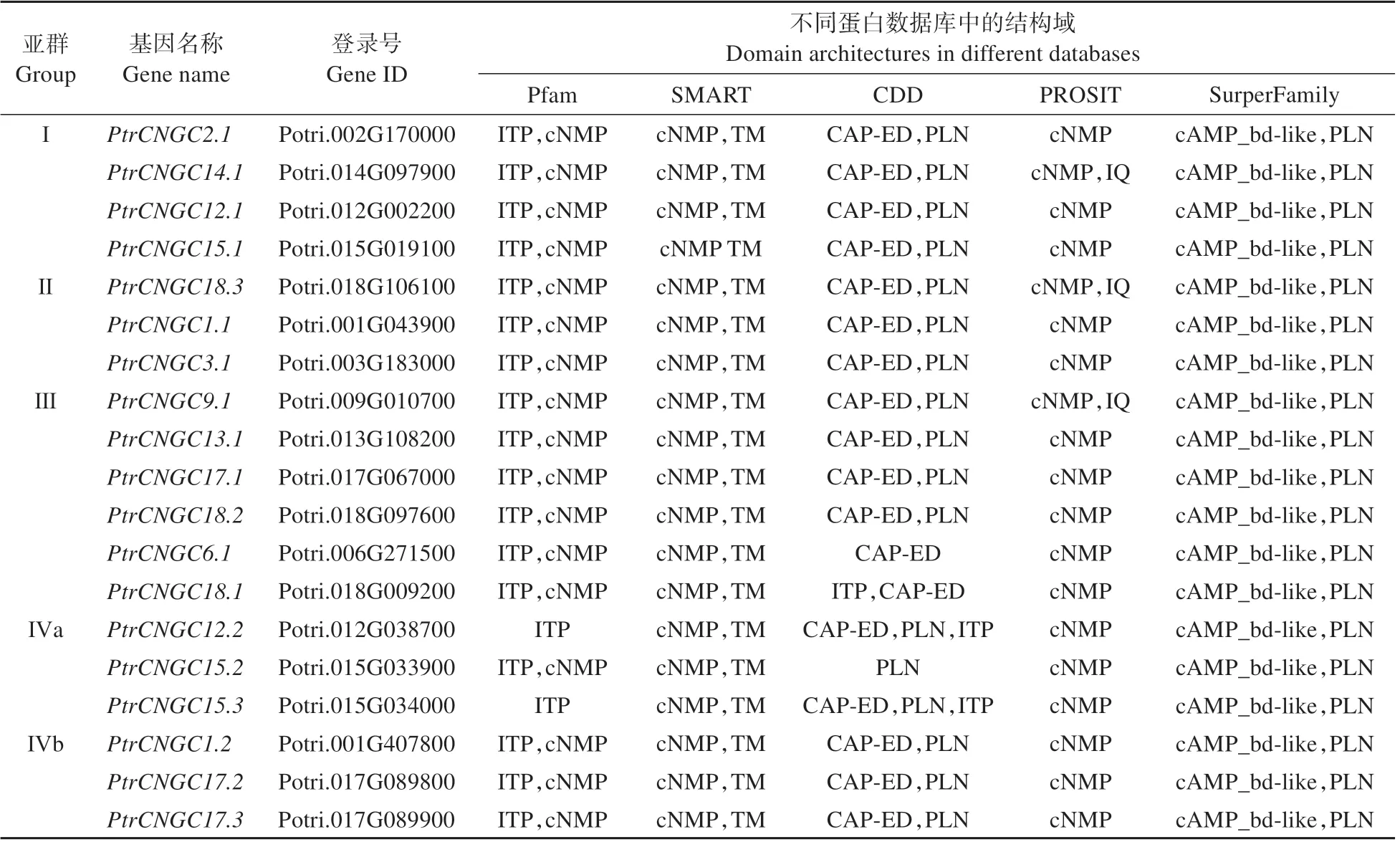
表2 毛果杨CNGC基因家族结构域Table 2 Domain architecture of PtrCNGC gene family
2.2毛果杨CNGC家族成员及其理化性质分析
经鉴定在毛果杨基因组中共筛选出19 个家族成员,并根据家族成员所在的染色体号及位置对其进行命名(见表3)。19 个的基本特征如表3 所示:家族的蛋白长度为684~785 个氨基酸,这些蛋白的分子 质 量 在78.532 21~89.681 54 kDa,pI 为7.53~9.55,19 个家族成员全部为碱性蛋白,此外,除PtrCNGC17.3为疏水性蛋白,其余成员均为亲水性蛋白。亚细胞定位预测分析显示,除PtrCNGC17.1分布在过氧化物酶体上,其余PtrCNGC 蛋白均分布在细胞质中。
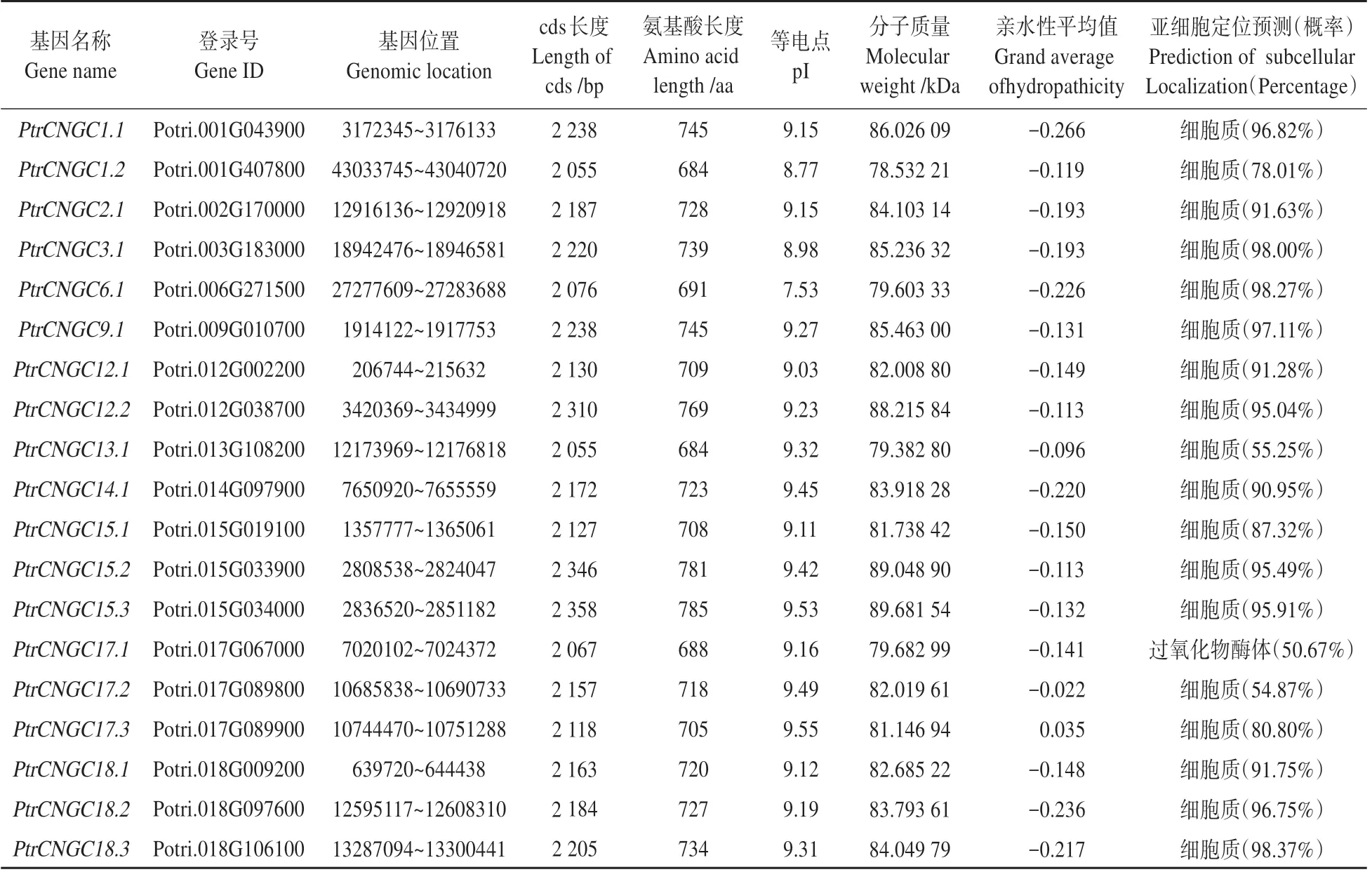
表3 毛果杨CNGC 家族概况Table 3 Summary of PtrCNGC gene family
2.3毛果杨CNGC家族成员的系统进化分析
将鉴定得到的19 个毛果杨PtrCNGCs 的氨基酸序列与20 个拟南芥AtCNGCs、16 个水稻Os‑CNGCs,以及21 个梨PbrCNGCs的氨基酸序列进行比对,采用ML 法共同构建系统进化树。如图1 参考拟南芥家族的聚类结果,可将毛果杨家族分成4 个亚群Ⅰ、Ⅱ、Ⅲ和Ⅳ(Ⅳa、Ⅳb)。系统进化树结果显示,毛果杨、拟南芥、水稻和梨在5 组(Ⅰ、Ⅱ、Ⅲ、Ⅳa 和Ⅳb)中分别含有20、13、21、9和13个成员,其中毛果杨家族中的亚群Ⅰ含有4 个成员;亚群Ⅱ含有3 个成员;亚群Ⅲ含有的成员数最多为6 个;亚组Ⅳa 和Ⅳb 都各含有3 个成员。根据系统发育关系,发现在家族亚群Ⅲ含有的成员最多,亚组Ⅳa含有的成员最少。进化树分析表明,毛果杨与拟南芥和梨的亲缘关系较近,与水稻的亲缘关系较远,这或许与水稻为单子叶植物而毛果杨与拟南芥和梨共为双子叶植物有关。
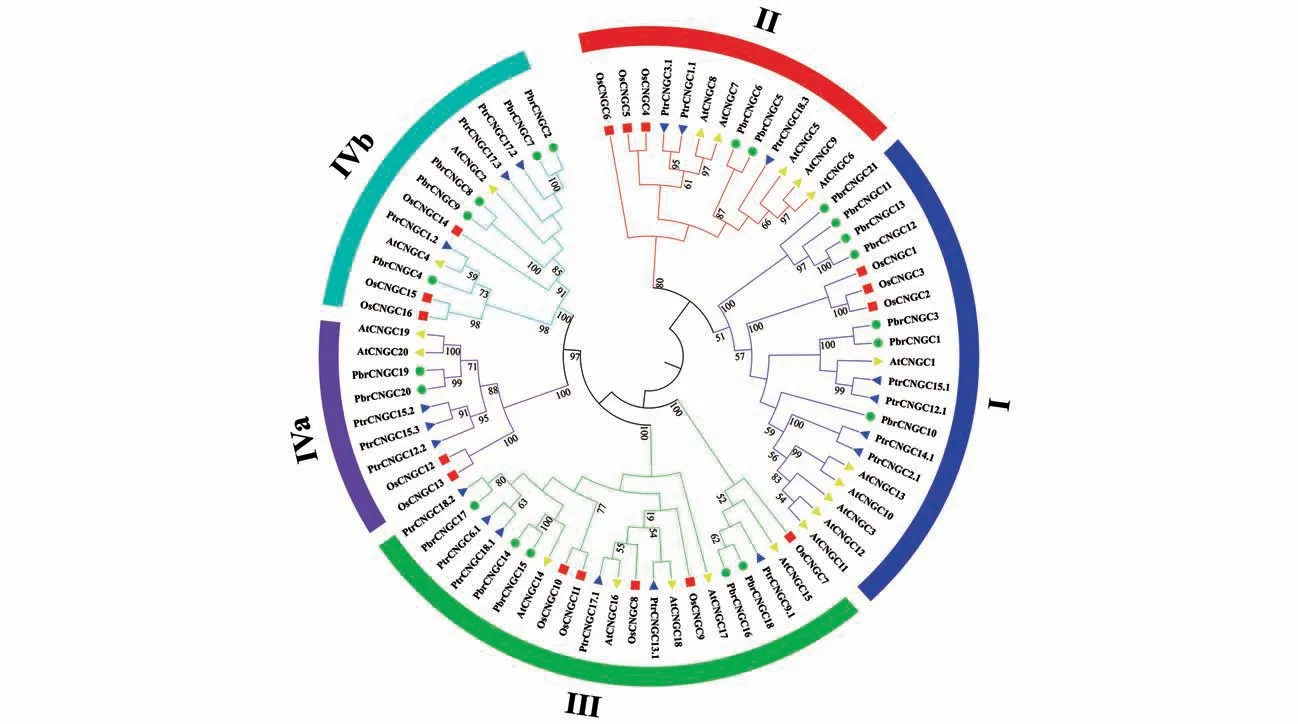
图1 毛果杨与拟南芥、水稻和梨的CNGC家族的系统发育进化树蓝色三角、黄色三角、红色方形和绿色圆形分别代表毛果杨、拟南芥、水稻和梨,分支上的数值代表Bootstrap 1 000 个循环的置信度(只显示50%以上的置信度)Fig.1 Phylogentic Tree of CNGC gene family from Populus trichocarpa(Ptr),Arabidopsis thaliana(At),Oryza sativa(Os)and Pyrus bretchneider(iPbr)Blue triangle,yellow triangle,red square and green circle represent Populus trichocarpa,Arabidopsis thaliana,Oryza sativa and Pyrus bretchneideri,the numbers represent the percentage of 1 000 Bootstrap replication(sonly above 50%showed)
2.4毛果杨CNGC 家族成员染色体定位以及同源基因的Ka/Ks比值分析
家族成员染色体分布情况如图2 所示,2号、3号、6号、9号、13号和14号染色体上有1个基因,1号和12号染色体上有2个基因,15号、17号和18 号染色体上各有3 个基因,发现19 个家族成员在染色体上呈不均匀分布,有8个染色体上没有该家族成员分布。
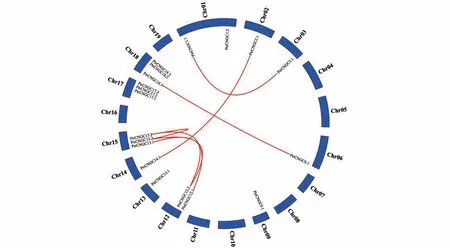
图2 PtrCNGC家族基因染色体定位及同源关系红色线连接有同源关系的同源基因Fig.2 Chromosome distribution of PtrCNGC gene familyThe line connects the homologous genes
利 用BLAST(https://blast.ncbi.nlm.nih.gov/Blast. cgi? PROGRAM=blastn & PAGE_TYPE =BlastSearch&LINK_LOC=blasthome)对家族成员的核苷酸序列进行在线分析,结果显示(见表4),和、和、和、和、和以及、和之间的序列相似性超过80%,表明具有同源关系,且上述各具基因对是由于基因复制事件进化形成旁系同源基因。其中和共 在15 号 染 色 体上,且物理距离小于200 kb,表明发生了串联重复。
为了阐明选择压力在家族进化中的作用,利用TBtools 软件分析了家族同源基因的值、值和/值。结果表明(见表4),家族7 对同源基因的/值均远小于1,表明家族在进化过程中经历了较强的纯化选择。
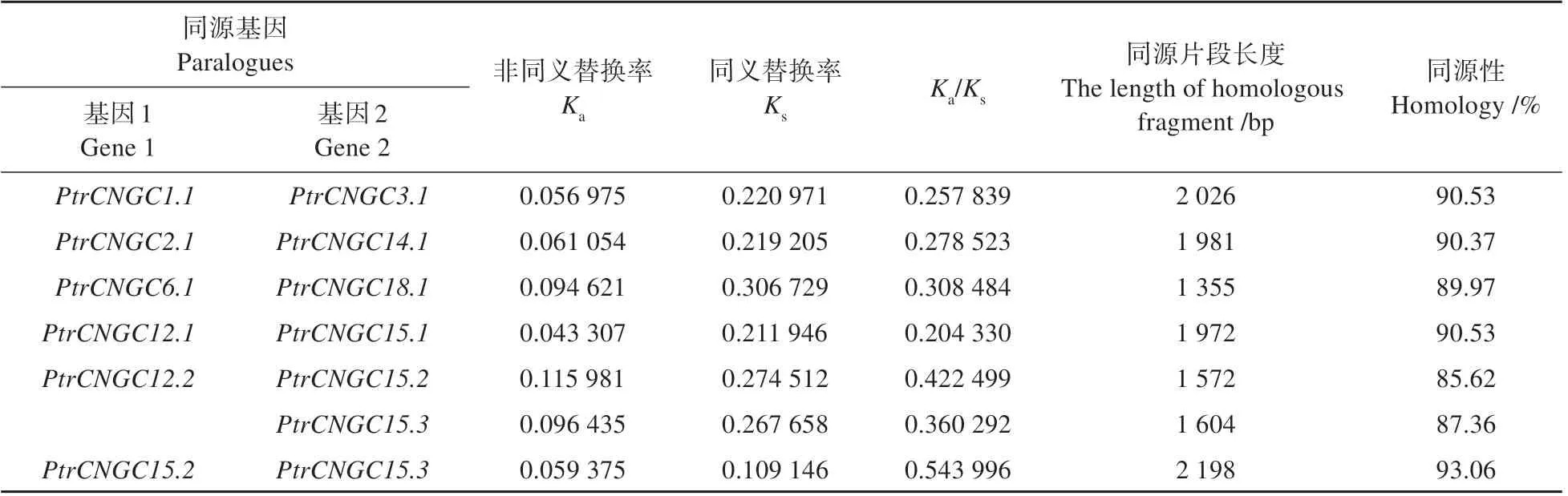
表4 同源基因的Ka/Ks比值及同源性Table 4 Ka/Ks ratio and homology of homologous genes
2.5毛果杨CNGC 家族基因结构及保守基序的分析
CNBD 是植物CNGC 中典型的结构域,如图3所示其保守基序为L-X2-G-[DNG]-[FV]-X-G-[ED]-L-[LF]-X-W-X-L-X(10,20)-S-X(2)-T-X(7)-E-A-F-X-L,该基序与其他植物基本吻合。如图3 所示谷氨酸(E)、丙氨酸(A)、苯丙氨酸(F)和亮氨酸(L)在HR 中是保守的(100%),亮氨酸(L)、甘氨酸(G)和色氨酸(W)在PBC 中是保守的(100%),与其他植物相比,家族中HR 的保守性无明显差异,但PBC 较其他植物表现得更为保守。
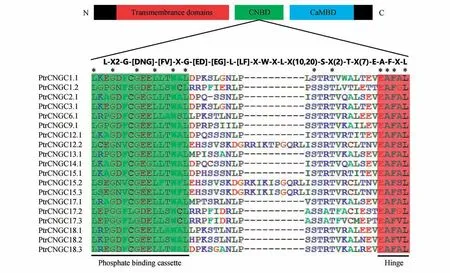
图3 PtrCNGC蛋白CNBD中PBC和HR顶部图形代表了PtrCNGC 蛋白的3 个区域:TM、CNBD 和CaMBD;星号表示19 个PtrCNGC 成员之间氨基酸100%保守,绿色和红色区域分别代表PBC和HRFig.3 The PBC and the HR within the CNBD of PtrCNGC proteinsThe diagram at the top represents three regions of PtrCNGC proteins:TM,CNBD and CaMBD;The asterisk indicate 100% identity among the 19 PtrCNGCs;green and red regions represent the PBC and HR
分析了19个家族成员的外显子—内含子的结构分布(见图4),结果显示,家族中Ⅰ、Ⅱ、Ⅲ亚群以及Ⅳa 亚组中成员外显子的数量为6~8 个,而Ⅲ亚群中的的外显子数量最少仅有2 个,Ⅳb 亚组中外显子的数量最多且均为12 个,发现位于同一分支上的家族成员的外显子数量接近。此外、和的外显子—内含子结构分布与其他成员存在较大差异,表明在进化过程中可能存在基因结构变异。

图4 PtrCNGC家族基因的外显子—内含子结构分布Fig.4 Exon-intron structure of PtrCNGC gene family
为进一步研究毛果杨家族的特征,使用MEME 对其蛋白保守基序进行了分析(见图5)。结果显示,家族编码蛋白的10 个保守基序(Motif 1~10)中,同一亚群及同一亚组的成员保守基序的位置分布相似。19个成员具有9 个共同的基序,表明这些基序在家族中具有重要作用。此外缺失了基序4,而具有2 个基序4,说明家族基因编码蛋白结构在具有保守性的同时,功能上也存在一定差异。
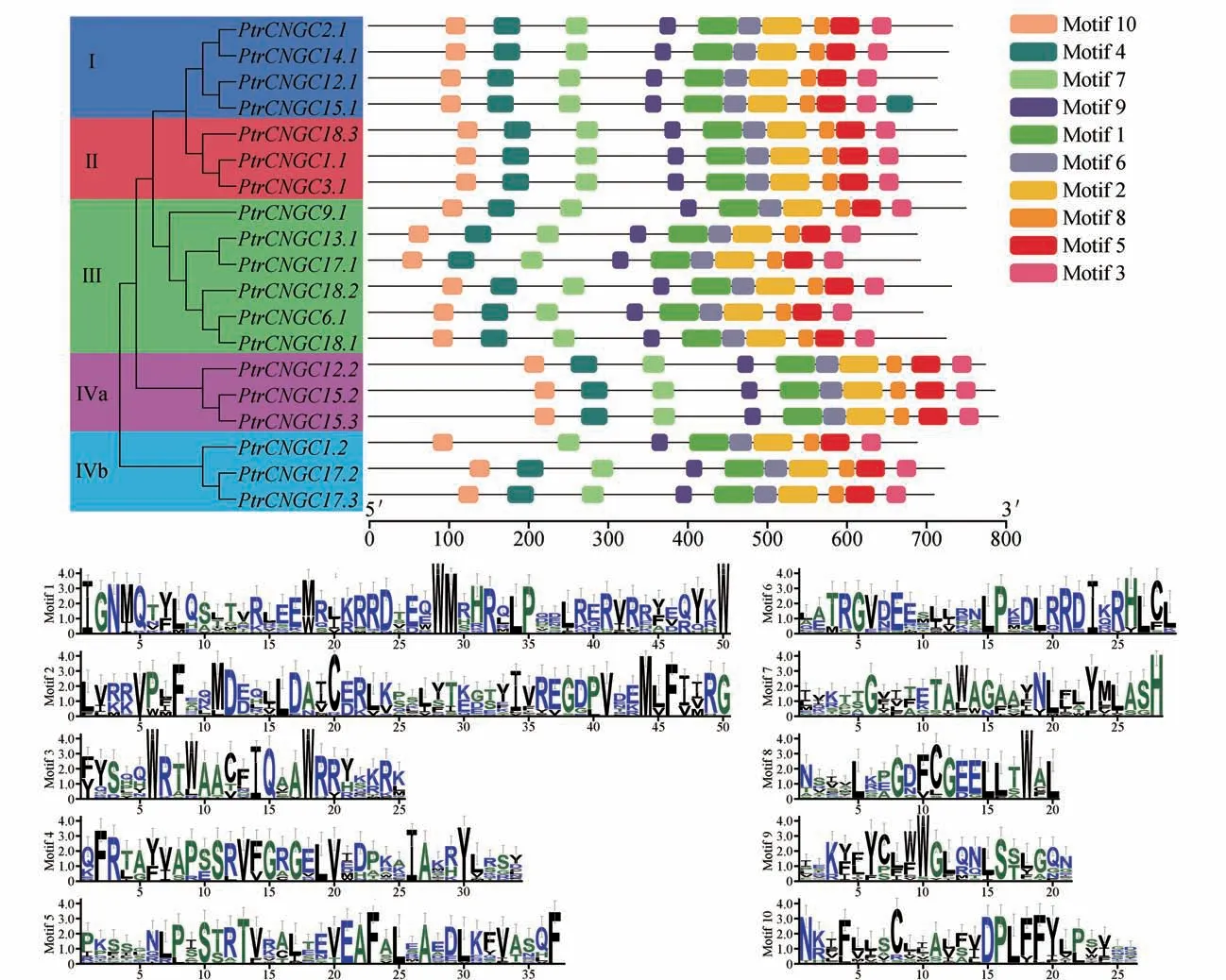
图5 PtrCNGC家族蛋白保守基序分析Motif 1~10用不同的颜色表示;Motif 1~10序列展示在下方Fig 5 The protein conserved motif of PtrCNGC gene familyMotif 1-10 are indicated by different colors;The sequences of the motifs 1-10 are shown below
2.6毛果杨CNGC家族启动子特征分析
通过PlantCARE 数据库分析,在的启动子区域内发现了大量的顺式作用元件,其中包括多种响应非生物胁迫及植物激素的顺式作用元件。结果(见图6)表明,在毛果杨家族中发现的响应非生物胁迫的元件有厌氧诱导元件(ARE)、干旱响应元件(MBS)、光响应元件(Gbox)、低温响应元件(LTR),以及响应光、盐、低温的元件(AE-box);响应植物激素的元件包括水杨酸响应元件(TCA-element)、脱落酸响应元件(ABRE)、乙烯响应元件(ERE)、茉莉酸甲酯响应元件(CGTCA-motif)、赤霉素响应元件(P-box),以及生长素响应元件(TGA-element)。这些结果说明家族成员极有可能被盐、干旱、低温和生长素等非生物胁迫诱导表达,表明毛果杨家族在非生物胁迫以及激素响应中可能具有重要作用。

图6 PtrCNGC族启动子区顺式作用元件Fig.6 Analysis of cis-acting elements in the promoter region of PtrCNGC gene family
2.7毛果杨CNGC 家族在各组织中的表达模式分析
通过qRT-PCR 对毛果杨基因家族成员在根、茎和叶中的表达情况进行了分析。结果(见图7)表明,在家族中,有13 个成员在茎中表达量最高,4 个成员在叶中表达量最高,而仅有2个成员在根中表达量最高,表明家族的多数成员在毛果杨茎中表达量较高,而在根和叶中表达量较低。此外,发现同一分支的多数成员在不同组织中的表达模式相似。
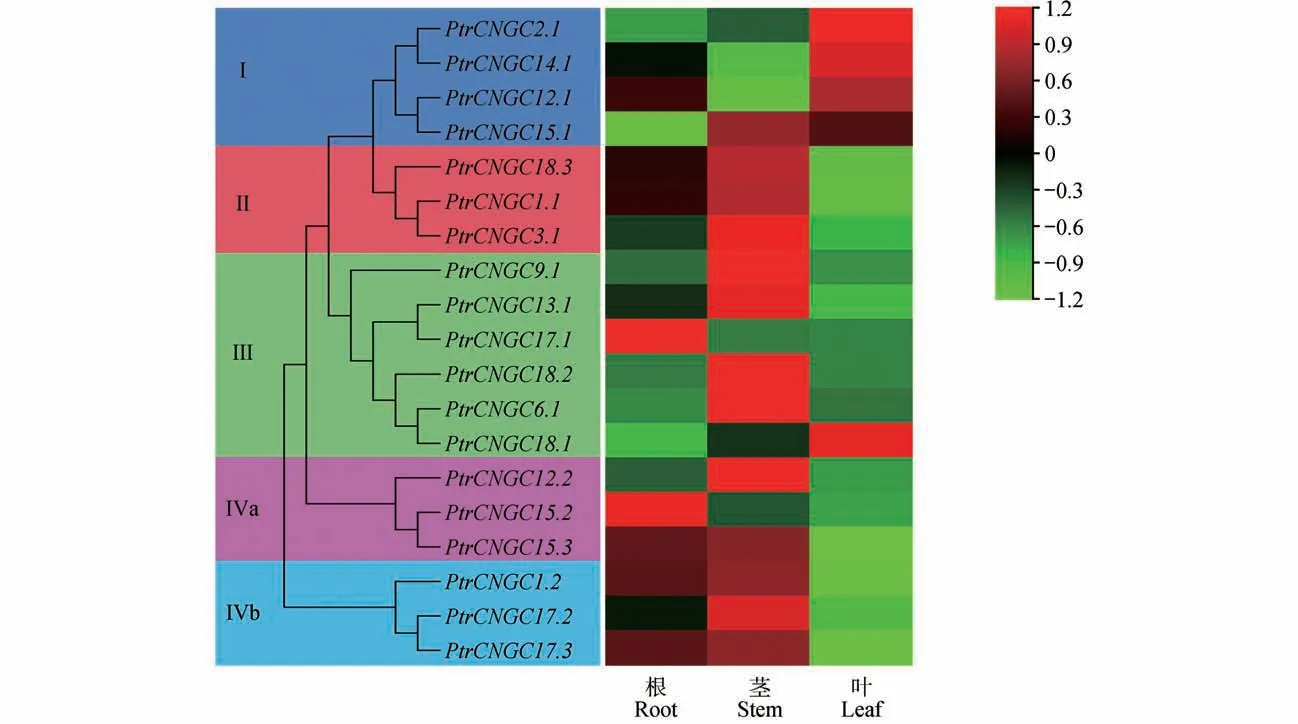
图7 PtrCNGC基因组织表达特异性分析Fig.7 Tissue-specific expression analysis of PtrCNGC gene family
2.8毛果杨CNGC 家族在盐胁迫下的表达模式分析
进一步了解家族的非生物胁迫响应特性,对盐胁迫下的家族成员的表达模式进行了分析。如图8 所示,在根中,多数家族成员在24 或48 h 表达量最高,而Ⅳb 亚组成员随时间的增加表达量呈下降趋势且在0 h 时表达量最高。在茎中,多数家族成员在胁迫处理6 h 的表达量最高,在72 h 时表达量最低。在叶中,Ⅰ、Ⅱ、Ⅲ亚群以及Ⅳa亚组中的多数成员在胁迫处理24 或48 h 时表达量最高,而Ⅳb 亚组中的在0 h时表达量最高、和分别在12 和24 h 表达量最高。说明家族成员在植物不同组织中对盐胁迫的响应程度不同,可能在毛果杨响应盐胁迫过程中发挥了重要作用。
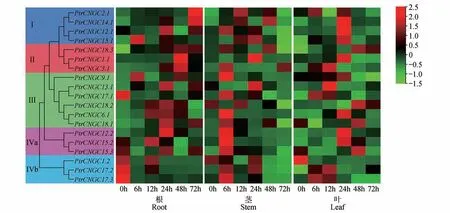
图8 盐处理下PtrCNGC基因的表达水平分析Fig.8 Analysis of expression level of PtrCNGC gene family in salt treatment
2.9毛果杨CNGC 家族在PEG 胁迫下的表达模式分析
利用PEG模拟干旱胁迫下家族成员的表达情况如图9所示,在根中,Ⅱ亚群全部成员与Ⅲ亚群中的、、及Ⅳa 亚组中的在0 h时表达量最高;Ⅳb 亚组中的全部成员与亚群Ⅰ中的和、Ⅳa 亚 组 中和、Ⅳb 亚组中在胁迫处理24 h 时表达量最高。在茎中,多数家族成员在胁迫处理0 或6 h 时表达量达到峰值,而Ⅳb 亚群中的所有成员均在胁迫处理48 h 时表达量达到峰值。在叶中,多数r家族成员在胁迫处理48 或72 h 时表达量最高,且随着时间的增加,表达量呈先下降后上升的趋势。结果表明19个成员均在PEG 处理后有不同程度的响应模式,发现同一分支上的多数成员表现出相似的表达模式。
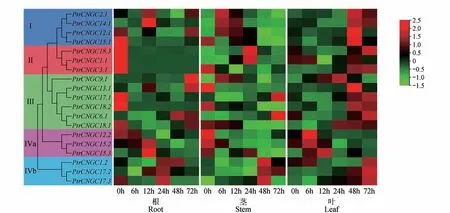
图9 PEG处理下PtrCNGC基因的表达水平Fig.9 Analysis of expression level of PtrCNGC gene family in PEG treatment
3 讨论
家族基因广泛存在于植物中,参与调控植物的生长发育及胁迫响应过程。本研究从毛果杨全基因组中共鉴定出19 个家族成员,根据系统发育关系将毛果杨家族分为5 组(Ⅰ、Ⅱ、Ⅲ、Ⅳa和Ⅳb),其中Ⅰ、Ⅱ、Ⅲ亚群之间的亲缘关系比较近,而第Ⅳ亚群与其他亚群的关系较远且2个亚组成员之间的距离也相对较远,这与此前拟南芥家族的研究结果一致。亚细胞定位预测结果显示,毛果杨CNGC 蛋白主要定位在细胞质中。而在水稻家族中,亚细胞定位预测结果发现其多数成员分布在细胞质膜上,且在细胞质、叶绿体类囊体膜和线粒体内膜上也有分布。此外,现有研究结果表明多数拟南芥家族成员蛋白定位于细胞膜质上,这与植物CNGC具有转运阳离子的能力以及含有TM、ITP等功能结构域有关。
此外,同一分支家族成员的蛋白保守基序分布及其数量基本一致,推测同一亚群成员的生物学功能具有相似性。在家族发现7 对旁系同源基因,表明基因复制事件在家族扩张中起主要作用,基因复制可以增加基因的数量以及功能的多样性。同时7 对同源基因的/比值均远小于1,说明这些相关基因是在较强的纯化选择下进化而来的,根据已有研究发现拟南芥和梨的家族经历了较强的正向选择压力,说明植物家族进化方式存在差异。此外,Ⅳa 亚组全部的3 个成员互为旁系同源基因,说明该亚组在进化过程中较为保守,其功能可能具有高度相似性。
毛果杨家族中Ⅳb亚组成员的外显子数量最多,它们提高了发生选择性剪接的可能性,从而增加了家族功能的多样性。通过研究毛果杨家族在根、茎和叶中的表达情况,发现其在茎中的表达量较高,而在根和叶中的表达量较低,说明家族在不同组织中的表达具有特异性。此外,发现亚群Ⅱ中的在根中的表达量较高,茎和叶中的表达量较低,而亚群Ⅱ的其余两个成员(和)在根和叶中表达量较高,茎中的表达量较低,同时根据家族基因结构分析结果,发现的内含子明显长于和,因此推测此亚群成员在植物不同组织的表达情况或许与其基因结构有关。
根据顺式作用元件分析,家族成员含有数量不等的与响应植物激素和非生物胁迫过程有关的顺式作用调控元件,如干旱响应元件(MBS),响应光、盐、低温的元件(AE-box)等,推测它们在植物的生长发育和响应胁迫过程中起着重要作用。根据所研究的19 个在盐胁迫与干旱胁迫下的表达情况,发现家族成员在相同胁迫处理下,同一分支上的成员表达模式相似,如在盐胁迫处理下,在根中的Ⅱ亚群所有家族成员表达模式呈先上升后下降的趋势,且均在胁迫处理48 h 时表达量最高。而在不同胁迫处理下,家族成员的表达模式不同,如亚群Ⅱ中的成员在根中的表达量随处理时间的增加而上升,达到峰值后开始下降,但在干旱胁迫处理下,其表达量明显下降,表明家族在不同胁迫处理下的响应能力不同。本研究为进一步研究毛果杨家族响应非生物胁的功能提供参考。
猜你喜欢
新医学(2023年10期)2023-12-09 15:04:51
故事作文·高年级(2023年3期)2023-05-30 19:00:18
现代临床医学(2023年1期)2023-03-24 08:30:06
中国医药科学(2022年5期)2022-05-05 23:58:07
南方医科大学学报(2022年3期)2022-04-13 01:51:26
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10 07:07:36
中国卫生标准管理(2015年3期)2016-01-14 03:41:45
分析测试学报(2015年5期)2016-01-13 06:18:50
安徽医药(2014年4期)2014-03-20 13:13:40
中成药(2014年11期)2014-02-28 22:30:08
