
基于生命表的大兴安岭兴安杜鹃(Rhododendron dauricum)种群数量特征及动态分析
2022-07-30 11:10卢玉鹏王小玲何兴元
生态学报 2022年13期
卢玉鹏,陈 玮,王小玲,高 柱,何兴元,*
1 江西省科学院生物资源研究所, 南昌 330096
2 中国科学院沈阳应用生态研究所, 沈阳 110016
兴安杜鹃(Rhododendrondauricum)俗名达子香,是一种分布于大兴安岭、小兴安岭和长白山的灌木。兴安杜鹃是我国东北森林区灌木层中优势种,是杜鹃属植物在我国分布的北界[1]。研究发现,我国兴安杜鹃的遗传多样性水平较高,长白山脉和辽东半岛可能是其遗传多样性中心,而俄罗斯境内兴安杜鹃遗传多样性最高的是西西伯利亚南部的居群[2—3]。在人工栽培方面,兴安杜鹃以种子进行苗木繁殖不存在问题,但幼苗形态建成较缓慢,且对光照和上层树种有一定要求,适宜林下种植[4—6]。在药用方面,兴安杜鹃的叶中含有杜鹃素、伞形花内酯、东莨菪素、杜鹃醇等物质,可治疗慢性支气管炎、冠心病等疾病;根中含强心苷、香豆素内酯及三萜化合物,可以治疗急性痢疾、肠炎[7—8]。此外,兴安杜鹃中还被提取分离出具有抗HIV活性、抗氧化活性和抑制组胺释放活性的物质[9—10]。目前兴安杜鹃的野生种群受到关注较少,种群的数量特征未见报道,种群的生存现状不是十分清楚。
生命表广泛应用于种群数量研究[11],其中静态生命表的核心思想是“空间代替时间”,关键是对植物年龄的划分,通常使用一些易于测量和识别的特征来划分年龄结构,比如胸径(地径)、高度、叶片疤痕等。静态生命表操作方便可以在短时间内取得数据,又可延伸出生存分析函数、时间序列分析等数量分析方法[12—14]。因此对于生长周期较长的植物,多采用静态生命表进行种群结构与动态分析。金慧等[15]采用生命表分析了长白山区濒危植物朝鲜崖柏(Thujakoraiensis)的种群数量,发现种群龄级结构呈“∩”,属于衰退型种群,应适当采取人工抚育。拓锋等[16]采用生命表分析了祁连山青海云杉(Piceacrassifolia)种群数量动态,发现种群中幼苗和小树储量丰富,存活曲线为Deevey Ⅱ型,为稳定增长型种群,应加强对幼龄级植株生存环境的保护和改善。应用静态生命表分析技术可以快速理解兴安杜鹃种群生存现状和发展趋势。
种群结构与动态,除与物种自身生物学特征有关外,还会受到非生物和生物环境影响[17—18]。海拔作为一个综合性环境因子,与气候、土壤、植被等多种因子相关,是影响植物种群分布的重要环境因子[19]。林型是影响郁闭度、林下小气候和林下植被的重要生物因素,兴安杜鹃作为一种林下灌木,与上层林木(林型)密切相关[20]。因此通过分析大兴安岭兴安杜鹃种群统计特征,并分析种群统计特征与海拔和林型的关系,进而掌握种群的生存现状、发展动态及其影响机制,为科学合理地利用和保护野生兴安杜鹃资源提供科学依据和参考建议。
1 研究材料和方法
1.1 样地设置和群落调查
调查样地隶属于黑龙江省大兴安岭地区的呼中区,位于大兴安岭的北部山脉伊勒呼里山北坡(51°14′40″— 52°25′N, 122°39′30″— 124°21′E),总面积约7.42×103km2(图1)。该地区属于寒温带大陆性季风气候,夏季较短,冬季漫长且寒冷,年平均气温-4.3℃,绝对最低温度-52℃,绝对最高温度32℃,年平均降水量497.7mm,年平均蒸发量905.7mm,年平均日照时数2365h,积温1800—2000℃(≥10℃),年度无霜期80—100d(图2)。呼中区的平均海拔是812m,最高的两座山峰为大白山(伊勒呼里山主峰,大兴安岭北部最高峰)海拔1528m,小白山海拔1404m,最低海拔420m,位于呼玛河北部。土壤类型有棕色针叶林土、草甸土、沼泽土、河滩森林土和石质土,其中最具代表性的是棕色针叶林土。植被属于泛北极区系中的东西伯利亚区系,以西伯利亚区系为主,与东北区系和蒙古区系混合,地带性植被类型为寒温带针叶林。乔木层的优势种主要有落叶松(Larixgmelinii)、樟子松(Pinussylvestris)、红皮云杉(Piceakoraiensis)、白桦(Betulaplatyphylla)、山杨(Populusdavidiana)、钻天柳(Choseniaarbutifolia)等;灌木层的优势种主要有偃松(Pinuspumila)、兴安杜鹃、杜香(Ledumpalustre)和越橘(Vacciniumvitis-idaea)等;草本层的优势种主要有小叶章(Deyeuxiaangustifolia)、薹草属(Carex)和野豌豆属(Vicia)等[21—22]。
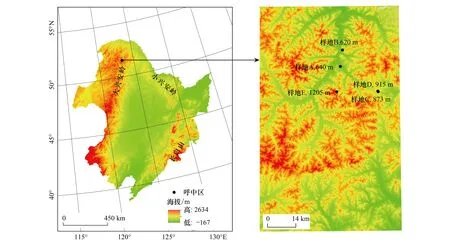
图1 研究区及样地设置信息Fig.1 The information of study area and plots样地设置中,样地C和样地D经纬度接近,但坡位和坡向不同

图2 研究区生物气候图解Fig.2 The bioclimatograph of study area
2018年5月—8月选择三个海拔梯度(600m、900m和1200m)共设置了5个面积为60m×60m样地(图1,表1)。样地A、B为600m海拔梯度,样地C、D为900m海拔梯度,样地E为1200m海拔梯度。在每个样地,用手持GPS和地质罗盘测量和记录了经纬度、海拔、坡向、坡度、坡位、林型、林龄和郁闭度等信息,开展群落调查。
1.2 兴安杜鹃龄级测量和划分
采用标准木方法,通过树木年轮分析仪测量标准木的年龄,建立兴安杜鹃地径与年龄的关系模型。在所有样地内,测量和记录了兴安杜鹃的地径,同时采集部分植株的地径部分用于测量年龄。将取样的地径部分切成2cm左右的小段后,用砂纸打磨至年轮清晰可视后,利用树木年轮仪(Olympus—SZX7)测量了取样植株的年龄[23]。然后利用SigmaPlot12.5软件对兴安杜鹃年龄和地径进行了建模拟合。根据拟合结果以及样地内兴安杜鹃个体径级大小,多数个体地径在0—10mm范围内,10—12mm的个体数量较少,而12mm以上的个体极少,将兴安杜鹃分为6个龄级:地径≤4mm为Ⅰ龄级,地径4—6mm为Ⅱ龄级,地径6—8mm为Ⅲ龄级,地径8—10mm为Ⅳ龄级,地径10—12mm为Ⅴ龄级,地径≥12mm为Ⅵ龄级。其中,Ⅰ—Ⅱ为幼龄级,Ⅲ—Ⅳ为中龄级,Ⅴ—Ⅵ为老龄级。

表1 样地特征
1.3 静态生命表
以径级代替龄级的方法,统计各径级的个体数,进而编制了兴安杜鹃种群的静态生命表[24—26]。5个样地被视为5个相互独立种群。静态生命表中,纵表头为划分的龄级,横表头生命表参数,包括ax、lx、lnlx、dx、qx、Lx、Tx、ex、Kx和Sx。其中,x为龄级,ax为x龄级内现有的存活数;lx为存活份数,即x龄级开始时的标准化存活数;dx为x龄级和x+1龄级间标准化死亡数;qx为x龄级的个体死亡率;Lx为区间寿命,即x龄级和x+1龄级间的存活个数;Tx为总寿命,即x龄级及其以上各龄级的存活个体总数;ex为期望寿命(生命期望或存活余年),即总寿命与该龄级标准化存活个体数之比;Kx为消失率;Sx为存活率。各参数计算公式如下:
dx=lx-lx+1
Tx=∑xLx
Kx=lnlx-lnlx+1
1.4 生存分析函数
生存分析函数包括种群生存率函数S(i)、累计死亡率函数F(i)、死亡密度函数f(ti)和危险率函数λ(ti),可以更全面地分析种群数量变化[27—28]。4种函数计算公式如下:
S(i)=S1×S2×S3,…,×Si
F(i)=1-S(i)
式中,hi为龄级宽度。
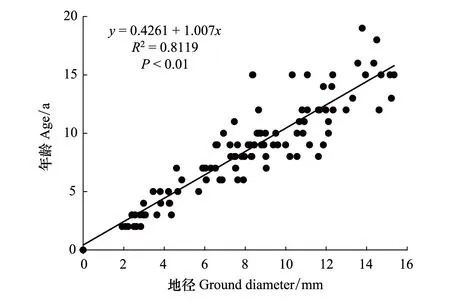
图3 兴安杜鹃地径与年龄的关系 Fig.3 Relationship between ground diameter and age of Rhododendron dauricum
1.5 时间序列分析
采用时间序列分析中的一次移动平均法,对种群数量未来发展趋势进行了预测[18, 29]。计算公式如下:

2 结果与分析
2.1 兴安杜鹃年轮和地径的关系
根据兴安杜鹃地径和年龄的拟合结果,二者存在明显的线性关系,拟合曲线为y=0.4261+1.007x(R2=0.8119,P<0.01),x为兴安杜鹃地径(mm),y为兴安杜鹃年龄(a)(图3)。
2.2 兴安杜鹃种群静态生命表
根据种群各龄级的个体数,计算生命表参数,得到兴安杜鹃种群的静态生命表(表2)。此外,为了保持数据的准确性,并没有对数据进行平滑处理,因此对于部分数据死亡率存在负值。结果表明,低海拔梯度(600m)的落叶松林中(种群A),兴安杜鹃幼龄级个体数量最多,占个体总数的67.31%。幼龄级个体的死亡率和消失率较低,存活率较高。因此,种群A为增长型种群。中低海拔梯度(600—900m)的白桦林中(种群B、C、D),中龄级个体数最多,分别占个体总数的47.49%、47.96%和45.64%。Ⅰ—Ⅲ龄级个体的死亡率和消失率较低,存活率较高,且期望寿命表明个体均可存活到Ⅳ或Ⅴ龄级。因此,种群B、C、D为稳定型种群。高海拔梯度(1200m)的落叶松林中(种群E),种群个体总数远少于其他种群。Ⅰ龄级个体数量最多,占个体总数的33.52%,死亡率和消失率较高,存活率较低,表明Ⅰ龄级个体成长为Ⅱ龄级的概率较低。而Ⅱ—Ⅵ龄级的个体数量较为接近,Ⅱ—Ⅴ龄级死亡率和消失率较低,存活率较高,且期望寿命表明个体均可存活到Ⅴ或Ⅵ龄级。因此,种群E为增长型种群。

表2 兴安杜鹃种群生命表
2.3 兴安杜鹃种群存活曲线
种群的存活曲线表明,中低海拔梯度种群(A、B、C、D)的存活曲线为Deevey Ⅰ型曲线,即凸曲线,表明种群早期死亡率较低,种群内大多数个体能活到一定的生理年龄,之后死亡率上升,个体逐渐死亡(图4)。而高海拔梯度的种群(E)为Deevey Ⅲ型曲线,即凹曲线,表明种群早期死亡率较高,种群内大多数个体无法生长成为成熟个体,但一旦生长到一定生理年龄,死亡率大大降低。
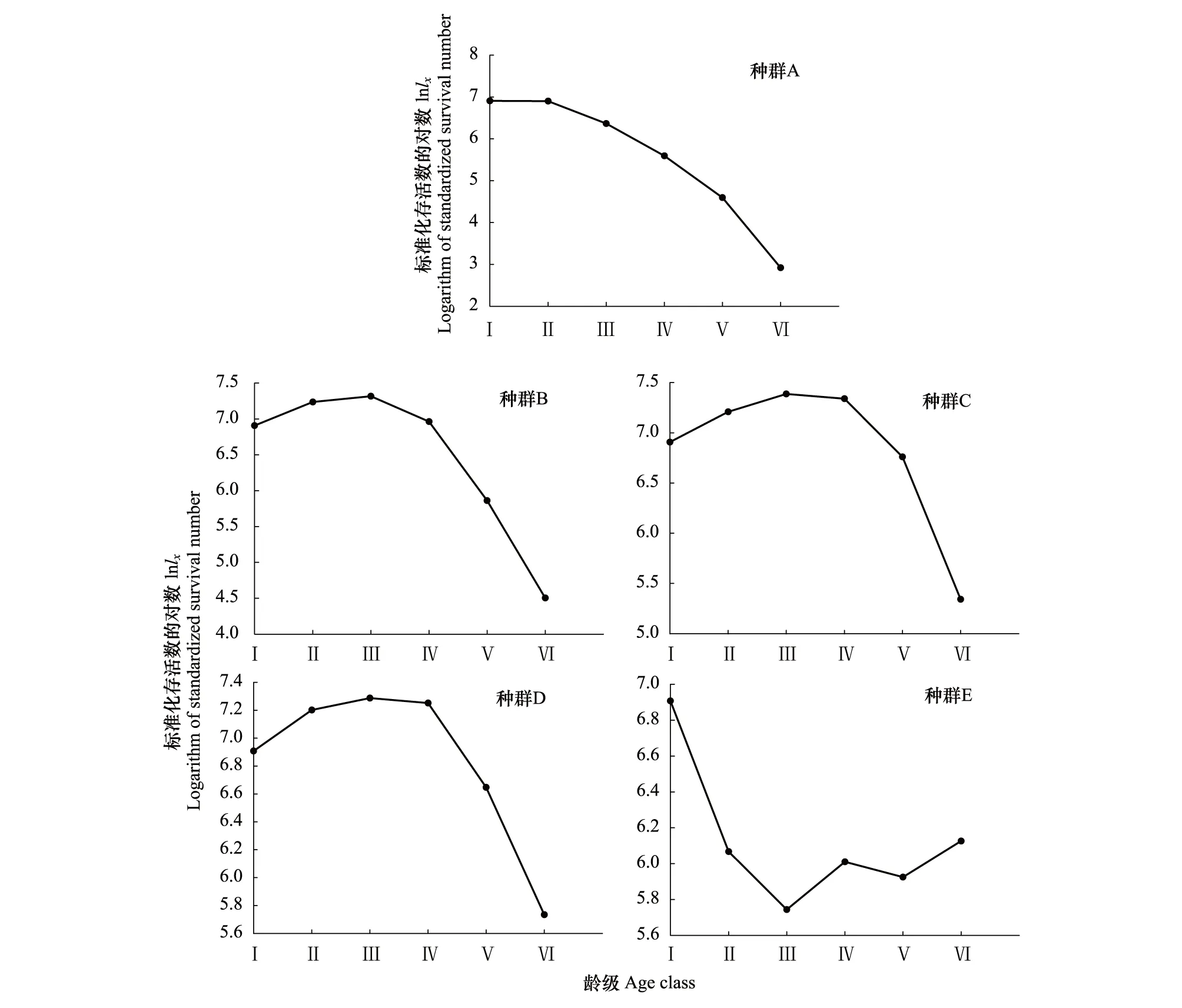
图4 兴安杜鹃种群存活曲线Fig.4 The survival curves of Rhododendron dauricum populationA、B、C、D、E为种群编号,分别代表代表样地A、B、C、D、E内的兴安杜鹃种群;Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ为龄级编号,分别代表地径≤4mm、4—6mm、6—8mm、8—10mm、10—12mm、≥12mm的个体
2.4 兴安杜鹃种群生存分析函数
种群生存分析函数表明,低海拔落叶松林中(种群A),随龄级增大,生存率逐渐降低,累积死亡率和危险率逐渐升高。在Ⅱ和Ⅲ龄级之间,生存率和累计死亡率达到平衡,死亡密度最高,此阶段之后种群进入衰退期(表3)。中低海拔白桦林中(种群B、C、D),在Ⅰ—Ⅲ龄级阶段,生存率较高且基本一致,之后逐渐下降,危险率则相反。累积死亡率与生存率为互补趋势,种群B、C、D分别在Ⅲ—Ⅳ、Ⅳ—Ⅴ和Ⅳ—Ⅴ龄级达到平衡,同时死亡密度达到最高点,之后种群进入衰退期。高海拔落叶松林中(种群E),Ⅰ—Ⅴ龄级生存率较为接近且均小于50%,累积死亡率均大于50%。随龄级增大,危险率呈下降趋势,死亡密度在Ⅰ和Ⅳ—Ⅴ龄级阶段较高,表明种群早期死亡率很高,在Ⅳ—Ⅴ龄级阶段之后,种群进入衰退期。

表3 兴安杜鹃种群生存分析函数计算值
2.5 兴安杜鹃种群预测
时间序列分析表明,低海拔落叶松林中(种群A),经过1、2、3、4个龄级时间后,各龄级的个体数量均呈增加趋势,表明种群未来数量为增长趋势(图5)。中低海拔白桦林(种群B、C、D),经过1、2、3、4个龄级时间后,Ⅱ和Ⅲ龄级的个体数量呈下降趋势,Ⅴ和Ⅵ龄级的个体数量呈增加趋势,表明种群在未来阶段逐渐趋于衰退。对于高海拔落叶松林种群(E),经过1、2、3、4个龄级时间后,低龄级个体呈增加趋势,高龄级个体呈下降趋势。因此,在未来阶段,种群E有一定程度的增长趋势,但趋向稳定。

图5 兴安杜鹃种群时间序列分析Fig.5 Time series analysis of Rhododendron dauricum population
3 讨论
3.1 兴安杜鹃种群数量总体现状
根据生命表分析结果,大兴安岭兴安杜鹃种群为增长型或稳定型种群,即在没有外部干扰的条件下,种群近期不存在衰退或消失的风险。从系统进化的角度讲,杜鹃属植物对高海拔和寒冷的环境都有一定的适应能力[30]。兴安杜鹃为半常绿灌木,幼枝被柔毛和鳞片,叶片近革质,因此适宜分布于寒冷环境。成熟植株高度最高可达2m,在灌木层中具有较强的竞争优势。自身生物学和生态位特征决定兴安杜鹃种群数量具有一定稳定性[20, 31]。
研究表明杜鹃属植物种群的数量特征则存在一定程度的种间差异。金慧等[24]研究了长白山地区牛皮杜鹃(R.aureum)的种群数量特征,发现其为衰退型种群,且海拔越高,种群的衰退期越早。郭其强等[32]研究了西藏色季拉山薄毛海绵杜鹃(R.aganniphum)的种群年龄结构,发现其幼龄个体数量相对不足。李贵祥等[33]研究了云南省腾冲县的大树杜鹃(R.protistum)种群数量特征,发现其为稳定性种群。其中,牛皮杜鹃为濒危植物,大树杜鹃为极危植物。牛皮杜鹃自身幼苗存活率低,繁殖能力差,且生长在高海拔的严苛环境中,种群呈衰退趋势。大树杜鹃为高大乔木,目前数量较少主要是受人为干扰的影响,随着近年来保护力度的加强,种群数量趋于稳定。对于薄毛海绵杜鹃,幼龄级个体缺乏也是由于其生长的高海拔环境不利于幼苗生长。可以认为,影响杜鹃属植物种群数量的主要因素是自身生物学和生态学特点,其次是外部环境条件,其中人为干扰不可忽视。
3.2 兴安杜鹃种群数量动态
时间序列分析结果表明,白桦林中兴安杜鹃种群(B、C、D),随着时间的推移,幼龄级和中龄级个体数量减少,老龄级个体数量增加,种群呈衰退趋势。究其原因可以认为是种内的“自疏效应”在发挥主导作用。种群B、C、D的种群密度较高,占5个种群个体总数的84.25%,通常会导致种内竞争加剧,进而可能导致“自疏”效应[34]。大量成熟个体的存在,必然消耗大量资源,导致种内竞争力相对较弱的幼龄级个体生长空间狭小,幼龄个体存活率下降。有研究表明,荒漠绿洲过渡带的柽柳(Tamarixchinensis)种群,随着径级的增大,个体对环境资源的争夺,加剧了种内竞争,从而使个体数量递减[35]。胡尔查等[36]研究了乌拉山自然保护区的白桦种群结构,发现在Ⅲ、Ⅳ径级种内竞争最为激烈,为了争夺生长空间和必需的光照,产生了自疏现象。因此,对于种群B、C、D,可以认为种内竞争是导致未来种群数量下降的重要原因。这里需强调的是,这种衰退也是可逆的,随着生存空间的变化,种群数量有趋于稳定的可能性。
3.3 兴安杜鹃种群数量与海拔和林型的关系
立地条件是影响植物种群数量的重要因素[37]。王立龙等[18]研究了不同立地条件下裸果木(Gymnocarposprzewalskii)种群数量特征,发现水冲滩地的种群数量最大,其次是山间冲沟,最后是平缓戈壁。而海拔和林型均是立地条件的重要组成因素,5个兴安杜鹃种群的立地条件可划分为3个海拔高度和2种林型。随着海拔高度的上升,兴安杜鹃种群大小有下降的趋势,种群B和C总个体数最多,种群A和D总体数量相对较少,而种群E最少,且远远小于前四个种群,仅占5个种群总数量的1.25%。可以认为海拔是制约兴安杜鹃种群数量的重要因素。高海拔区域寒冷、多风、乔木层稀疏且地表多为山石、土壤结构与厚度不良,不利于兴安杜鹃生存。同时在高海拔梯度,灌木层中偃松占据绝对优势,使兴安杜鹃的生态位宽度受到压缩[20]。
种群生命表和生存分析函数等分析结果表明,低海拔落叶松林中种群(A)幼龄级个体数量较多,且种群衰退期的转折点在Ⅱ和Ⅲ龄级之间,这远远早于其他4个种群。这是因为落叶松林郁闭度较大,无法保障兴安杜鹃生长所需的光照条件。对于高海拔落叶松林中种群(E),幼年个体的死亡率很高,特别是龄级Ⅰ到龄级Ⅱ的死亡率为0.569,因此年幼的个体很难生长成为成熟个体。这与种群E所在的高海拔环境有关,当个体到达龄级Ⅱ之后,死亡率大大降低,表明兴安杜鹃幼龄虽然不适宜海拔1200m的环境,但成熟个体能够在这一海拔高度正常生存。对于中低海拔白桦林下种群(B、C、D),种群数量较为稳定,表明其立地条件适宜兴安杜鹃生存,即海拔梯度600—900m、郁闭度45%—50%的环境。因此,兴安杜鹃对海拔和光照条件具有一定的需求,这与其他杜鹃属植物的相关研究一致[1, 38]。比如,在重庆白马山林场,灌木状、大灌木状和乔木状的杜鹃属植物分布的海拔梯度逐渐升高,需要的光照条件逐渐从中等日照强度向强日照、光照充足的环境过渡[39]。在云南省小白草岭,林分密度和郁闭度较大,使大王杜鹃(R.rex)种子光照不足而影响萌发和向幼苗、幼树转化[40]。
4 结论
大兴安岭兴安杜鹃种群一般为增长型或稳定型种群,幼龄级和中龄级(Ⅰ—Ⅳ)的个体,即地径在0—10mm的个体数量较多,且个体的死亡率和消失率较低,存活率较高,种群的期望寿命为Ⅳ或Ⅴ龄级,即个体存活至后期的概率较大。目前兴安杜鹃种群整体稳定,海拔和林型是影响兴安杜鹃种群数量的重要因素。600—900m海拔梯度中,兴安杜鹃种群数量较多,是灌木层中的优势种。而1200m海拔梯度中,兴安杜鹃种群数量较少,不再是灌木层中的优势种,且Ⅰ龄级的个体死亡率和消失率较高,存活率较低。郁闭度45%—50%的白桦林中,兴安杜鹃种群数量较多,为稳定型种群。而落叶松林中,郁闭度相对较高,兴安杜鹃种群数量相对较少。
猜你喜欢
农业科技与信息(2022年12期)2022-08-09
西北农林科技大学学报(自然科学版)(2021年7期)2021-08-09
心声歌刊(2021年3期)2021-08-05
草原歌声(2020年3期)2021-01-18
河南农业大学学报(2020年2期)2020-05-22
小聪仔(幼儿版)(2020年12期)2020-02-01
文苑(2019年22期)2019-12-07
草原歌声(2018年2期)2018-12-03
民族音乐(2018年5期)2018-11-17
乡村地理(2018年3期)2018-11-06
