
YWHAZ在小鼠早期胚胎发育过程中的作用
2022-07-25 06:35:18赵怡凡马增友方俊博孟碟方
中国兽医学报 2022年3期
赵怡凡,马增友,方俊博,孟碟方,彭 辉
(福建农林大学 动物科学学院(蜂学学院),福建 福州 350002)
酪氨酸-色氨酸羟化酶激活蛋白(tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein,又称为YWHA蛋白)普遍存在于真核生物中且高度保守。YWHA蛋白通常以单体相对分子质量约为30 kDa的同源或异源二聚体形式存在。该蛋白家族在哺乳动物中有7个成员,分别为YWHAB、YWHAG、YWHAQ、YWHAH、YWHAZ、YWHAE和SFN[1-3]。YWHA蛋白又被称为磷酸化丝氨酸/磷酸化苏氨酸结合蛋白,有研究表明,YWHA蛋白可以与靶蛋白的磷酸化位点结合,如激酶、磷酸酶、跨膜受体、转录因子等[4]。通过蛋白质组学和生物化学方法已经检测到200多种可能与YWHA蛋白相互作用的蛋白质,这些蛋白参与调控细胞周期、信号转导、应激反应、细胞凋亡、细胞代谢等生命活动[5-6]。YWHA蛋白可结合特定的磷酸化丝氨酸和磷酸化苏氨酸序列,并通过改变靶蛋白细胞定位、改变酶活性和调节蛋白-蛋白相互作用来发挥功能[7-9]。
YWHAZ是YWHA蛋白家族成员之一,YWHAZ参与多种细胞信号通路,在细胞增殖和细胞周期中发挥作用[10]。有研究表明,C.elegans的PAR-5蛋白与哺乳动物中的YWHAZ蛋白高度同源[11],PAR-5在C.elegans早期胚胎非对称发育中起重要作用,PAR-5蛋白水平显著降低会导致胚系缺陷和不育[12]。YWHAZ蛋白在哺乳动物早期胚胎发育过程中的研究尚未见报道,本试验通过RT-PCR、Western blot、免疫荧光和RNAi等方法研究YWHAZ蛋白在小鼠早期胚胎发育过程中的表达、定位和作用。
1 材料与方法
1.1 实验动物雌性和雄性ICR小鼠购自福建省福州市闽侯县吴氏实验动物中心,小鼠饲养条件良好,自由采食及饮水,饲养过程中的昼夜明暗循环为14 h/10 h,室温控制在25℃。
1.2 主要试剂人绒毛膜促性腺激素(hCG)和孕马血清促性腺激素(PMSG)购自宁波第二激素厂;YWHAZ兔多克隆抗体和KSOM培养液购自Sigma;Ywhaz siRNA购自Santa Cruz;siPORTTMAmine Transfection Agent购自Applied Biosystems;CellAmpTMWhole Transcriptome Amplification Kit购自TaKaRa。
1.3 早期胚胎的收集与培养8~10周龄雌性小鼠腹腔注射10 IU PMSG刺激卵泡生长发育,48 h后注射10 IU hCG促排,将处理后的母鼠与公鼠合笼,14 h后检查阴道栓,采取脱颈法将有阴道栓的雌性小鼠处死。取出输卵管置于H-KSOM操作液中,在体视显微镜下用无菌的1 mL皮下注射器划破输卵管,释放卵丘卵母细胞复合体,再将卵丘卵母细胞复合体吸至透明质酸酶溶液中,消化卵丘细胞收集合子,将合子转移至平衡好的培养皿中,在37℃、5% CO2饱和湿度下恒温培养。
1.4 RT-PCR和qRT-PCR使用CellAmpTMWhole Transcriptome Amplification Kit试剂盒分别将10个1-细胞、2-细胞、4-细胞、8-细胞、桑葚胚和囊胚直接逆转录合成cDNA,然后对反转录的cDNA进行扩增。合成的cDNA按50倍稀释后作为模板分别进行RT-PCR和qRT-PCR扩增,特异性引物参见表1。
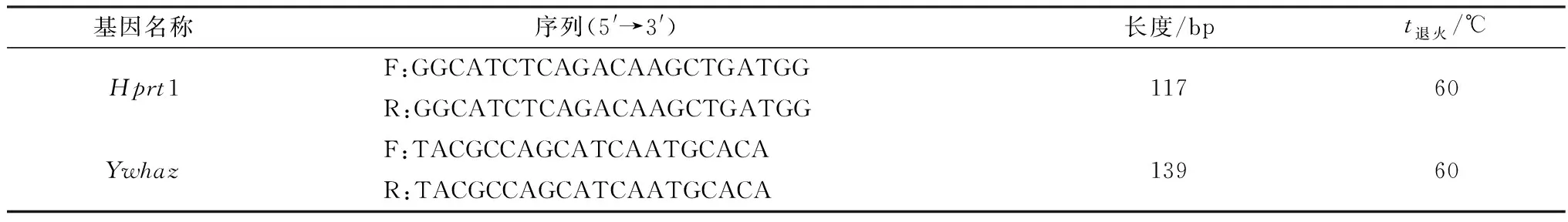
表1 基因特异性引物信息
1.5 Western blot检测收集1-细胞、2-细胞、4-细胞、8-细胞、桑葚胚和囊胚各50个,将胚胎加入含有全蛋白酶抑制剂(Roche)和IP裂解液(1∶25)的离心管中,冰上孵育2 min使胚胎充分裂解,再加入SDS上样缓冲液,煮沸变性10 min后进行SDS-PAGE电泳,用湿转法将目的蛋白转移到NC膜上,5%封闭液室温孵育4 h,之后将NC膜浸入一抗中4℃孵育过夜。0.1% TBST摇洗3次,IRDye®近红外荧光二抗按1∶10 000的比例稀释,NC膜浸入二抗溶液中室温避光孵育2 h。0.1% TBST摇洗3次后,再用TBS摇洗3次,将NC膜置于双色红外激光成像系统Odyssey CLx中曝光拍照。
1.6 免疫荧光检测收集1-细胞、2-细胞、4-细胞、8-细胞、桑葚胚和囊胚各10个,PBS洗涤3次,将各时期胚胎置于4%多聚甲醛溶液中固定1 h。之后将胚胎吸入0.1% Triton X-100中,室温孵育20 min,增加细胞膜通透性。通透打孔后的胚胎在免疫染色封闭液中室温封闭4 h,将封闭后的胚胎吸入一抗溶液中4℃孵育过夜。免疫荧光洗涤液摇洗3次后加入荧光二抗,室温避光孵育2 h。免疫荧光洗涤液摇洗3次,加入DAPI染色液染核10 min,免疫荧光洗涤液摇洗3次后将胚胎吸至载玻片上,滴加荧光抗淬灭剂封片,在荧光显微镜下观察拍照。
1.7 siRNA转染收集合子置于酸性台氏液中10~20 s消化透明带,然后向培养皿中加入1 mL H-KSOM终止消化,吸出合子用H-KSOM清洗2次备用。在Opti-MEM培养基中加入siPORTTMAmine Transfection Agent,室温孵育10 min。将siRNA加入到Opti-MEM培养基中。将上述配好的试剂1∶1混合,室温静置孵育10 min使siRNA和转染试剂充分结合。用Opti-MEM冲洗电转皿,最后吸取混合液加入电转皿的两根电极之间,再将合子加入到电转皿的凹槽中并使合子呈线性排列。电转仪的参数设置为:电压22 V、脉冲时间1 ms、脉冲间隔500 ms、脉冲次数3次。电转之后,用吸卵针吸出处理后的合子并在H-KSOM操作液中清洗,置于平衡好的KSOMaa培养液中,在37℃、5% CO2饱和湿度下恒温培养。
1.8 统计分析各试验均独立重复至少3次,所有分析均采用SPSS 19.0软件,使用单因素方差分析进行组间比较,P<0.05为差异显著,P<0.01为差异极显著。
2 结果
2.1YwhazmRNA及蛋白在小鼠早期胚胎发育过程中的表达利用RT-PCR方法对Ywhaz在小鼠早期胚胎发育过程中的表达进行检测,结果表明,YwhazmRNA在1-细胞、2-细胞、4-细胞、8-细胞、桑葚胚和囊胚中均有表达(图1A)。同时,提取不同时期早期胚胎的蛋白,经过SDS-PAGE电泳使蛋白分离,再通过Western blot检测YWHAZ蛋白表达情况,结果显示,在早期胚胎发育过程中均能检测到该蛋白的表达(图1B)。
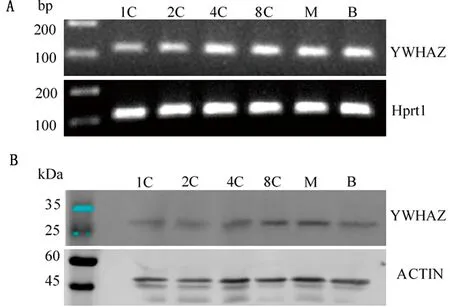
A.Ywhaz mRNA在小鼠早期胚胎中的表达;B.YWHAZ 蛋白在小鼠早期胚胎中的表达
2.2 YWHAZ蛋白在小鼠早期胚胎中的定位取小鼠的1-细胞、2-细胞、4-细胞、8-细胞、桑葚胚和囊胚进行免疫荧光染色,研究YWHAZ蛋白在小鼠早期胚胎中的定位。结果表明,YWHAZ蛋白定位在小鼠早期胚胎的细胞质和细胞核中(图2)。
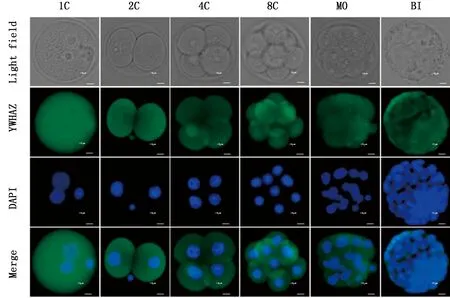
蓝色荧光为细胞核;绿色荧光为YWHAZ蛋白
2.3YwhazsiRNA的沉默效率为了研究YWHAZ在早期胚胎发育过程中的作用,首先通过电转方式向1-细胞胚胎中转入YwhazsiRNA,检测基因沉默效率。结果显示,当YwhazsiRNA浓度达到0.7 μmol/L时,该基因的沉默效率为91%(图3A)。使用Western blot检测干扰组与阴性对照组的YWHAZ蛋白表达差异,结果表明,Ywhaz干扰培养24 h后,YWHAZ蛋白表达明显减少(图3B)。经Image studio软件分析显示,YWHAZ蛋白表达量减少了45%(图3C)。
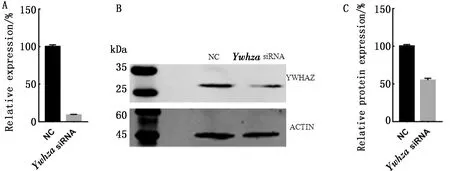
A..沉默Ywhaz降低其mRNA水平的表达;B.沉默Ywhaz降低其蛋白水平的表达;C.YWHAZ蛋白在对照组和干扰组中的表达分析
2.4 沉默Ywhaz对小鼠早期胚胎发育的影响为了研究YWHAZ在小鼠早期胚胎发育过程中是否发挥作用,本试验将YwhazsiRNA转入1-细胞胚胎,之后在体外将其培养至囊胚。胚胎发育4.5 dpc时,其形态学特征如图4A所示。阴性对照组与干扰组之间发育情况存在明显差异,干扰组2-细胞、4-细胞、8-细胞、桑葚胚和囊胚的发育率极显著低于对照组(P< 0.01)。在4.5~5.0 dpc的过程中,对照组囊胚率无明显变化,而干扰组囊胚率显著增加,但
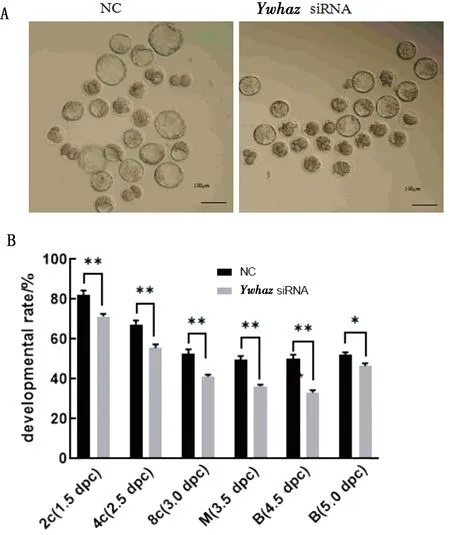
A.Ywhaz沉默后的胚胎培养4.5 dpc的形态学特征;B.培养不同天数的胚胎发育百分率;*P<0.05;** P<0.01
对照组与干扰组间仍存在显著性差异(P<0.05)(图4B),表明沉默Ywhaz可导致小鼠早期胚胎发育率降低,虽大部分胚胎能发育到囊胚,但存在明显的胚胎发育延迟现象。
3 讨论
本研究通过RT-PCR和Western blot技术检测了YwhazmRNA及其编码的蛋白在小鼠早期胚胎中的表达情况。结果发现,YwhazmRNA在小鼠胚胎发育过程中均有表达,并且表达较为稳定。本试验结果与Ywhaz基因在人、兔、牛等早期胚胎发育过程中的表达情况一致[13-15]。本研究结果显示,YWHAZ蛋白在小鼠早期胚胎的各个时期均有表达,同时定位于早期胚胎的细胞质与细胞核中,提示该蛋白可能在小鼠早期胚胎发育过程中发挥一定的作用。
小鼠受精卵在体内经过3.5 dpc可发育至囊胚,体外培养条件下需要4.0~4.5 dpc发育至囊胚。本研究采取1.5,2.5,3.0,3.5,4.5 dpc时观察2-细胞、4-细胞、8-细胞、桑葚胚和囊胚发育情况。结果显示,Ywhaz沉默组各时期发育率都显著低于对照组,表明沉默Ywhaz可导致小鼠早期胚胎发育率降低。将观察时间延长至5.0 dpc,沉默组囊胚率显著增高,但仍低于对照组,提示沉默Ywhaz可影响早期胚胎发育进程,囊胚发育延迟。有研究表明,YWHAZ蛋白通过与CDC25蛋白结合并调节其功能而参与了G1/S期转变和G2/M期转变,在有丝分裂间期发挥作用[16]。此外,YWHAZ与Polo样激酶1(polo-like kinase 1,Plk1)在有丝分裂过程中存在相互作用,并共同定位于胞质分裂过程中的中心体。YWHAZ的缺失导致细胞染色体分离滞后,处于胞质分裂期的细胞明显增多[17],这也可能是沉默Ywhaz延迟早期胚胎发育的原因。
本试验虽然使用siRNA将Ywhaz基因的表达量降低了91%,但YWHAZ蛋白表达量仅降低了45%,早期胚胎中残留的YWHAZ蛋白可以继续支持早期胚胎发育。此外,YWHA家族成员在结构和功能上类似,不同的亚型可以与相同的配体结合。因此,YWHA家族其他成员也可能代偿了YWHAZ蛋白的功能,以致Ywhaz沉默组大部分胚胎仍可以发育至囊胚。研究表明,YWHAZ通过形成同源或异源二聚体与靶蛋白结合而发挥作用[18]。在早期胚胎发育过程中,YWHAZ也可能与其他蛋白形成复合体进而调节早期胚胎的发育,但该蛋白与何种蛋白形成复合体还有待于进一步的研究。
综上所述,本研究利用RT-PCR、Western blot、免疫荧光和RNAi技术阐明了YwhazmRNA及其编码的蛋白在小鼠早期胚胎发育过程中的表达、定位和作用,揭示了在合子中沉默Ywhaz基因可导致小鼠早期胚胎发育率降低,且出现囊胚发育延迟现象。
猜你喜欢
中国生殖健康(2020年5期)2021-01-18 02:59:48
生殖医学杂志(2020年12期)2020-12-22 03:26:18
浙江医学(2020年17期)2020-09-21 09:06:30
中国生殖健康(2018年5期)2018-11-06 07:15:38
中成药(2017年9期)2017-12-19 13:34:27
CHINESE JOURNAL OF AERONAUTICS(2017年1期)2017-11-21 12:54:14
中成药(2017年5期)2017-06-13 13:01:12
新疆医科大学学报(2015年10期)2015-12-26 12:33:30
中国医学科学院学报(2015年5期)2015-03-01 04:03:36
郑州大学学报(医学版)(2015年2期)2015-02-27 14:50:51
