
基于UPLC-Triple-TOF-MS/MS结合SWATH技术对不同泌乳期人乳脂质变化分析
2022-08-02 03:11王筱迪周金杭高文浩尹乾隆任皓威
食品科学 2022年14期
王筱迪,周金杭,曹 雪,高文浩,尹乾隆,任皓威,2,刘 宁,2,*
(1.东北农业大学食品学院,乳品科学教育部重点实验室,黑龙江 哈尔滨 150030;2.哈尔滨腾凝科技有限公司,黑龙江 哈尔滨 150030)
母乳喂养是为婴儿健康成长和发育提供理想食物的一种最佳方式,对婴儿和母亲的健康都有独特的生理和情感影响。人乳是6 个月内的婴儿最理想的天然食品。世界卫生组织提倡对0~6 个月婴儿母乳喂养,且继续母乳喂养至1 岁以上。母乳不仅为婴儿提供了超过50%的膳食能量需求,而且还作为一种载体在膳食中提供必需脂肪酸和脂溶性维生素。其中的脂质不仅作为婴儿神经和视网膜系统的结构组成部分,还可能通过抑制细菌肠毒素参与抗菌功能,保护婴儿的健康成长。在人乳所有营养成分中,人乳脂肪是最复杂的天然脂质之一,并显示出最大的可变性。人乳脂质的主要成分为甘油酯(glycerides lipid,GL)、甘油磷脂(glycerophospholipid,GP)、鞘脂(sphingolipid,SP)、甾醇脂(sterol lipid,ST)与脂肪酸及其衍生物(fatty acids,FAs)。
人乳是一个动态系统,其成分因母亲而异(人种和个体),并受到泌乳阶段、饮食、健康状况等因素的影响。人乳不是静态的流体,而是随着时间的推移而变化,尤其是泌乳阶段中脂质的变化部分地反映了婴儿的需求。虽然已有一些研究对不同泌乳期人乳中脂肪酸组成进行了研究,但很少有研究报道整个泌乳期过程中人乳脂质组成的变化。Shi Yudong等对内蒙古地区不同泌乳阶段人乳样品的化学成分进行了分析。Wang Yonghua等分析了广州地区初乳和成熟乳的脂肪酸组成。George等通过液相-离子迁移质谱仅对产后3 个月的人乳进行了详细分析,共鉴定出205 种甘油三酯(triglyceride,TAG),其中的98 种此前未在人乳中发现。Tu Anqi等利用超临界流体色谱-四重飞行时间质谱联用技术对来自中国不同地区的过渡乳、成熟乳和国外婴儿配方奶粉进行了比较分析。Wang Lina等将人的成熟乳与牛奶作比较,其中的215 种脂质可作为掺假分析的潜在生物标志物;而人乳与羊奶比较得出的生物标志物共有147 种。Garcia等通过气相色谱法测定法国母亲的初乳、过渡乳、成熟乳中TAG、总磷脂和缩醛磷脂的含量以及脂肪酸组成。但是由于不同国家人乳的脂肪酸组成不同,西方人乳中脂类的特征并不能完全反映中国人乳脂类的组成和结构,所以尽快建立属于中国的人乳脂质组学库十分重要。
上述研究中,数据均采用信息依赖型采集(information dependent acquisition,IDA),其应用广泛,检测准确,已在研究脂质组学方面奠定了基础,且数据分析相对简单。与传统的IDA采集模式相比,顺序窗口采集所有理论质谱模式(sequential window acquisition of all theoretical mass spectra,SWATH)扫描均匀且扫描范围广,依据峰面积定量,避免了灵敏度低与重现性差等问题。SWATH技术作为一项全新的超高效液相色谱质谱法的采集模式,可通过一次进样鉴定所有的脂质成分,具有灵敏度高和样品定量相关性高等优点。SWATH技术与超高效液相色谱-高分辨率串联质谱结合,使其同时具有高采集命中率和获得所有质谱信息的特点。目前,国内外尚未有研究使用SWATH采集模式对人乳中脂质成分进行检测,国外只有部分药物脂质代谢物或蛋白分析用到了SWATH检测模式。本研究将超高效液相色谱-三重四极杆串联飞行时间质谱(ultrahigh performance liquid chromatography-triple quadrupole time of flight tandem mass spectrometry,UPLC-Triple-TOF-MS/MS)与SWATH对不同泌乳阶段人乳中的脂质进行鉴定及相对定量分析,在建立中国人乳脂质组学库的同时为科学模拟各阶段母乳及开发不同段龄婴幼儿奶粉的研发提供参考依据。
1 材料与方法
1.1 材料与试剂
人乳取自黑龙江省哈尔滨市第一医院,采样对象为哈尔滨本地人或在哈尔滨生活3 a以上的22~36 岁足月分娩且身体健康的汉族产妇。采样经产妇及家属同意并签署知情同意书。
氯仿、氯化钠、异丙醇、乙酸铵、甲醇和乙腈 德国默克有限公司。试剂均为色谱纯。
1.2 仪器与设备
Acquity UPLCBEH C色谱柱(2.1 mmh100 mm,1.7 μm)、Acquity超高效液相色谱仪 美国Waters公司;Triple TOF5600质谱分析仪 美国SCIEX公司;BYN200-2氮吹仪 天津奥特赛恩斯仪器有限公司。
1.3 方法
1.3.1 人乳采集
产妇定时用吸奶器采集初乳(1~5 d)、过渡乳(6~20 d)和成熟乳(21 d以后),放入-80 ℃超低温冰箱中进行冷冻保存,在1 个月内完成实验。
1.3.2 人乳脂质提取
参考改良的Folch法。取人乳1 mL放入10 mL离心管,使用0.8 mL超纯水将其稀释,向其中加入4 mL氯仿-甲醇混合溶剂(2∶1,/),混合均匀。在输出功率150 W超声处理10 min后,加入0.2 倍体积的质量分数0.9%氯化钠溶液,旋混数秒。混合体系在1 854h离心15 min,静置2 min,分层后将有机相转移至螺口玻璃管中。之后再向水相中加入4.0 mL氯仿-甲醇混合溶液(2∶1,/),重复上述操作2 次,合并有机相并将其转移至玻璃管中。氮吹后,用1 mL氯仿-甲醇混合溶液(2∶1,/)复溶,过滤膜后等待检测。
1.3.3 液相色谱条件
色谱柱为Acquity UPLCBEH C(2.1 mmh 100 mm,1.7 μm);进样量5 μL;流动相A为水-甲醇-乙腈混合溶液(1∶1∶1,/)(含有2 mmol/L乙酸铵),B为异丙醇-乙腈混合溶液(1∶1,/)(含有2 mmol/L乙酸铵);流速0.35 mL/min;柱温35 ℃;分流比50∶1。梯度洗脱时间和流动相的比例见表1。
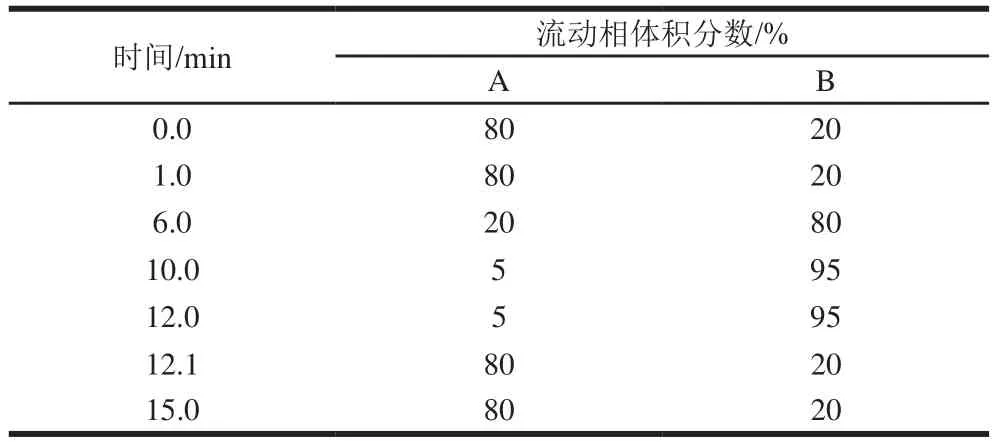
表1 人乳脂质分析的梯度洗脱条件Table 1 Gradient elution conditions for analysis of human milk lipids
1.3.4 质谱条件
电喷雾离子源,正离子模式条件:离子源喷射电压5 500 V;去簇电压80 V;碰撞能量10 eV;离子源气体1压力60 psi;离子源气体2压力60 psi;帘式气体压力35 psi;离子源温度550 ℃;质量扫描范围/50~1 700。负离子模式条件:离子源喷射电压-4 500 V;分解电位-80 V;碰撞能量-40 eV;离子源气体1压力60 psi;离子源气体2压力60 psi;帘式气体压力35 psi;离子源温度550 ℃;质量扫描范围为/50~1 700。
SWATH定量模式:正离子模式;间隔窗口3.8 u;从/50开始并增加到/1 700;每步的碰撞能量为35 eV,并且每次产物离子扫描施加15 eV的碰撞能量扩散,依次均匀扫描。
1.4 数据处理
每个样本做3 次平行。由Analyst 1.7.1软件得到的数据经PeakView 2.2和MasterView软件进行一级质谱分析,将得到的保留时间结果范围导入LipidView 1.2软件,并结合二级碎片分析定性脂质类化合物。脂质化合物的定量分析通过MultiQuant3.0.3和PeakView 2.2软件共同处理。所得数据应用Excel、SPSS软件进行单因素方差分析,化学结构式采用ChemDraw Professional 16.0软件进行绘制。
2 结果与分析
2.1 不同泌乳期人乳脂质的定性分析
参考本课题组优化的分析人乳中磷脂组学的方法进行实验参数设定。灵敏度、精密度、准确度、加标回收率均在规定范围内,可满足对样品测定的要求。检出限与定量限均可表明在所选用的实验条件下,仪器呈现较高的灵敏度。
不同类别的脂类化合物在二级质谱图中具有特殊的裂解规律,根据不同位点的化学键断裂产生的中性丢失碎片离子或特异性二级碎片(大部分为[M+H-FA]碎片离子)推导出附着在甘油分子上的单个FAs,以鉴定脂质组分的结构。图1以磷脂酰乙醇胺(phosphatidylethanolamine,PE)为例,在负离子模式下,PE 38:2生成/770.724的[M-H]准分子离子,/140.007和/196.025为PE的特征离子碎片,/480.229代表失去了一个氢原子的断裂的溶血型磷脂酰乙醇胺(lysophosphatidylethanolamine,LPE,18:0)单链,/283.235和/307.228分别是断裂的C和C脂肪酸失去一个氢原子产生的二级碎片。针对于PE 38:2这种物质,不同长度的酰基链和不同的酰基链双键数目形成了许多可能的酰基链组合。除了上述组合方式外,还有/281.218和/309.242、/279.207和/311.258,它们分别代表PE 38:2(18:1/20:1)和(18:0/20:2)。但由于其强度最高,PE 38:2主要以C和C脂肪酸组成为主。一般来说,酰基链随不同的长度(即不同的碳原子数)、不同的不饱和度、潜在的支链等而变化。脂质头基和酰基链的这些变化形成了大量的脂质分子种类,提升了脂质定性分析的难度。

图1 PE 38:2在负离子模式下的二级质谱图Fig.1 MS/MS spectra of PE 38:2 in negative ion mode
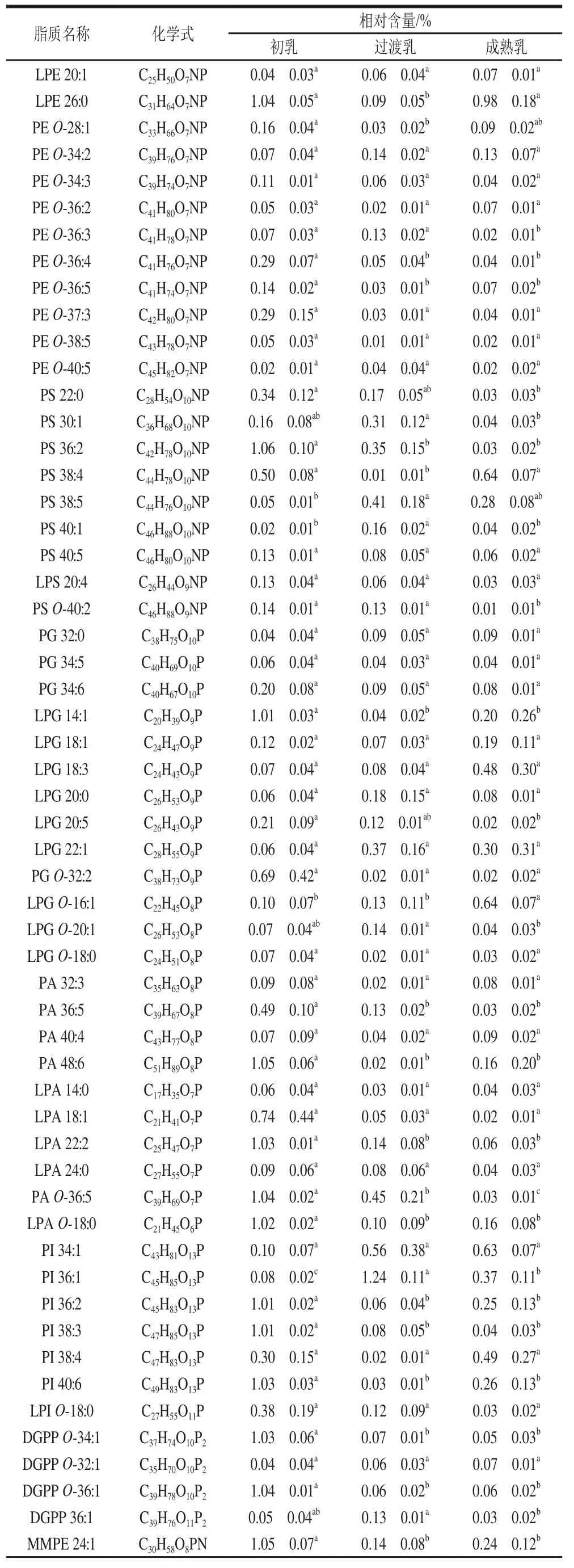
按照以上结构鉴定方法,结合高通量、高覆盖度、高准确性的SWATH采集技术在正负离子下检测到的脂质化合物共229 种,如表2所示。除了检测人乳中含量丰富的脂质,如GL和GP外,还一同测定了SP、ST和FAs,它们在整个泌乳阶段含量较低且种类较少。由于SWATH采集技术可以高效测定极低丰度分子的特性,本研究检测到10 种ST,如麦角甾醇(Erg)、脂质白蛋白纳米颗粒(lipid-albumin nanoparticles,LAN)、去氧孕烯(Des)等。FAs共4 种,为游离脂肪酸(free fatty acid,FFA)FFA 14:1~22:6之间。GP是种类最丰富的脂类,共108 种。其中PE及LPE共33 种,是整个泌乳期中磷脂种类最丰富的;其次是磷脂酰胆碱(phosphatidylcholine,PC)及溶血型磷脂酰胆碱(lysophosphatidylcholine,LPC)类脂质共21 种;磷脂酰丝氨酸(phosphatidylserine,PS)及溶血型磷脂酰丝氨酸(lysophosphatidylserine,LPS)共8 种;磷脂酰甘油(phosphatidylglycerol,PG)及溶血型磷脂酰甘油(lysophosphatidylglycerol,LPG)共13 种;磷脂酸(phosphatidic acid,PA)及溶血型磷脂酸(lysobisphosphatidic acids,LPA)共10 种;磷脂酰肌醇(phosphatidylinositol,PI)及溶血型磷脂酰肌醇(lysophosphatidylinositol,LPI)共7 种;其他类的GP共16 种。SP共22 种,其中鞘磷脂(sphingomyelin,SM)12 种。SWATH技术将整个质谱扫描范围分为若干个窗口,高速、循环地对每个窗口的所有离子进行碎裂、扫描,从而无遗漏、无差异地采集全部子离子信息,拥有更高的覆盖度。除常见物质己糖神经酰胺(HexCer)外,通过SWATH采集技术还发现了硫苷脂(SGalCer)、1-磷酸神经酰胺(CerP)、神经酰胺磷脂酰乙醇胺(PE-Cer)、神经酰胺磷脂酰肌醇(PICer)、长链碱基(LCB)等。其中SGalCer是通过硫酸基与半乳糖部分的3-羟基的酯化反应从葡萄糖神经酰胺中衍生出来的物质,LCB则是鞘氨醇的衍生物,然而这些极低丰度的物质在脂质数据库中不常见,如LIPID MAPS数据库中的脂质。人乳中的GL主要包括甘油二酯(diglyceride,DAG)和TAG,还有一些不常见的物质如单烷基二酰甘油(monoalkyl diacylglycerol,MADAG)等。GL是整个泌乳阶段中含量最多的脂质,同时由于甘油不同位点上的羟基被不同碳数、双键数、双键位置和构象的脂肪酸酯化形成,导致GL共81 种,其中包括24 种DAG、28 种TAG(TAG 34:0~64:12)和其余29 种GL。虽然本研究检测到的TAG种类并不是很多,但仅仅这28 种TAG就在成熟乳中占总脂质化合物总含量的一半。

表2 不同泌乳期人乳中脂质种类及其相对含量Table 2 Types and relative contents of lipids in human milk during different lactation periods
续表2

续表2

续表2

续表2
2.2 不同泌乳期人乳脂质的相对定量分析
由图2可知,初乳中5 类脂质的相对含量分别为GL 68.44%、GP 19.13%、SP 6.15%、ST 4.02%、FAs为2.27%;过渡乳中5 类脂质的相对含量分别为GL 69.63%、GP 18.65%、SP 6.22%、ST 3.67%、FAs为1.83%;成熟乳中5 类脂质的相对含量分别为GL 71.04%、GP 17.55%、SP 6.54%、ST 3.52%、FAs为1.35%。其中,GP和SP中的SM属于磷脂(phospholipid,PL)。这些PL在初乳、过渡乳、成熟乳中的相对含量分别为22.95%、21.86%、21.07%(表2),在整个泌乳阶段呈下降趋势,与Ingvordsen Lindahl、Bitman等研究发现一致。此外,60%的PL和85%的ST与乳脂球有关,作为乳脂球膜的结构成分存在于乳中。上述PL和ST的下降表明,随着哺乳期的变化,脂肪球周围的膜厚度减少,乳脂肪球的大小随着哺乳的进行而增加。人乳中总脂质含量随泌乳期时间的延长而增加,并与PL呈反比关系。
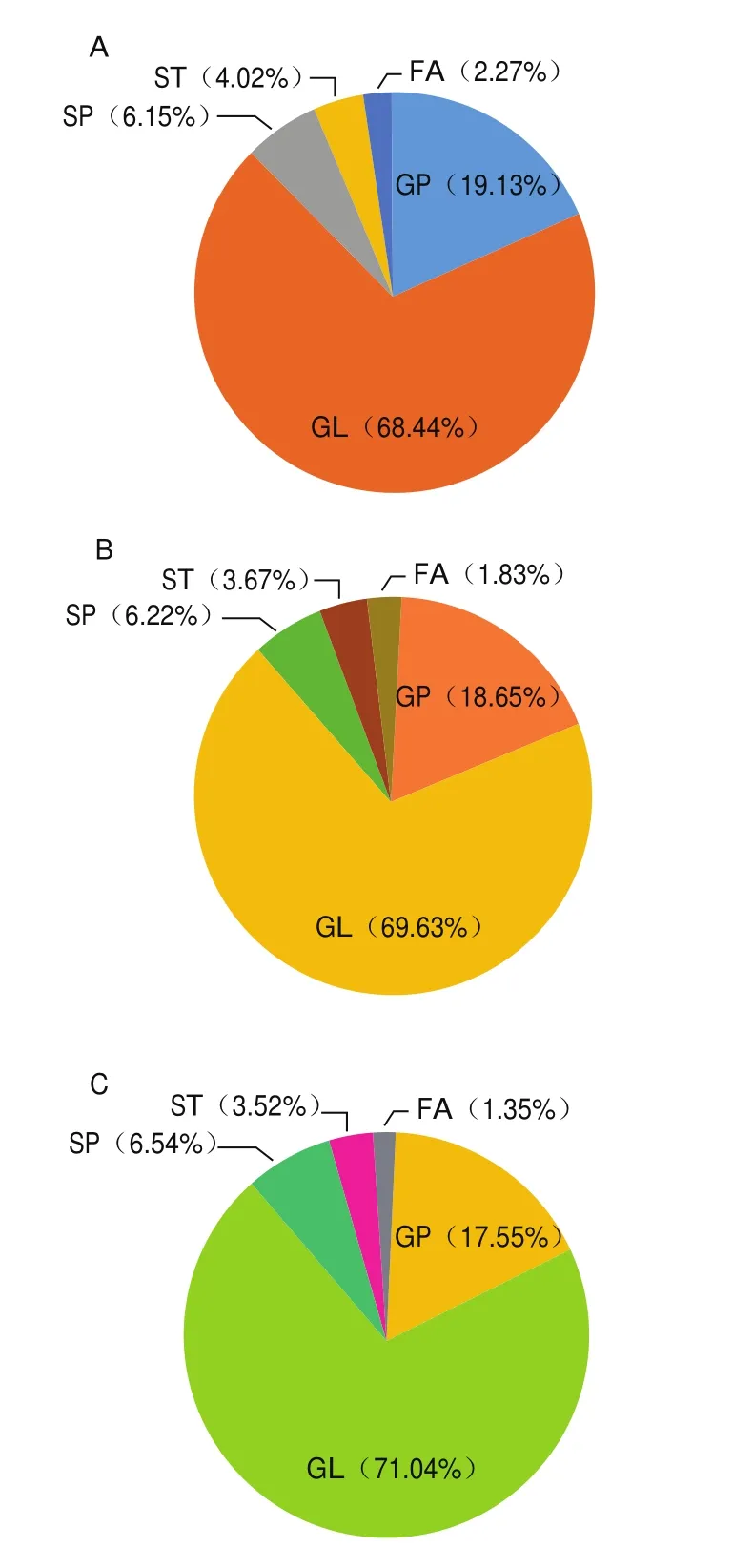
图2 初乳(A)、过渡乳(B)、成熟乳(C)中每类脂质占总脂质的相对含量Fig.2 Relative contents of each class of lipids in colostrum (A),transitional milk (B), and mature milk (C)
2.3 不同泌乳期人乳极性脂质相对含量分析
人乳中各PL的相对含量随着泌乳期的变化也处于动态变化中,如图3所示。文献表明无论在初乳、过渡乳还是成熟乳中,PC、PE、SM为PL主要组成。然而在本研究中(图3),初乳中相对含量最多的PL是PA,约占总PL的20.7%,其相对含量随着泌乳期的延长显著下降,并且成熟乳中的PA含量明显低于其他PL。在整个泌乳期中,PS和PI的相对含量相对恒定,但在初乳中略高于成熟乳。有研究表明,PI所含的-6系列的花生四烯酸(arachidonic acid,ARA,20:4)仅次于PE。由图3可知,从初乳到成熟乳SM含量显著增加。PE和PG在过渡乳中的含量明显低于其他乳,并在成熟乳中达到最大值。PE和SM在成熟乳中的含量相似(22.2%、22.4%),是成熟乳中最丰富的PL。虽然在本研究中PE的含量为先下降后上升,与Harzer、Bitman等研究有所区别,但在成熟乳中PE占总磷脂的含量大体相似(约为20%)。此外,PC呈先上升后下降,在过渡乳中含量最高,约占总磷脂的37.6%(图3)。由图3可知,PC在整个泌乳期磷脂类中所占比例最大(占总磷脂的67.30%),其次是SM(占总磷脂的53.51%)与PE(占总磷脂的52.44%)。然而,Cilla等报道PL在人乳中的相对含量在不同研究之间存在显著差异。何扬波表明,除PC外,其他磷脂都随泌乳期的变化稳步升高。Sala-Vila等却认为SM和PC占磷脂的主要部分,同时SM在整个泌乳阶段含量相对恒定。Bitman和Benoit等报告,SM是人乳中含量最多的磷脂。而Wang Liwen和Ingvordsen Lindahl等观点相同,都认为PE是整个泌乳期中最主要的磷脂。但人乳的成分是一个动态的过程,并受多种因素的影响,目前尚未确定其真正标准组成。
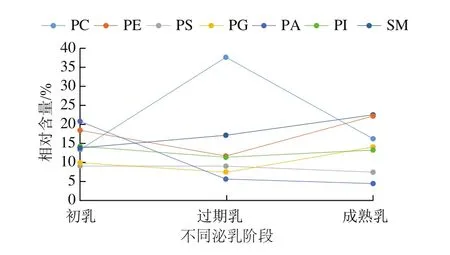
图3 人乳中不同极性脂质随泌乳期的含量变化Fig.3 Changes in percentages of different polar lipids in human milk with lactation time
2.3.1 不同泌乳期人乳磷脂亚类相对含量分析
由表2可知,在不同泌乳期人乳的磷脂酰甘油中,各个泌乳阶段中含量最多的PG均为LPG。PG在本研究中拥有最多溶血磷脂种类,共有9 种。其中,含量最多的为初乳的LPG 14:1,约占总脂质的1.01%,与其他泌乳期人乳的LPG 14:1含量存在显著差异。携带二十碳五烯酸(eicosapentaenoic acid,EPA)的LPG 20:5也在初乳中含量较高,并随泌乳时间的延长相对含量下降。LPG是PG中的主要成分,在整个泌乳时期占总PG含量的64%以上,尤其占成熟乳总PG含量的90%。LPE占初乳PE总含量的47%。Van Beusekom等发现,加勒比海地区母亲的成熟乳汁中PE的相对含量约为13%,LPE约占其PE含量的54%。这些溶血磷脂是指其脂肪酸链已从-1或-2位移除的磷脂,鉴于它们来源的不确定性,如此高比例的溶血磷脂很可能是在乳样品冷藏过程中发生水解。由表2可知,在整个泌乳阶段共测得24 种溶血磷脂(数量高于Wang Lina和Ingvordsen Lindahl等的研究结果),其中可以为婴儿提供胆碱的LPC只有LPC 16:0,且在初乳中的相对含量最多并与其他乳存在显著差异,约为总脂质的0.44%,并随泌乳期的延长,含量显著降低。胆碱作为神经传递过程中乙酰胆碱的前体对婴儿的脑发育存在重要作用,同时还参与构建细胞膜的结构。PC和SM都是胆碱的重要来源,约占人乳中胆碱含量的17%。由表2可知,在整个泌乳期的PC中,LPC 16:0、PC 34:3/PC 35:1/PC-36:4、PC 35:4分别为初乳、过渡乳、成熟乳中含量最多的脂质,分别占总脂质的0.44%、1.24%和0.68%。PC的主要脂肪酸组成依次为C、C、C、C,大多数研究表明,这4 种脂肪酸共占PC总脂肪酸的90%。然而对于最不饱和的磷脂——PE来说,如表2所示,初乳中LPE 18:2为PE中含量最多的脂质,占总脂质的1.08%;过渡乳中PE 38:4(18:0/20:4)含量最多,占总脂质的0.18%;成熟乳中LPE 26:0是含量最多的脂质,占总脂质的0.98%,其次是PE 40:6(18:0/22:6),占总脂质的0.65%;同时LPE 26:0、LPE 18:2、PE 40:6(18:0/22:6)也是整个泌乳阶段含量最多的PE,如图4所示。PE比PC、SM含有更丰富的长链多不饱和脂肪酸,如:二十二碳六烯酸(docosahexaenoic acid,DHA)、EPA、ARA等,这与Ingvordsen Lindahlida等研究一致。观察本研究的PL特性发现PC中有4 种脂质分子含有长链多不饱和脂肪酸,PE共8 种,其他PL中仅含1 种或2 种。由表2可知,含有长链多不饱和脂肪酸的PE(PE 38:5(18:1/20:4)、PE 40:4(18:0/22:4)、PE 40:6(18:0/22:6或18:1/22:5)),在成熟乳中含量最高,初乳中含量最低。尤其是PE 40:4及PE 40:6在成熟乳中的含量与其初乳中含量存在显著差异。PE作为含多不饱和脂肪酸最多、饱和脂肪酸最少的磷脂,提供了婴儿所需的重要生物活性成分,如ARA、EPA和DHA,对新生儿的神经和视觉发育、生长至关重要。

图4 不同泌乳期PE组成Fig.4 Composition of PE in different lactation periods
2.3.2 不同泌乳期人乳鞘脂含量及特性分析
由表2可知,在不同泌乳期的SM中,SM 36:1; 2、SM 40:2; 2、SM 36:2; 2分别为初乳、过渡乳、成熟乳中SM含量最多的脂质,分别占总脂质的1.09%、1.26%、0.80%。且从初乳到成熟乳,SM含量显著增加(分别占总PL的13.92%、17.18%和22.41%)。张雪等表明,SM与GP的脂肪酸组成存在显著差异,SM所含的饱和脂肪酸在所有磷脂中最高,多不饱和脂肪酸含量最低。由于SP含有-酰胺化的脂肪酰基,通常比磷脂的脂肪酰基长得多,这也意味着SP通常含有比GP中最常见的脂肪酰基更长的脂肪酰基;而SP具有的“双峰”物种分布(图5)说明其主要含有的-酰胺化C为最短脂肪酸组成,C~C为最长,很少存在-酰胺化C和C。
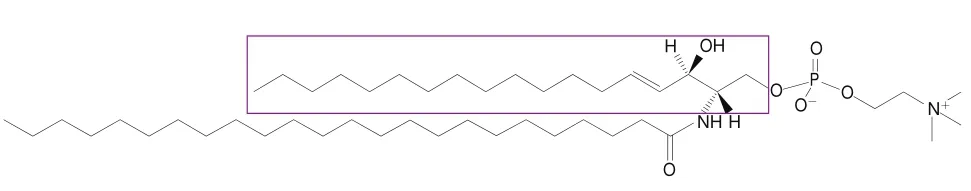
图5 鞘脂的“双峰”结构图Fig.5 “Bimodal” structure of sphingolipids
SM中的多不饱和脂肪酸还含有C和C。C即为人们常说的神经酸(nervonic acid,NA)天然存在于人乳中(<2%),主要在SM中酯化,即使在含多不饱和脂肪酸最多的PE中也鲜有存在。NA是神经元膜的重要组成部分,其在早期髓鞘形成和预防营养不良等问题中起着至关重要的作用,与DHA和ARA一起对新生儿的神经发育有积极的影响。由表2可知,含有NA的SM 40:1; 2(16:0/24:1)、SM 40:2; 2(16:1/24:1)、SM 42:2; 2(18:1/24:1)在过渡乳中的相对含量远高于其他两个泌乳阶段,尤其是SM 40:1; 2在过渡乳中的含量与其他泌乳阶段均存在显著差异。Yu Jiahu等表明,在成熟乳中NA的含量会减少25%,且个体间NA含量的差异可能主要由内源性因素引起,受母体饮食的影响较小。
2.4 不同泌乳期人乳中性脂质相对含量分析
中性脂质主要包括TAG、DAG,并以TAG作为主要脂质。在整个泌乳阶段,人乳中TAG的不同脂肪酸组成对婴幼儿具有不同的潜在作用,这对制备用于婴儿营养的合成TAG尤为重要。考虑到这些中性脂质在人乳中随泌乳的进行而不断变化,可假设它们反映了婴儿成长发育的部分需求并具有生物学重要性。此外,在选择脂肪源以制备婴儿配方食品时,特殊TAG、DAG的存在可能是重要的,配方食品应尽可能含有TAG“标记”,并且其平均浓度应与不同阶段人乳中发现的浓度相似。本研究并未对TAG中脂肪酸酯化位置进行探寻,故下列组成并不表明脂肪酸在甘油分子中的位置。
如图6所示,在整个泌乳阶段,只有初乳时DAG的总量略高于TAG。过渡乳和成熟乳中,TAG的急剧增加导致TAG分别占总物质的40.63%和49.32%,使TAG成为人乳中的主要脂质组分。由表2可知,在DAG中,DAG 34:3为初乳中含量最多的脂质,由两种脂肪酸组成(16:0/18:3或16:1/18:2),占总脂质1.91%;DAG 34:0(16:0/18:0)为过渡乳中含量最多的脂质,占总脂质的1.24%;DAG 36:1(18:0/18:1)为成熟乳中含量最多的脂质。然而,Wang Lina等表明DAG(18:2/18:2)、DAG(18:1/18:1)、DAG(14:0/14:0)、DAG(18:1/12:0)是人乳样品中最常见的DAG种类。由图7和表2可知,初乳中TAG 34:0为TAG中含量最多的脂质,占总脂质的1.42%;在过渡乳中,相对含量超过2%的TAG共有5 种,以TAG 38:0含量最多(占总脂质的7.42%),其次是TAG 40:2(10:0/12:0/18:2)(占总脂质的6.07%);在成熟乳中,虽然TAG 38:0和TAG 34:0是TAG中含量最多的两种脂质,共占总脂质的14.45%,但其余含量较多(>2%)的TAG均由不饱和脂肪酸组成(以TAG 52:4(16:0/18:2/18:2)为首),共占总脂质的18.97%。在3 个泌乳阶段中,虽然仍是含有饱和脂肪酸的TAG相对含量较高,但随着泌乳期的延长,含不饱和脂肪酸的TAG相对含量激增,并在成熟乳中共占总脂质的31.61%,约为成熟乳中饱和脂肪酸组成的TAG含量总和的1.78 倍。TAG中亚麻酸和亚油酸等长链多不饱和脂肪酸含量的升高,可能是由于生长中婴儿对于多不饱和脂肪酸的需求增加,亚麻酸和亚油酸作为DHA、ARA的前体物质也需要更多的储备。
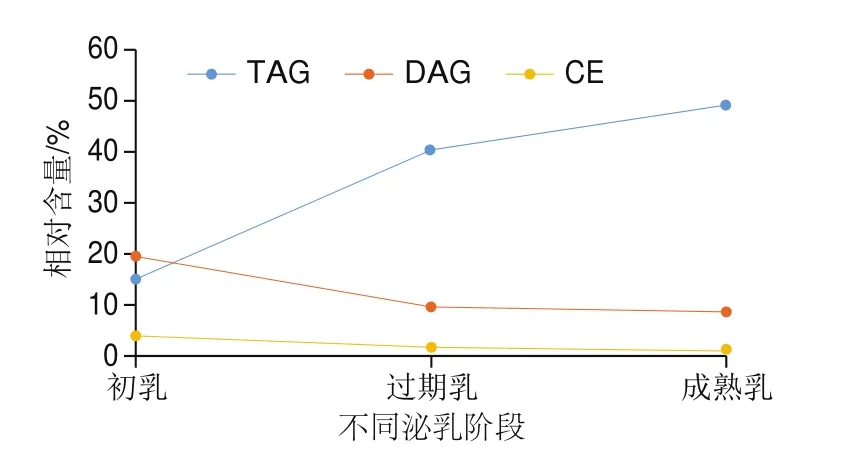
图6 人乳中不同脂质亚类随泌乳期的含量变化Fig.6 Changes in percentages of different lipid subclasses in human milk with lactation time

图7 不同泌乳期的人乳TAG组成Fig.7 Composition of TAG in human milk at different lactation periods
2.5 不同泌乳期人乳含有DHA的TAG含量分析
Liu Zhiqian等从牛乳、人乳和不同品牌的婴儿配方奶粉中共检测出56 种含DHA的TAG,婴儿配方奶粉含有56 种中的大部分。本研究只发现了4 种含有DHA的TAG,分别为TAG 54:8(14:0/18:2/22:6)、TAG 56:11(14:0/20:5/22:6)、TAG 64:8(18:2/24:0/22:6)、TAG 64:12(20:0/22:6/22:6),它们一般是中等或低丰度的。其中,TAG 64:12含有2 个DHA残基,这种含有2 种或2 种以上DHA分子的TAG主要存在于婴儿配方奶粉中,很少存在于人乳。对这4 种TAG进行统计学分析(图6及表2),发现TAG 54:8在3 个泌乳阶段均存在显著差异,其相对含量从初乳到成熟乳逐步增加(分别为0.22%、1.25%和2.63%);TAG 64:8在过渡乳中与其他两个泌乳阶段存在显著差异,其中在过渡乳中的相对含量分别是在初乳和成熟乳中的9.3 倍和3.7 倍;TAG 56:11和TAG 64:12的含量随泌乳期而增长,但其增长幅度小于其他TAG。婴幼儿依赖于母体循环中预先形成的DHA供应(无论是来自母体饮食、组织储备还是-亚麻酸的合成)以及自身亚麻酸的合成。从妊娠的最后3 个月到两岁,婴儿大脑每天对DHA和ARA的需要量约为3 mg。由于婴儿必需脂肪酸去饱和链延长的能力有限,自身转化率约为1%,合成速率明显不足。妊娠晚期补充DHA(200 mg/d)可防止母体DHA水平的下降。这可能是由于在成熟乳中,母亲在孕期补充的营养物质及组织储备已基本消耗完毕,因此与其他短链脂肪酸相比,富含DHA残基的长链多不饱和的TAG相对含量逐渐下降。Garcia等对实验数据进行估量计算后表明,仅有15%和44%的人乳样本可以满足早产儿和足月儿对DHA的要求。
2.6 不同泌乳期人乳甾醇脂类与游离脂肪酸含量分析
ST中的胆固醇作为乳脂球膜的一种,随着泌乳的进行呈下降趋势(图2)。与Bitman等的结论相似,在初乳中的胆固醇含量最高,并在过渡乳和成熟乳中逐渐降至较低水平。由表2可知,初乳中的CE 16:1和CE 22:1含量较高,分别占总脂质的1.90%和1.59%,与其他泌乳阶段的含量存在显著差异;CE 2:0和CE 14:1分别是过渡乳和成熟乳中含量最多的胆固醇。胆固醇不仅对神经系统的神经元发育很重要,还与激素和VD的合成有关。同时有研究表明,母亲的饮食对人乳中的胆固醇并没有显著影响,且经母乳喂养的婴儿血浆中总胆固醇和低密度脂蛋白胆固醇的浓度高于经婴幼儿配方奶粉喂养的婴儿血浆。与配方奶粉喂养相比,母乳喂养与生命后期较低的血液总胆固醇浓度有关(成年后血液胆固醇降低0.15 mmol/L),可使心血管疾病的人群患病率降低5%。尽管围绕点估计值的置信限和观察到的研究间异质性并不排除母乳喂养对心血管的潜在有益或不利影响,但对现有数据的荟萃分析也不能得出关于母乳喂养和成人心血管疾病全因死亡率之间关系的明确结论。如表2所示,不同泌乳期的FAs中,FFA 20:0为初乳和过渡乳中含量最多的脂质,分别占总脂质的1.89%和0.42%;FFA 22:4为成熟乳中含量最多的脂质,占总脂质的0.64%。初乳中的总FAs含量最高。
3 结 论
采用具有更高的质量精确度和灵敏度检测技术,详细分析人乳中脂质的分子种类、脂肪酸酰基组成、相对含量的差异。利用UPLC-Triple-TOF-MS/MS结合SWATH采集技术并使用定义的窗口来同时获取所有前体离子以获得更宽的动态范围。根据/和二级质谱图,共鉴定出229 种脂质化合物,其中PE 33 种、PC 21 种、PS 8 种、PG 13 种、PA 10 种、PI 7 种;SP共22 种;DAG 24 种,TAG 28 种;ST共10 种;FAs共4 种;以及众多不在常见脂质数据库中的脂质。然而,本研究并未对TAG中脂肪酸酯化位置进行探寻,也未详细的对人乳中的糖脂等进行分析,后续实验仍需要从TAG位置,从更精密、更有效的微量成分、生物活性物质入手,进一步分析人乳脂质成分,着力生产出尽可能与母乳100%相接近并满足婴儿不同阶段生长的需求的婴儿配方奶粉。
猜你喜欢
动物营养学报(2022年9期)2022-10-12
医学概论(2022年4期)2022-04-24
昆明医科大学学报(2021年4期)2021-07-23
智慧健康(2021年33期)2021-03-16
中国乳业(2019年12期)2020-01-17
河南畜牧兽医(2020年9期)2020-01-11
分析化学(2018年4期)2018-11-02
现代畜牧科技(2018年7期)2018-10-21
国外畜牧学·猪与禽(2018年4期)2018-05-14
现代畜牧科技(2018年3期)2018-05-14
