
木薯SR45亚家族基因鉴定及表达
2022-08-02 07:14:12李智博董世满李淑霞曾长英
华南农业大学学报 2022年5期
李智博,董世满,李淑霞,彭 明,曾长英
(1 海南大学 热带作物学院, 海南 海口 570228; 2 中国热带农业科学院 热带生物技术研究所, 海南 海口 571101;3 中国热带农业科学院 海南热带农业资源研究院, 海南 海口 571101)
木薯Manihot esculenta是一种热带作物,原产自非洲、南美洲热带亚热带地区,木薯淀粉储存根是重要的粮食、动物饲料和工业原料。木薯在全球粮食安全和经济发展中发挥着重要作用[1],然而,木薯被归为一种对低温敏感的物种,所以木薯种植区域大多数在我国年平均气温较高的海南、广西等地区[2]。低温和冻害是影响植物发育、限制作物地理分布和产量提高的重要环境因素之一[3]。研究木薯在低温逆境中的适应机制,提高其抗逆性,对于提高木薯产量具有重要意义。
在植物中,可变剪接(Alternative splicing)是一种重要的转录后调控机制,通过选择不同的剪切位点产生具有改变稳定性、定位或翻译效率功能的不同mRNA转录物来调节基因表达[3]。目前可变剪接已被证明在植物对非生物胁迫的反应中发挥作用,包括热、干旱和冷胁迫[4-7]。精氨酸和丝氨酸富集蛋白 (Arginine/serine-rich proteins,SR)基因在冷和热胁迫下剪接模式的变化表明剪接变异体在应激反应中的比例改变可能在植物对这些应激的适应中发挥了作用[8]。Li等[9]通过对木薯转录组测序的大规模分析揭示了可变剪接异构体的数量和丰富度受到低温应激的特异性调控。Leviatan等[3]通过平铺阵列揭示低温下新型冷应答基因,基因表达水平未发生改变,但是可变剪接模式的相对丰度却显示出变化。
SR蛋白是一种非小核核糖核蛋白(Non-small nuclear ribonucleoprotein,Non-snRNP)剪接体蛋白家族,是前体mRNA组成型剪接和可变剪接中保守且关键的调控因子[10-14]。SR蛋白形成1个保守的RNA结合蛋白家族,具有独特的结构域结构,是由1个或2个RNA识别基序(RRM)和C端富含精氨酸/丝氨酸(RS)的1个特征性结构域组成,而SR-like(SR45)蛋白中RS结构域的分布位置与SR基因家族其他成员不同,其RS结构域位于N端[15-16]。植物SR蛋白可分为6个亚家族:SR、RS、SCL、RSZ、RS2Z和SC35,不包含SR-like亚类,但具有类似SR蛋白特征且与SR蛋白系统发育关系密切的蛋白质被分类为SR-like蛋白[15,17]。在本文中,为了方便起见,我们用SR这一名称统称7大亚类(包含SR-like亚类)。在拟南芥中鉴定出18种SR基因家族成员以及SR-like(AtSR45和AtSR45a)。在木薯中,前人报道SR基因家族有18个成员,包含SR、RS、SCL、RSZ、RS2Z、SC35共 6大亚类[18-19],缺少 SR-like亚类,为此,本研究在前人工作的基础上对木薯SR基因家族SR-like亚类进行补充鉴定。值得注意的是,SR家族在细胞对各种非生物胁迫的适应过程中发挥着重要作用[13,20-21]。例如,在拟南芥中,SR剪接因子基因的表达和剪接模式受到低温的差异调节[11],AtSR45基因的缺失导致盐胁迫敏感性增加,调控盐胁迫的基因表达模式与剪接模式发生改变[22]。
木薯是热带亚热带地区优质的粮食作物,低温与冻害成为限制扩大其种植面积的重要因素,因此提高木薯的抗低温能力和选育耐低温的木薯品种是减少木薯在低温逆境中受损的有效方法之一,近年来对木薯抗逆基因的研究具有一定的进展,但是SR蛋白尤其SR45蛋白抗低温逆境的研究相对较少。本研究通过转录组测序分析筛选出木薯SR45相关基因,并运用生物信息学的方法对其系统进化、理化性质以及结构特征进行深入分析,以及对SR-like亚类进行了补充;同时结合荧光定量PCR(RT-qPCR)分析MeSR45在木薯组织中的时空表达模式以及在低温胁迫下的表达模式,以期进一步了解木薯SR-like亚类中SR45基因的生物学功能以及在低温胁迫中的响应,同时为其他物种SR基因家族的生物信息学研究提供参考。
1 材料与方法
1.1 试验材料与处理
研究材料为‘木薯60444’组培苗,该品种由中国热带农业科学院热带生物技术研究所功能基因组研究实验室提供。选取高度、长势大致相同的组培苗,平均分成2份,分别用于木薯低温胁迫试验、基因时空表达检测试验与干旱胁迫试验。在低温胁迫试验中,组培苗在4 ℃低温处理24 h,并以(24±2) ℃室温处理为对照,取组培苗茎顶端组织(包含芽尖以及2片完全展开的功能叶);时空表达试验材料取组培苗根、茎、叶和芽4部分组织;在干旱胁迫试验中,采用20%(w)PEG6000处理木薯组培苗6 h。以上试验所取组织均用于RNA抽提,本试验中每个试验样品均取3次重复,试验结果以平均值呈现。
1.2 基因家族成员的序列
从JGI植物基因组数据库(https://phytozome.jgi.doe.gov/pz/portal.html)获得木薯SR基因ID、编码区(CDS)长度和氨基酸长度,使用序列处理在线工具包(http://www.bio-soft.net/sms/prot_mw.html)计算蛋白质相对分子质量,使用Expasy网站(https://web.expasy.org/compute_pi/)计算木薯SR45相关蛋白成员理论等电点,亚细胞定位预测网址为http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/。
1.3 基因家族分析、氨基酸序列比对和结构特征分析
从JGI植物基因组数据库(https://phytozome.jgi.doe.gov/pz/portal.html)下载拟南芥、木薯的SR蛋白序列。利用DNAMAN6.0软件对木薯SR45蛋白序列进行多序列比对。利用MEGA11对2个物种46条氨基酸序列构建系统进化树(Neighborjoining法和1 000次迭代bootstrap测试)。利用TB-tools软件对木薯SR-like蛋白结构以及Motif基序进行绘制。
1.4 木薯总RNA提取
称取木薯组织样品1.0 g,用液氮冰冻后充分研磨,按照RNAprep Pure多糖多酚植物总RNA提取试剂盒(天根生化科技有限公司)的说明书,提取木薯总RNA,使用超微量分光光度计NanoDrop 2000检测总RNA的浓度和纯度。参照FastKing Gdna Dispelling RT SuperMix 反转录试剂盒 (TaKaRa公司)的说明书进行反转录,RNA抽提后存放于-80 ℃冰箱。
1.5 RT-qPCR分析
以木薯内参基因MeActin为参照,使用RT-qPCR分析基因的相对表达量,引物序列见表1。RT-qPCR所用试剂为TaKaRa公司TB Green®Premix Ex Taq™ II(Tli RNaseH Plus),使用的仪器为 Applied Biosystems StepOneTMand StepOnePlusTMReal-Time PCR Systems,采用 2-ΔΔCt法计算分析基因相对表达量。本试验RT-qPCR基因特异性引物均利用NCBI网站设计,引物合成由北京擎科生物科技有限公司海口合成部(hk-synth@tsingke.net)合成。
表1 MeSR45基因RT-qPCR引物序列Table 1 Primer sequences of MeSR45 genes for RT-qPCR
1.6 转录组数据
RNA-seq数据提交至NCBI的Sequence Read Archive (SRA),登录号为 SRP101302。
2 结果与分析
2.1 木薯SR45蛋白理化性质与基因序列分析
本研究对木薯转录组数据进行分析,筛选出与木薯可变剪接事件相关的SR45蛋白,分别获得5个木薯SR45同源基因,参考拟南芥同源基因命名方式依次命名为MeSR45-1~MeSR45-5,基因号依次为 Manes.14G016200、Manes.06G171800、Manes.15G168400、Manes.07G135700、Manes.S056000。MeSR45-1~MeSR45-4基因分别分布在木薯14、6、15、7号染色体上,MeSR45-5的染色体定位未知;蛋白长度为135~423 aa。推测MeSR45蛋白相对分子质量介于 14 970~47 210;MeSR45-1~MeSR45-4 等电点(PI)介于 10.97~12.34,为碱性蛋白,而MeSR45-5蛋白的等电点为5.19,为酸性蛋白。利用亚细胞定位预测网站对MeSR45基因家族蛋白进行预测,结果表明,MeSR45-1~MeSR45-4蛋白定位于细胞核和叶绿体,而MeSR45-5蛋白则定位于细胞核。
2.2 木薯SR基因家族系统进化树分析
本研究通过转录组测序分析筛选出与木薯可变剪接事件相关的SR45蛋白,从属于SR基因家族SR-like亚类。前人研究中木薯SR基因家族仅有6个亚类[18-19],因此通过Blast软件对已知的拟南芥SR蛋白序列在木薯蛋白数据库进行序列比对,筛选鉴定出木薯26种SR蛋白。对获得的木薯(26种)和拟南芥(20种)SR蛋白,利用MEGA 11重新构建系统发育进化树,如图1所示。整个系统发育进化树分为 SR、SC、SCL、RSZ、RS2Z、RS和SR-like 7个大类,其中SR45和SR45a聚为SR-like亚类。SR45分支由拟南芥SR45与木薯SR45-1~SR45-5组成,MeSR45-4和MeSR45-5亲缘关系更近;SR45a分支由AtSR45a和MeSR45a-1~MeSR45a-3组成,MeSR45a-1和 MeSR45a-2亲缘关系更近。

图1 SR蛋白系统发育进化树Fig.1 Phylogenetic evolution tree of SR protein
2.3 MeSR45蛋白保守Motif结构分析和基因结构分析
为了比较木薯MeSR45蛋白的相似性,通过MEME网站构建MeSR45蛋白Motif并利用TB-tools工具进行绘图。Motif分析表明Motif 3和Motif 6基序共同组成SR45 N端RS结构域,Motif 2和Motif 1基序共同组成RRM结构域。在木薯SR45 亚类的 5 个成员中均有 Motif 6、Motif 2 和Motif 1基序,表明这3个Motif是MeSR45蛋白序列的重要组成部分;从基序结构上来看,MeSR45-3与MeSR45-4蛋白具有相同基序,证明其蛋白结构与功能可能相似;MeSR45-1与MeSR45-2蛋白基序差异较小,与MeSR45-2相比,MeSR45-1缺失Motif 3基序且在中间位置多出 Motif 8基序;MeSR45-5蛋白基序较少且相对保守,在木薯中可能具有重要功能(图2A、2C、2D)。
从基因结构来看,木薯SR45亚类5个基因中,外显子与内含子分布差异较大,MeSR45-3、-4、-5预测基因内含子较少,其中MeSR45-4、-5被1个内含子分割成2个片段,MeSR45-3有3个外显子;MeSR45-1和MeSR45-2氨基酸序列中外显子与内含子较多,且编码区零散分布在7 kb的基因组上(图2B)。
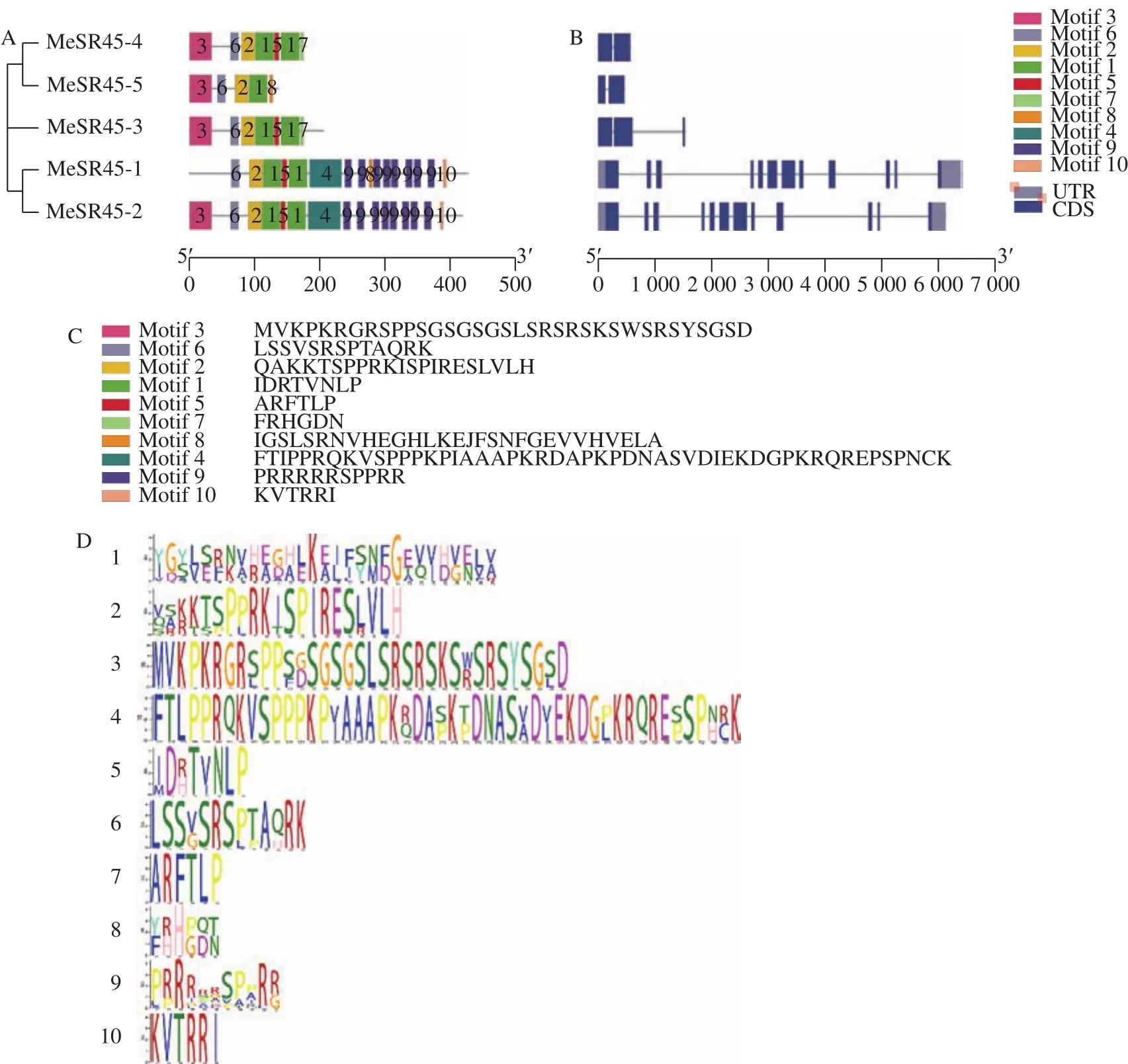
图2 MeSR45蛋白保守Motif结构和基因结构Fig.2 Consevered motif and gene structures of MeSR45 protein
2.4 木薯SR45蛋白结构分析
利用DNAMAN6.0把木薯的5个氨基酸序列进行多序列比对(图3),结果表明,多序列相似性比对结果为56.27%。MeSR45 N端结构域较为保守,有RRM结构域以及RS结构域,而C端仅有MeSR45-1和MeSR45-2拥有RS结构域,属于SR45基因。
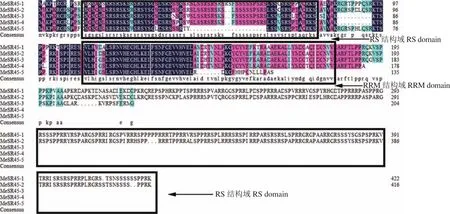
图3 MeSR45氨基酸序列比对Fig.3 Amino acid sequence alignment of MeSR45
2.5 干旱和低温胁迫诱导的木薯SR基因家族表达模式
为研究木薯SR基因家族在非生物胁迫(低温与干旱)条件下的表达模式,通过转录组测序数据分析(图4)发现,低温处理24 h后,木薯SR基因家族中,SR-like亚类SR45基因除MeSR45-3外均上调表达,其中MeSR45-2表达量最高;SR基因家族其他亚类基因MeRS2Z32、MeRS2Z33和MeRSZ22低温处理后表达量上升,而MeSCL33低温处理后下调表达,且表达量处于较低水平。木薯干旱处理后,仅有MeSR20和MeSR45-5表达量较高,MeSR45中仅有MeSR45-5上调表达,而SR-like亚类中其他SR45基因下调表达,MeSR45a-1表达量处于较低水平。

图4 木薯低温与干旱胁迫条件下SR基因家族表达热图Fig.4 Heat map of SR gene family expression in cassava under low temperature and drought stress
2.6 低温胁迫诱导的MeSR45基因表达模式
通过RT-qPCR分析木薯幼苗顶端叶片以及芽尖在 4 ℃ 低温分别处理 12、24、48 h 后,MeSR45基因家族成员的基因表达情况(图5)。木薯叶片低温处理后MeCOR、MeSR45-1、MeSR45-2、MeSR45-4、MeSR45-5表达水平显著提高,表明其均响应低温胁迫。MeCOR为木薯中响应低温的基因,MeCOR在 12、24、48 h 显著上调表达,在 12 h 表达最为显著,表明木薯受到低温胁迫。木薯SR45基因中MeSR45-2上调倍数最高,在低温处理24、48 h 后分别大约为未处理前 (0 h)的 2 100、2 500倍。MeSR45-1、MeSR45-4和MeSR45-5响应木薯低温,在处理12 h时的表达量达到最高。MeSR45-3在4 ℃低温处理0~48 h期间相对表达量表达较低,整体呈上升趋势,在48 h时基因相对表达量达到最大值,为木薯低温处理0 hMeSR45-3相对表达量的4倍。结果表明,木薯SR45基因在低温胁迫中可能具有相当重要的作用。

图5 低温胁迫下不同处理时间木薯MeSR45基因表达模式Fig.5 MeSR45 gene expression pattern in cassava under low temperature stress at different treatment time
2.7 MeSR45基因在木薯不同组织部位中的特异性表达
为研究木薯MeSR45基因空间表达模式,本研究运用RT-qPCR对木薯不同组织部位进行检测。结果(图6)表明,MeSR45-2在木薯叶组织中表达量最高,为根组织表达量的160倍左右,在茎与芽中表达水平较低。MeSR45-4和MeSR45-5基因在木薯根与叶中表达量较高,在茎与芽中较低。MeSR45-1在根与芽中表达量较高,在茎与叶中较低。MeSR45-3在芽中表达量较高,在其他组织中较低。这5个基因在木薯不同组织的表达模式差异较大,可能表明它们在不同组织中行使的功能具有差异。
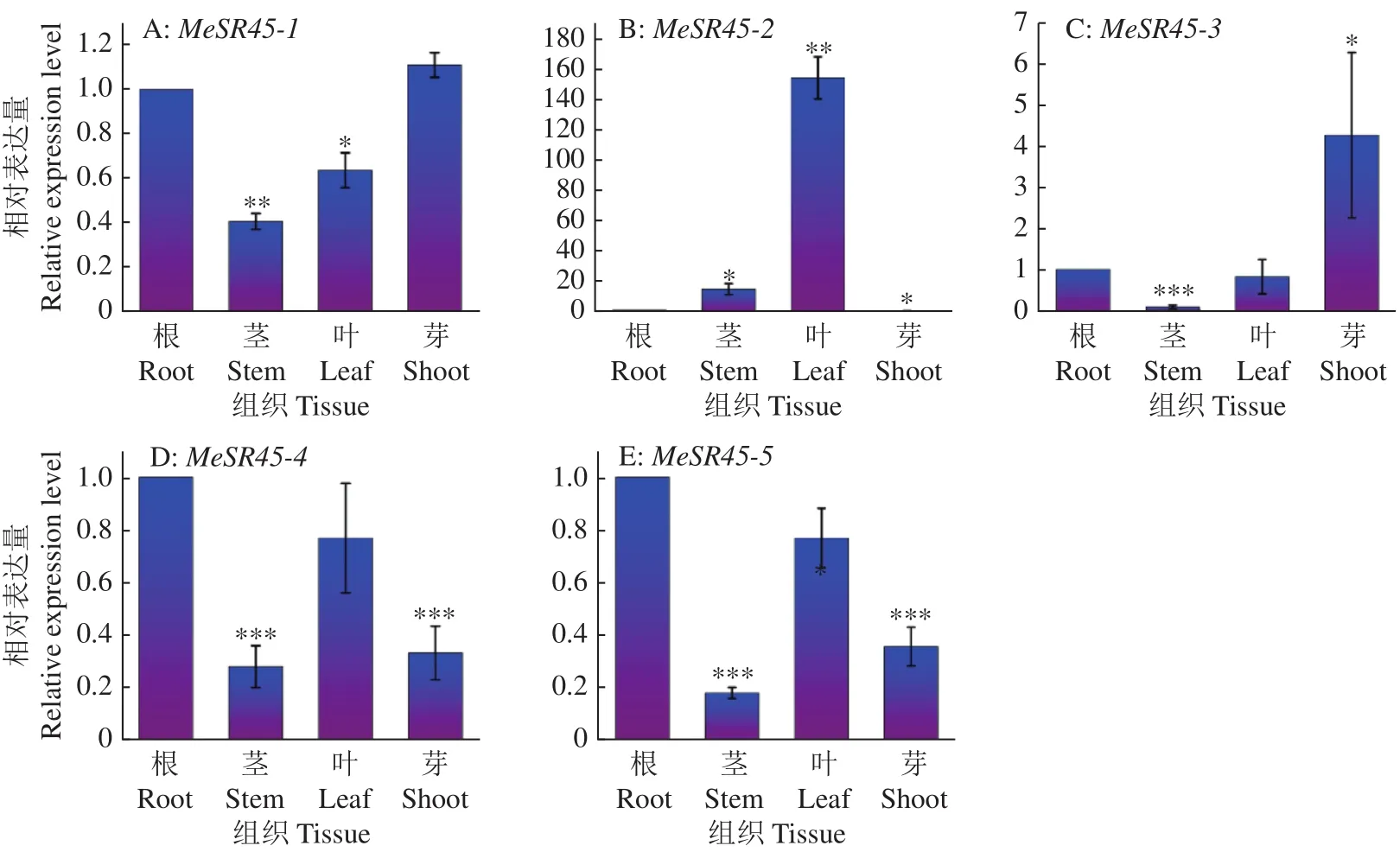
图6 MeSR45亚家族基因在木薯不同组织部位中的表达模式Fig.6 Expression patterns of MeSR45 subfamily gene in different cassava tissues
3 讨论与结论
3.1 讨论
SR45在植物细胞中是一类重要的剪接因子,可以参与植物生长发育过程并且具有应对非生物胁迫的作用[12,23]。在拟南芥中,SR45蛋白通过与mRNA前体、U1-70K和U2AF35b相互作用,分别将 U1 snRNP 和 U2AF 招募到 5′和 3′剪接位点,并调节可变剪接过程[24];拟南芥SR45蛋白缺失突变体SR45-1普遍表现出发育迟缓、根生长缓慢以及叶和花形态改变。2种拟南芥SR45可变剪接亚型具有不同的功能,一种亚型恢复根系生长,另一种亚型弥补突变体的花表型,这一过程表明SR45可变剪接具有重要功能意义[12,25-26]。在水稻中,OsFKBP20-1b在体内与OsSR45在细胞核和细胞质中相互作用,表明这2种蛋白可能共同参与了植物细胞核和细胞质的mRNA加工[27]。此外,SR45蛋白在应对盐胁迫时具有重要的作用,拟南芥中SR45缺失突变体导致盐胁迫敏感性增强,调控盐胁迫基因的表达与可变剪接发生变化,可变剪接长亚型SR45.1可以挽救盐胁迫敏感特性[22]。目前,植物SR45蛋白研究具有一定的进展,但是有关木薯SR45基因特性、系统进化与非生物胁迫响应的报道较少。
本研究在Gu等[18]研究基础上进一步鉴定木薯SR基因家族,补充SR-like亚类SR45、SR45a共8个基因;补充SC亚类MeSC35-2蛋白并去除RSZ亚类中MeRSZ23蛋白,MeRSZ23蛋白与RSZ亚类亲缘关系较远,影响SR基因家族系统进化树的构建,最终重新鉴定木薯SR基因家族为7大亚类26个成员。木薯基因结构与保守分析表明,MeSR45-1和MeSR45-2拥有相似的基序,并且外显子在基因中分布相似;而MeSR45-3和MeSR45-4保守基序相同,因此推测MeSR45-1和MeSR45-2、MeSR45-3和MeSR45-4可能存在基因复制事件。通过木薯SR45序列比对,发现5个氨基酸序列均有RRM结构域和N端RS结构域,但在C端MeSR45-3、MeSR45-4和MeSR45-5缺失RS结构域,这可能表明RS结构域在两端具有不同的功能,且N端RS结构域保守。有趣的是Tanabe等[28]发现AtSR45a蛋白的C端RS结构域是与 U1-70K、U2AF35b、AtSR45、AtSCL28、PRP38-like蛋白及其自身相互作用所必需的,而N端RS结构域则提高了相互作用的效率。因此,木薯SR45蛋白保守结构域的差异性有待进一步研究。
关于植物SR亚家族(SR45)成员应对非生物胁迫的研究相对较少,目前仅有部分模式作物在应对干旱胁迫与盐胁迫上具有相关研究进展[29]。因此,木薯作为一种热带低温敏感和耐贫瘠作物,发掘木薯SR45基因资源,对木薯以及其他热带作物应对低温、干旱胁迫以及其他非生物胁迫具有重要的意义。本研究发现木薯SR45基因与木薯应对低温胁迫具有密切的关系,全部成员均响应木薯低温,尤其是MeSR45-2基因,在低温处理48 h后基因表达量比未处理木薯上调约2 600倍,猜测MeSR45-2为SR45家族基因响应木薯低温胁迫最重要成员。基因组织特异性表达可以初步预测其相应的功能[30],在木薯组织表达中,MeSR45-2基因在叶片中特异性表达,而叶片为木薯响应低温胁迫的主要器官之一,因此MeSR45-2基因功能有待深入研究。除此之外,木薯根可能为木薯SR45蛋白响应其功能的重要组织。Zhang等[26]发现拟南芥SR45-1蛋白缺陷突变体相比正常拟南芥表现出根生长缺陷和花瓣缺陷表型,而拟南芥过表达SR45.2(SR45蛋白亚型2)株系可以恢复根生长缺陷的表型。本试验研究发现MeSR45-1、MeSR45-4和MeSR45-5在木薯根中表达量较高。因此在接下来的研究中,木薯的叶与根为主要研究器官。
3.2 结论
本研究通过生物信息学对木薯SR45基因亚家族成员进行分析,确定了木薯SR基因家族的分类和进化关系。通过转录组测序分析了低温、干旱胁迫处理下的基因表达特性,并通过RT-qPCR研究了低温胁迫条件下SR45基因的表达特性以及SR45基因的组织特异性表达模式。本研究表明SR45亚家族基因参与木薯低温和干旱胁迫应答和生长发育过程,而木薯SR45亚家族基因中MeSR45-2基因对低温胁迫显著响应,并且在木薯的叶片中表达量较高。该研究把MeSR45-2基因列为调控低温逆境变化的候选基因,同时将木薯的根与叶列为研究SR45基因亚类成员的主要植物组织,也为进一步研究木薯MeSR45基因应对低温等非生物胁迫提供了重要参考。
猜你喜欢
新医学(2023年10期)2023-12-09 15:04:51
传奇·传记文学选刊(2022年8期)2022-05-30 10:48:04
南方医科大学学报(2022年3期)2022-04-13 01:51:26
草业学报(2022年3期)2022-03-26 02:27:28
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10 07:07:36
世界热带农业信息(2018年11期)2018-01-17 10:08:19
中国实验诊断学(2017年5期)2017-06-05 15:03:23
广东第二课堂·小学(2017年5期)2017-05-27 20:09:13
上海精神医学(2014年6期)2014-12-08 08:14:51
中华临床免疫和变态反应杂志(2014年2期)2014-04-08 09:30:01
